Investigation of Solanum carolinense Dominance and Phytotoxic Effect in Festuca arundinacea with Special Reference to Allelochemical Identification, Analysis of Phytohormones and Antioxidant Mechanisms

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Investigation of Competition Patterns between Festuca arundinacea and Solanum carolinense
2.2.1. Investigation of Competition Patterns according to the Seeding Ratio of Festuca arundinacea and Solanum carolinense
2.2.2. Investigation of Growth Patterns of Festuca arundinacea according to the Growth Period of Solanum carolinense
2.3. Investigation of Phytotoxicity of SCE on Festuca arundinacea
2.3.1. Preparation of Crude Extract
2.3.2. Treatment of Crude Extract in Festuca arundinacea
2.4. Determination of Reactive Oxygen Species (ROS)
- Superoxide anion radical (O2) measurement
- Hydrogen peroxide (H2O2) measurement
- Visualization of ROS (H2O2)
2.5. Determination of Antioxidant Enzyme
- Superoxide dismutase (SOD) measurement
- Catalase (CAT) measurement
- Peroxidase (POD) measurement
2.6. Quantification of Endogenous Phytohormones
- Abscisic acid (ABA) measurement
- Salicylic acid (SA) measurement
2.7. Identification of Allelochemical in Solanum carolinense
2.7.1. Herbicidal Activity Assay
2.7.2. Isolation, Purification, and Identification of Allelochemicals
2.8. Statistical Analysis
3. Results
3.1. Investigation of Competition Patterns
3.1.1. Evaluation of Competition Patterns according to the Seeding Ratio
3.1.2. Competition Patterns according to the Growth Period
3.2. Investigation of Phytotoxicity of SCE on Festuca arundinacea
3.2.1. Effect on Morphological Characteristics and Chlorophyll Content
3.2.2. Determination of Reactive Oxygen Species (ROS)
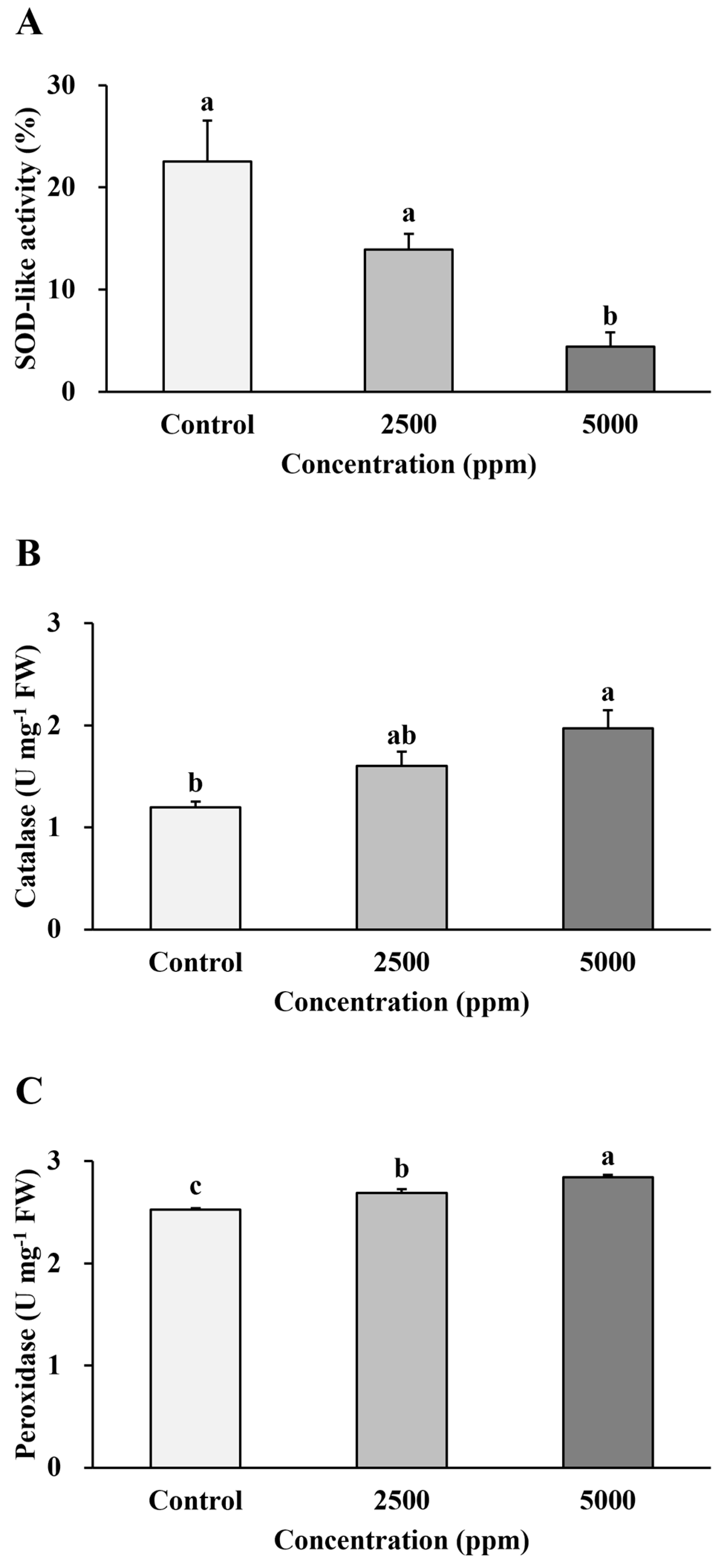
3.2.3. Determination of Antioxidant Enzyme
3.2.4. Abscisic Acid (ABA) and Salicylic Acid (SA) Quantification
3.3. Identification of Allelochemical in Solanum carolinense
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
References
- Barnes, R.F. Importance and problems of tall fescue. In Biotechnology in Tall Fescue Improvement; CRC Press: Boca Raton, FL, USA, 2018; pp. 1–12. [Google Scholar]
- Chen, L.; Auh, C.K.; Dowling, P.; Bell, J.; Chen, F.; Hopkins, A.; Dixon, R.A.; Wang, Z.Y. Improved forage digestibility of tall fescue (Festuca arundinacea) by transgenic down-regulation of cinnamyl alcohol dehydrogenase. Plant Biotechnol. J. 2003, 1, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Shahabzadeh, Z.; Mohammadi, R.; Darvishzadeh, R.; Jafari, M.; Breeding, F.P.; Research, G. Investigation of genetic diversity of forage yield and morphological traits in tall fescue (Festuca arundinacea Schreb.) populations. Mol. Biol. Rep. 2020, 28, 17–36. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.H.; Lee, E.J. Resistance of plant communities to invasion by tall fescue. An experimental study combining species diversity, functional traits and nutrient levels. Basic Appl. Ecol. 2022, 58, 39–49. [Google Scholar] [CrossRef]
- Tanveer, A.; Khaliq, A.; Ali, H.H.; Mahajan, G.; Chauhan, B.S. Interference and management of parthenium: The world’s most important invasive weed. Crop Prot. 2015, 68, 49–59. [Google Scholar] [CrossRef]
- Follak, S.; Strauss, G. Potential distribution and management of the invasive weed Solanum carolinense in Central Europe. Weed Res. 2010, 50, 544–552. [Google Scholar] [CrossRef]
- Fekry, M.I.; Ezzat, S.M.; Salama, M.M.; Alshehri, O.Y.; Al-Abd, A.M. Bioactive glycoalkaloides isolated from Solanum melongena fruit peels with potential anticancer properties against hepatocellular carcinoma cells. Sci. Rep. 2019, 9, 1746. [Google Scholar] [CrossRef]
- Altesor, P.; García, Á.; Font, E.; Rodríguez-Haralambides, A.; Vilaró, F.; Oesterheld, M.; Soler, R.; González, A. Glycoalkaloids of Wild and Cultivated Solanum: Effects on Specialist and Generalist Insect Herbivores. J. Chem. Ecol. 2014, 40, 599–608. [Google Scholar] [CrossRef]
- Sun, F.; Li, S.; He, D.; Cao, G.; Ni, X.; Tai, G.; Zhou, Y.; Wang, D. Effects of glycoalkaloids from Solanum plants on cucumber root growth. Phytochemistry 2010, 71, 1534–1538. [Google Scholar] [CrossRef]
- Waseem, M.; Badruzzaman, S.M. Allelopathy in Solanaceae plants. J. Plant Prot. Res. 2018, 58, 1–7. [Google Scholar] [CrossRef]
- Solomon, B.P. Autoallelopathy in Solanum carolinense: Reversible delayed germination. Am. Midl. Nat. 1983, 110, 412–418. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and Exotic Plant Invasion: From Molecules and Genes to Species Interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A biogeographical approach to plant invasions: The importance of studying exotics in their introduced and native range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Schandry, N.; Becker, C. Allelopathic plants: Models for studying plant–interkingdom interactions. Trends Plant Sci. 2020, 25, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowska, A. Allelochemicals as Bioherbicides—Present and Perspectives. In Herbicides—Current Research and Case Studies in Use; IntechOpen: Dallas, TX, USA, 2013. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef] [PubMed]
- Macias, F.A.; Molinillo, J.M.; Varela, R.M.; Galindo, J.C. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar] [CrossRef]
- Tlak Gajger, I.; Dar, S.A. Plant allelochemicals as sources of insecticides. Insects 2021, 12, 189. [Google Scholar] [CrossRef]
- Saraf, M.; Pandya, U.; Thakkar, A. Role of allelochemicals in plant growth promoting rhizobacteria for biocontrol of phytopathogens. Microbiol. Res. 2014, 169, 18–29. [Google Scholar] [CrossRef]
- Vyvyan, J.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. J. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef]
- Bogatek, R.; Gniazdowska, A. ROS and phytohormons in plant-plant allelopathic interaction. Plant Signal. Behav. 2007, 2, 317–318. [Google Scholar] [CrossRef]
- Ali, M.; Ahmad, H.; Hayat, S.; Ghani, M.I.; Amin, B.; Atif, M.J.; Wali, K.; Cheng, Z. Application of garlic allelochemicals improves growth and induces defense responses in eggplant (Solanum melongena) against Verticillium Dahliae. Ecotoxicol. Environ. Saf. 2021, 215, 112132. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef] [PubMed]
- Staszek, P.; Krasuska, U.; Ciacka, K.; Gniazdowska, A. ROS Metabolism Perturbation as an Element of Mode of Action of Allelochemicals. Antioxidants 2021, 10, 1648. [Google Scholar] [CrossRef] [PubMed]
- Das, C.; Dey, A.; Bandyopadhyay, A. Allelochemicals: An emerging tool for weed management. In Evidence Based Validation of Traditional Medicines; Springer: Singapore, 2021; pp. 249–259. [Google Scholar] [CrossRef]
- Kong, C.-H.; Xuan, T.D.; Khanh, T.D.; Tran, H.-D.; Trung, N.T. Allelochemicals and Signaling Chemicals in Plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [PubMed]
- Navari-Izzo, F.; Pinzino, C.; Quartacci, M.F.; Sgherri, C.L. Superoxide and hydroxyl radicai generation, and superoxide dismutase in PII membrane fragments from wheat. Free Radic. Res. 1999, 31, 3–9. [Google Scholar] [CrossRef]
- Jana, S.; Choudhuri, M.A. Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquat. Bot. 1982, 12, 345–354. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B.J. Subcellular localization of H2O2 in plants. H2O2 accumulation in papillae and hypersensitive response during the barley—Powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 673–684. [Google Scholar]
- Pütter, J. Peroxidases. In Methods of Enzymatic Analysis; Elsevier: Amsterdam, The Netherlands, 1974; pp. 685–690. [Google Scholar]
- Kang, S.-M.; Adhikari, A.; Lee, K.-E.; Khan, M.A.; Shahzad, R.; Dhungana, S.K.; Lee, I.-J. Inoculation with Indole-3-acetic acid-producing rhizospheric Rhodobacter sphaeroides KE149 augments growth of adzuki bean plants under water stress. J. Microbiol. Biotechnol. 2020, 30, 717–725. [Google Scholar] [CrossRef]
- Seskar, M.; Shulaev, V.; Raskin, I. Endogenous methyl salicylate in pathogen-inoculated tobacco plants. Plant Physiol. 1998, 116, 387–392. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.M.; Nguyen, V.; Schroeder, J.I. The role of reactive oxygen species in hormonal responses. Plant Physiol. 2006, 141, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Zafra, A.; Rodríguez-García, M.I.; Alché, J.d.D. Cellular localization of ROS and NO in olive reproductive tissues during flower development. BMC Plant Biol. 2010, 10, 36. [Google Scholar] [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Kusvuran, S.; Kiran, S.; Ellialtioglu, S.S. Antioxidant enzyme activities and abiotic stress tolerance relationship in vegetable crops. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Intech: Dallas, TX, USA, 2016; pp. 481–506. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z.-H.; Meng, H.-W. Transcriptomic insights into the allelopathic effects of the garlic allelochemical diallyl disulfide on tomato roots. Sci. Rep. 2016, 6, 38902. [Google Scholar] [CrossRef]
- Escobar-Bravo, R.; Ruijgrok, J.; Kim, H.K.; Grosser, K.; Van Dam, N.M.; Klinkhamer, P.G.; Leiss, K.A. Light intensity-mediated induction of trichome-associated allelochemicals increases resistance against thrips in tomato. Plant Cell Physiol. 2018, 59, 2462–2475. [Google Scholar] [CrossRef]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. Biosci. J. 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Berry, J.P.; Gantar, M.; Perez, M.H.; Berry, G.; Noriega, F.G. Cyanobacterial toxins as allelochemicals with potential applications as algaecides, herbicides and insecticides. Mar. Drugs 2008, 6, 117–146. [Google Scholar] [CrossRef]
- Beeler, J.E.; Rhodes, G.N.; Bates, G.E.; Main, C.L.; Mueller, T.C. Horsenettle (Solanum carolinense) control in tall fescue (Festuca arundinacea) and clover (Trifolium sp.) pastures with mixtures of 2,4-D and picloram. Camb. Core Weed Technol. 2004, 18, 1091–1095. [Google Scholar] [CrossRef]
- Gorrell, R.M.; Bingham, S.W.; Foy, C.L. Control of Horsenettle (Solarium carolinense) Fleshy Roots in Pastures. Camb. Core Weed Sci. 1981, 29, 586–589. [Google Scholar] [CrossRef]
- Nichols, R.L.; Cardina, J.; Lynch, R.L.; Minton, N.A.; Wells, H.D. Insects, Nematodes, and Pathogens Associated by Horsenettle (Solanum carolinense) in Bermudagrass (Cynodon dactylon) Pastures. Camb. Core Weed Sci. 1992, 40, 320–325. [Google Scholar] [CrossRef]
- Davies, K.W.; Sheley, R.L. A conceptual framework for preventing the spatial dispersal of invasive plants. Camb. Core Weed Sci. 2007, 55, 178–184. [Google Scholar] [CrossRef]
- Milner, S.E.; Brunton, N.P.; Jones, P.W.; O’Brien, N.M.; Collins, S.G.; Maguire, A.R. Bioactivities of glycoalkaloids and their aglycones from Solanum species. J. Agric. Food. Chem. 2011, 59, 3454–3484. [Google Scholar] [CrossRef]
- Yencho, G.C.; Kowalski, S.P.; Kennedy, G.G.; Sanford, L.L. Segregation of leptine glycoalkaloids and resistance to Colorado potato beetle (Leptinotarsa decemlineata (Say)) in F2 Solanum tuberosum (4x) × S. chacoense (4x) potato progenies. Am. J. Potato Res. 2000, 77, 167–178. [Google Scholar] [CrossRef]
- Fukuhara, K.; Shimizu, K.; Kubo, I. Arudonine, an allelopathic steroidal glycoalkaloid from the root bark of Solanum arundo Mattei. Phytochemistry 2004, 65, 1283–1286. [Google Scholar] [CrossRef]
- Sołtys-Kalina, D.; Murawska, Z.; Strzelczyk-Żyta, D.; Wasilewicz-Flis, I.; Marczewski, W. Phytotoxic potential of cultivated and wild potato species (Solanum sp.): Role of glycoalkaloids, phenolics and flavonoids in phytotoxicity against mustard (Sinapis alba L.). Acta Physiol. Plant. 2019, 41, 55. [Google Scholar] [CrossRef]
- Rudrappa, T.; Bonsall, J.; Gallagher, J.L.; Seliskar, D.M.; Bais, H.P. Root-secreted allelochemical in the noxious weed Phragmites australis deploys a reactive oxygen species response and microtubule assembly disruption to execute rhizotoxicity. J. Chem. Ecol. 2007, 33, 1898–1918. [Google Scholar] [CrossRef]
- Coelho, É.M.P.; Barbosa, M.C.; Mito, M.S.; Mantovanelli, G.C.; Oliveira, R.S.; Ishii-Iwamoto, E.L. The activity of the antioxidant defense system of the weed species Senna obtusifolia L. and its resistance to allelochemical stress. J. Chem. Ecol. 2017, 43, 725–738. [Google Scholar] [CrossRef]
- Weir, T.L.; Park, S.-W.; Vivanco, J.M. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant Biol. 2004, 7, 472–479. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Adhikari, A.; Lee, I.-J.; Loake, G.J.; Yun, B.-W. A Novel DUF569 Gene Is a Positive Regulator of the Drought Stress Response in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 5316. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Kazerooni, E.A.; Maharachchikumbura, S.S.; Adhikari, A.; Al-Sadi, A.M.; Kang, S.-M.; Kim, L.-R.; Lee, I.-J. Rhizospheric Bacillus amyloliquefaciens protects Capsicum annuum cv. Geumsugangsan from multiple abiotic stresses via multifarious plant growth-promoting attributes. Front. Plant Sci. 2021, 12, 821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, S.-W.; Shi, H.-L.; Zhao, K.; Wang, J.; Liu, Y.; Liu, X.-H.; Wang, W.J.P. Physiological and biochemical mechanisms mediated by allelochemical isoliquiritigenin on the growth of lettuce seedlings. Plants 2020, 9, 245. [Google Scholar] [CrossRef]
- Ren, X.; Yan, Z.-q.; He, X.-f.; Li, X.-z.; Qin, B. Allelopathic effect of β-cembrenediol and its mode of action: Induced oxidative stress in lettuce seedlings. Emir. J. Food Agric. 2017, 6, 441–449. [Google Scholar] [CrossRef]
- Krasuska, U.; Andrzejczak, O.; Staszek, P.; Borucki, W.; Gniazdowska, A. meta-Tyrosine induces modification of reactive nitrogen species level, protein nitration and nitrosoglutathione reductase in tomato roots. Nitric Oxide 2017, 68, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef]
- Ahmad, P.; Umar, S.; Sharma, S. Mechanism of free radical scavenging and role of phytohormones in plants under abiotic stresses. In Plant Adaptation and Phytoremediation; Springer: Dordrecht, The Netherlands, 2010; pp. 99–118. [Google Scholar] [CrossRef]
- Adhikari, A.; Khan, M.A.; Imran, M.; Lee, K.-E.; Kang, S.-M.; Shin, J.Y.; Joo, G.-J.; Khan, M.; Yun, B.-W.; Lee, I.-J. The Combined Inoculation of Curvularia lunata AR11 and Biochar Stimulates Synthetic Silicon and Potassium Phosphate Use Efficiency, and Mitigates Salt and Drought Stresses in Rice. Front. Plant Sci. 2022, 13, 816858. [Google Scholar] [CrossRef]
- Adhikari, A.; Khan, M.A.; Lee, K.-E.; Kang, S.-M.; Dhungana, S.K.; Bhusal, N.; Lee, I.-J. The Halotolerant Rhizobacterium—Pseudomonas koreensis MU2 Enhances Inorganic Silicon and Phosphorus Use Efficiency and Augments Salt Stress Tolerance in Soybean (Glycine max L.). Microoganisms 2020, 8, 1256. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Wang, B.; Song, G.; Shi, S. Comparative study of alleviation effects of DMTU and PCIB on root growth inhibition in two tall fescue varieties under cadmium stress. Ecotoxicol. Environ. Saf. 2020, 196, 110528. [Google Scholar] [CrossRef]
- Pirnajmedin, F.; Majidi, M.M.; Taleb, H.; Saeidi, G.; Shojaiefar, S. Genotypic-specific response to exogenous applied salicylic acid in tall fescue under different irrigation conditions. Crop Sci. 2020, 60, 1123–1130. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Zhuang, L.; Gao, Y.; Huang, B. Abscisic acid mediation of drought priming-enhanced heat tolerance in tall fescue (Festuca arundinacea) and Arabidopsis. Physiol. Plant. 2019, 167, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Rostami, M.; Rostami, S. Effect of salicylic acid and mycorrhizal symbiosis on improvement of fluoranthene phytoremediation using tall fescue (Festuca arundinacea Schreb). Chemosphere 2019, 232, 70–75. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, Z.; Yang, Y.; Li, M.; Xu, B. Abscisic acid and brassinolide combined application synergistically enhances drought tolerance and photosynthesis of tall fescue under water stress. Sci. Hortic. 2018, 228, 1–9. [Google Scholar] [CrossRef]
- Ervin, E.; Zhang, X.; Schmidt, R. Exogenous Salicylic Acid Enhances Post-Transplant Success of Heated Kentucky Bluegrass and Tall Fescue Sod. Crop Sci. 2005, 45, 240–244. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Y.; Liu, Q.; Zong, B.; Yuan, X.; Sun, H.; Wang, J.; Zang, L.; Ma, Z.; Liu, H.; et al. Nitric oxide is involved in abscisic acid-induced photosynthesis and antioxidant system of tall fescue seedlings response to low-light stress. Environ. Exp. Bot. 2018, 155, 226–238. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 2:1 | 1:1 | 1:2 | |
|---|---|---|---|---|
| Germination rate (%) | 81.7 ± 1.67 a | 65 ± 0 b | 56.7 ± 3.33 c | 50 ± 2.89 c |
| Plant height (cm) | 27.2 ± 0.92 a | 18 ± 1.01 b | 16.4 ± 0.79 b | 15.6 ± 0.91 b |
| Root length (cm) | 15.3 ± 0.88 a | 8.6 ± 0.84 b | 8.3 ± 0.58 b | 10.2 ± 1.05 b |
| Fresh weight (mg) | 1236 ± 174 a | 110 ± 19.5 b | 84 ± 13.3 b | 57 ± 9.9 b |
| Dry weight (mg) | 120 ± 17.51 a | 14.8 ± 2.4 b | 9.5 ± 1.79 b | 9.1 ± 1.58 b |
| Control | 20 DAS | 40 DAS | 60 DAS | |
|---|---|---|---|---|
| Germination rate (%) | 80 ± 0 a | 61.7 ± 4.41 b | 60 ± 2.89 b | 60 ± 2.89 b |
| Plant height (cm) | 22.9 ± 0.88 a | 11.3 ± 0.48 b | 9.8 ± 0.53 b | 8.2 ± 0.39 b |
| Root length (cm) | 12.8 ± 0.54 a | 4.1 ± 0.34 b | 3.9 ± 0.37 b | 3.5 ± 0.44 b |
| Fresh weight (mg) | 586.2 ± 58.7 a | 55.1 ± 6.47 b | 39.8 ± 13.74 b | 17.9 ± 2.61 b |
| Dry weight (mg) | 77.2 ± 11.4 a | 5 ± 0.7 b | 4.3 ± 1.59 b | 2.8 ± 0.37 b |
| Concentration (ppm) | Control | 2500 | 5000 |
|---|---|---|---|
| Plant height (cm) | 32.8 ± 1.21 a | 24.7 ± 1.54 b | 18.6 ± 2.42 c |
| Fresh weight (g) | 1.5 ± 0.06 a | 1.2 ± 0.05 b | 0.9 ± 0.04 c |
| Dry weight (g) | 0.21 ± 0.005 a | 0.18 ± 0.006 b | 0.12 ± 0.005 c |
| No | Name of the Compound | Peak Area (%) |
|---|---|---|
| 1 | Solanidan-3-ol, (3β,5α) | 51.96 |
| 2 | 1H,7H-[1,3]Benzodioxino[6,5-g] [1,3]benzodioxolo[5,6-a]quinolizine, 9,10,15b,16-tetrahydro-5,15-dimethoxy-, (S) | 11.59 |
| 3 | 5,5-Dimethylimidazolidin-2,4-dione | 5.65 |
| 4 | solanidan-3-one | 5.64 |
| 5 | D-Allose | 4.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, L.-R.; Adhikari, A.; Kang, Y.; Gam, H.-J.; Kang, S.-M.; Kim, K.-Y.; Lee, I.-J. Investigation of Solanum carolinense Dominance and Phytotoxic Effect in Festuca arundinacea with Special Reference to Allelochemical Identification, Analysis of Phytohormones and Antioxidant Mechanisms. Agronomy 2022, 12, 1954. https://doi.org/10.3390/agronomy12081954
Kim L-R, Adhikari A, Kang Y, Gam H-J, Kang S-M, Kim K-Y, Lee I-J. Investigation of Solanum carolinense Dominance and Phytotoxic Effect in Festuca arundinacea with Special Reference to Allelochemical Identification, Analysis of Phytohormones and Antioxidant Mechanisms. Agronomy. 2022; 12(8):1954. https://doi.org/10.3390/agronomy12081954
Chicago/Turabian StyleKim, Lee-Rang, Arjun Adhikari, Yosep Kang, Ho-Jun Gam, Sang-Mo Kang, Ki-Yong Kim, and In-Jung Lee. 2022. "Investigation of Solanum carolinense Dominance and Phytotoxic Effect in Festuca arundinacea with Special Reference to Allelochemical Identification, Analysis of Phytohormones and Antioxidant Mechanisms" Agronomy 12, no. 8: 1954. https://doi.org/10.3390/agronomy12081954