
Phenotypic Broad Spectrum of Bacterial Blight Disease Resistance from Thai Indigenous Upland Rice Germplasms Implies Novel Genetic Resource for Breeding Program

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Xanthomonas oryzae pv. oryzae Isolates
2.3. Screening for BB Resistance of Indigenous Upland Rice Germplasms
2.4. Resistance Abilities of Upland Rice to the Five Individual Xoo Isolates under Greenhouse Conditions
2.5. Resistance Abilities of Upland Rice to Five Mixed Xoo Isolates in the Field
2.6. Resistance Ability of Upland Rice to Natural Xoo in the Field
2.7. Data Analysis
3. Results
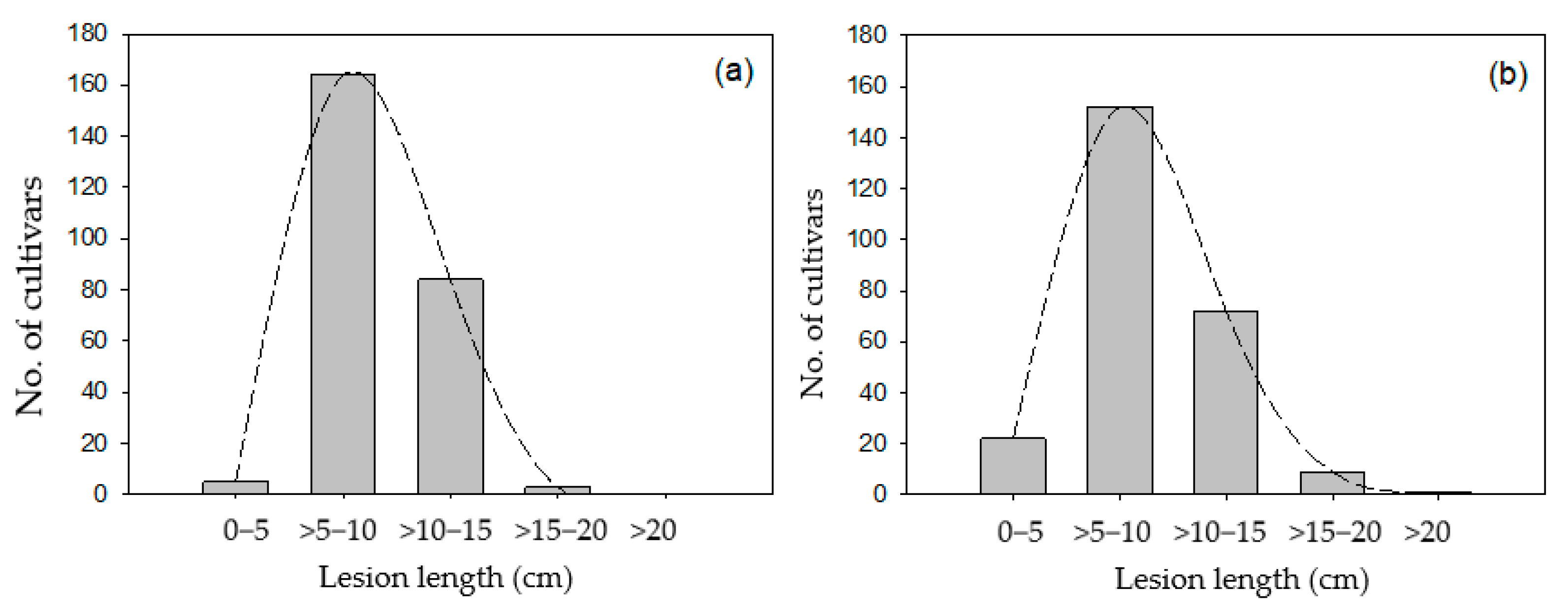
3.1. Screening for BB Resistance of Indigenous Upland Rice Germplasms
3.2. Resistance Abilities of Upland Rice to the Five Individual Xoo Isolates under Greenhouse Conditions
3.3. Resistance Abilities of Upland Rice to Mixed Five Xoo Isolates in the Field
3.4. Resistance Ability of Upland Rice to Natural Xoo in the Field
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.H. Rice Diseases, 2nd ed.; Commonwealth Mycological Institute: Kew, UK, 1985. [Google Scholar]
- Swings, J.; Van Den Mooter, M.; Vauterin, L.; Hoste, B.; Gillis, M.; Mew, T.W.; Kersters, K. Reclassification of the causal agents of bacterial blight (Xanthomonas campestris pv. oryzae) and bacterial leaf streak (Xanthomonas campestris pv. oryzicola) of rice as pathovars of Xanthomonas oryzae). Int. J. Syst. Evol. 1990, 40, 309–311. [Google Scholar] [CrossRef]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Hasan, M.M.; Oladosu, Y.A.; Magaji, U.G.; Akos, I.; Olalekan, K.K. Bacterial leaf blight resistance in rice: A review of conventional breeding to molecular approach. Mol. Biol. Rep. 2019, 46, 1519–1532. [Google Scholar] [CrossRef] [PubMed]
- Mew, T.W. Current status and future prospects of research on bacterial blight of rice. Annu. Rev. Phytopathol. 1987, 25, 359–382. [Google Scholar] [CrossRef]
- Adhikari, T.B.; Mew, T.W.; Leach, J.E. Genotypic and pathotypic diversity in Xanthomonas oryzae pv. oryzae in Nepal. Phytopathology 1999, 89, 687–694. [Google Scholar] [CrossRef]
- Noda Lui, D.; Ronald, P.; Bogdanove, A.J. Pathogen profile Xanthomonus oryzae pathovars: Model pathogen of a model crop. Mol. Plant Pathol. 2001, 7, 303–324. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, J.; Zhang, H.; Ji, G.; Zeng, L.; Li, Y.; Yu, C.; Fernando, W.G.D.; Chen, W. Bacterial Blight Induced Shifts in Endophytic Microbiome of Rice Leaves and the Enrichment of Specific Bacterial Strains With Pathogen Antagonism. Front. Plant Sci. 2020, 11, 963. [Google Scholar] [CrossRef]
- Gupta, P.C.; O’Toole, J.C. Upland Rice: A Global Perspective; International Rice Research Institute: Manila, Philippines, 1986; pp. 1–14. [Google Scholar]
- Jaruchai, W.; Monkham, T.; Chankaew, S.; Suriharn, B.; Sanitchon, J. Evaluation of stability and yield potential of upland rice genotypes in north and northeast Thailand. J. Integr. Agric. 2018, 17, 28–36. [Google Scholar] [CrossRef]
- Srichant, N.; Chankaew, S.; Monkham, T.; Thammabenjapone, P.; Sanitchon, J. Development of Sakon Nakhon Rice Variety for Blast Resistance through Marker Assisted Backcross Breeding. Agronomy 2019, 9, 67. [Google Scholar] [CrossRef]
- Kwanwah, M.R.; Wongsa, T.; Monkham, T.; Chankaew, S.; Falab, S.; Sanitchon, J. Thai Indigenous Lowland Rice Germplasms: Sources of Bacterial Blight Disease Resistance and Agronomic Attributes. Agrivita 2020, 42, 367–380. [Google Scholar] [CrossRef]
- Rao, K.K.; Lakshminarasu, M.; Jena, K.K. DNA markers and marker-assisted breeding for durable resistance to bacterial blight disease in rice. Biotechnol. Adv. 2002, 20, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.-P.; Jeung, J.-U.; Noh, T.-H.; Cho, Y.-C.; Park, S.-H.; Park, H.-S.; Shin, M.-S.; Kim, C.-K.; Jena, K.K. Development of breeding lines with three pyramided resistance genes that confer broad-spectrum bacterial blight resistance and their molecular analysis in rice. Rice 2013, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Rao, G.J.N.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef] [PubMed]
- Yugander, A.; Sundaram, R.M.; Singh, K.; Ladhalakshmi, D.; Rao, L.V.S.; Madhav, M.S.; Badri, J.; Prasad, M.S.; Laha, G.S. Incorporation of the novel bacterial blight resistance gene Xa38 into the genetic background of elite rice variety Improved Samba Mahsuri. PLoS ONE 2018, 13, e0198260. [Google Scholar] [CrossRef]
- Aung Nan, M.S.; Janto, J.; Sribunrueang, A.; Chankaew, S.; Monkham, T.; Sanitchon, J. Field Evaluation of RD6 Introgression Lines for Yield Performance, Blast, and Bacterial Blight Resistance; and their Cooking and Eating Qualities. Agronomy 2019, 9, 825. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Chiu, C.H.; Yap, R.; Tseng, Y.C.; Wu, Y.P. Pyramiding bacterial blight resistance genes in Tainung82 for broad-spectrum resistance using marker-assisted selection. Int. J. Mol. Sci. 2020, 21, 1281. [Google Scholar] [CrossRef]
- White, F.F.; Yang, B. Host and pathogen factors controlling the rice-Xanthomonas oryzae interaction. Plant Physiol. 2009, 150, 1677–1686. [Google Scholar] [CrossRef]
- Ryan, R.; Vorhölter, F.-J.; Potnis, N.; Jones, J.B.; Van Sluys, M.-A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding bacterium–plant interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef]
- Pandey, M.; Rani, N.S.; Sundaram, R.M.; Laha, G.S.; Madhav, M.S.; Rao, K.S.; Sudharshan, I.; Hari, Y.; Varaprasad, G.S.; Rao, L.V.S.; et al. Improvement of two traditional Basmati rice varieties for bacterial blight resistance and plant stature through morphological and marker-assisted selection. Mol. Breed. 2013, 31, 239–246. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Jiang, G.; Xu, Y.; He, Y. Pyramiding of Xa7 and Xa21 for the improvement of disease resistance to bacterial blight in hybrid rice. Plant Breed. 2006, 125, 600–605. [Google Scholar] [CrossRef]
- Perumalsamy, S.; Bharani, M.; Sudha, M.; Nagarajan, P.; Arul, L.; Saraswathi, R.; Balasubramanian, P.; Ramalingam, J. Functional marker-assisted selection for bacterial leaf blight resistance genes in rice (Oryza sativa L.). Plant Breed. 2010, 129, 400–406. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.R.; Biradar, S.; Laha, G.S.; Reddy, G.A.; Rani, N.S.; Sharma, N.P.; Sonti, R.V. Marker assisted introgression of bacterial blight resistance in Samba Mahsuri, an elite indica rice variety. Euphytica 2008, 160, 411–422. [Google Scholar] [CrossRef]
- Kauffman, H.E.; Reddy, A.P.K.; Hsieh, S.P.Y.; Merca, S.D. An improved technique for evaluation of resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- Sribunrueang, A.; Chankaew, S.; Thummabenjapone, P.; Sanitchon, J. Stability of four new sources of bacterial leaf blight resistance in Thailand obtained from indigenous rice varieties. Agrivita 2017, 39, 128–136. [Google Scholar] [CrossRef]
- IRRI. Standard Evaluation System for Rice (SES); International Rice Research Institute: Manila, Philippines, 2013. [Google Scholar]
- Ahn, S.W. International collaboration on breeding for resistance to rice blast. In Rice Blast Disease; Zeigler, R.S., Leong, S.A., Teng, P.S., Eds.; CAB International: Wallingford, UK, 1994; pp. 137–153. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research, 2nd ed.; John Wiley and Sons: New York, NY, USA, 1984; 680p. [Google Scholar]
- Abdul Fiyaz, R.; Shivani, D.; Chaithanya, K.; Mounika, K.; Chiranjeevi, M.; Laha, G.S.; Viraktamath, B.C.; Subba Rao, L.V.; Sundaram, R.M. Genetic Improvement of Rice for Bacterial Blight Resistance: Present Status and Future Prospects. Rice Sci. 2022, 29, 118–132. [Google Scholar] [CrossRef]
- Pinta, W.; Toojinda, T.; Thummabenjapone, P.; Sanitchon, J. Pyramiding of blast and bacterial leaf blight resistance genes into rice cultivar RD6 using marker assisted selection. Afr. J. Biotechnol. 2013, 12, 4432–4438. [Google Scholar] [CrossRef]
- Wongkhamchan, A.; Chankaew, S.; Monkham, T.; Saksirirat, W.; Sanitchon, J. Breeding Thai rice variety RD6 with xa5 gene from IR62266 for broad resistance to bacterial leaf blight in Northeastern Thailand. Agric. Nat. Resour. 2018, 52, 241–245. [Google Scholar] [CrossRef]
- Sundaram, R.M.; Vishnupriya, M.R.; Laha, G.S.; Rani, N.S.; Rao, P.S.; Balachandran, S.M.; Reddy, G.A.; Sarma, N.P.; Sonti, R.V. Introduction of bacterial blight resistance into Triguna, a high yielding, mid-early duration rice variety. Biotechnol. J. 2009, 4, 400–407. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, H.B.; Zhou, Y.; Zhang, Q.L.; Li, X.H.; Wang, S.P. Exploring the mechanism and efficient use of a durable gene mediated resistance to bacterial blight disease in rice. Mol Breed. 2018, 38, 18. [Google Scholar] [CrossRef]
- Win, K.M.; Korinsak, S.; Jantaboon, J.; Siangliw, M.; Lanceras-Siangliw, J.; Sirithunya, P.; Toojinda, T. Breeding the Thai jasmine rice variety KDML105 for non-age-related broad-spectrum resistance to bacterial blight disease based on combined marker-assisted and phenotypic selection. Field Crops Res. 2012, 137, 186–194. [Google Scholar] [CrossRef]
- Sombunjitt, S.; Sriwongchai, T.; Kuleung, C.; Hongtrakul, V. Searching for and analysis of bacterial blight resistance genes from Thailand rice germplasm. Agric. Nat. Resour. 2017, 51, 365–375. [Google Scholar] [CrossRef]
- Ji, Z.; Wang, C.; Zhao, K. Rice routes of countering Xanthomonas oryzae. Int. J. Mol. Sci. 2018, 19, 3008. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Xiao, S.; Li, D.; Chen, L.; Zhong, Q.; Yin, F.; Yu, T.; Ke, X.; Zhang, D.; Fu, J.; et al. Identification and expression pattern analysis of bacterial blight resistance genes in Oryza officinalis Wall ex Watt Under Xanthomonas oryzae pv. oryzae Stress. Plant Mol. Biol. Rep. 2019, 37, 436–449. [Google Scholar] [CrossRef]
- Yu, X.Y.; Liang, X.Y.; Liu, K.X.; Dong, W.X.; Wang, J.X.; Zhou, M.G. The thiG gene is required for full virulence of Xanthomonas oryzae pv. oryzae by preventing cell aggregation. PLoS ONE 2015, 10, e0134237. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.H.; Ji, Z.Y.; Liu, B.; Cheng, H.; Liu, H.; Liu, S.Z.; Yang, B.; Chen, G.Y. Xa1 allelic R genes activate rice blight resistance suppressed by interfering TAL efectors. Plant Commun. 2020, 1, 100087. [Google Scholar] [CrossRef]
- Anderson, J.P.; Gleason, C.A.; Foley, R.C.; Thrall, P.H.; Burdon, J.B.; Singh, K.B. Plants versus pathogens: An evolutionary arms race. Funct. Plant Biol. 2010, 37, 499–512. [Google Scholar] [CrossRef]
- Banerjee, A.; Roy, S.; Kumar Bag, M.; Bhagat, S.; Kumar Kar, M.; Mandal, N.P.; Mukherjee, A.K.; Maiti, D. A survey of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice germplasm from eastern and northeastern India using molecular markers. Crop. Prot. 2018, 112, 168–176. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Keller, B. Resistance: Double gain with one gene. Nat. Plants 2017, 3, 17019. [Google Scholar] [CrossRef]
- Zhao, C.; Yin, F.; Chen, L.; Li, D.; Xiao, S.; Zhong, Q.; Wang, B.; Ke, X.; Fu, J.; Li, X.; et al. Identification of bacterial blight resistance genes in rice landraces from Yunnan Province, China. Australas. Plant Pathol. 2022, 51, 59–69. [Google Scholar] [CrossRef]
- Zhang, F.; Zhuo, D.L.; Zhang, F.; Huang, L.Y.; Wang, W.S.; Xu, J.L.; Vera Cruz, C.; Li, Z.K.; Zhou, Y.L. Xa39, a novel dominant gene conferring broad-spectrum resistance to Xanthomonas oryzae pv. oryzae in rice. Plant Pathol. 2015, 64, 568–575. [Google Scholar] [CrossRef]
- Sontornkarun, T.; Chankaew, S.; Sanitchon, J. Donor parental determination for breeding the RD41 rice cultivar to improve bacterial blight resistance. Khon Kaen Agric. J. 2020, 48, 1162–1171. [Google Scholar] [CrossRef]
- Mishra, D.; Vishnupriya, M.R.; Anil, M.G.; Konda, K.; Raj, Y.; Sonti, R.V. Pathotype and Genetic Diversity amongst Indian Isolates of Xanthomonas oryzae pv. oryzae. PLoS ONE 2013, 8, e81996. [Google Scholar] [CrossRef]
- Rajpurohit, D.; Kumar, R.; Kumar, M.; Paul, P.; Awasthi, A.; Basha, P.O.; Puri, A.; Jhang, T.; Singh, K.; Dhaliwan, H.S. Pyramiding of two bacterial blight resistance and a semi dwarfing gene in Type 3 Basmati using marker-assisted selection. Euphytica 2010, 178, 111–126. [Google Scholar] [CrossRef]
- Luo, Y.; Sangha, J.S.; Wang, S.; Li, Z.F.; Yang, J.B.; Yin, Z.C. Marker-assisted breeding of Xa4, Xa21 and Xa27 in the restorer lines of hybrid rice for broad-spectrum and enhanced disease resistance to bacterial blight. Mol. Breed. 2012, 30, 1601–1610. [Google Scholar] [CrossRef]
- Xing, J.; Zhang, D.; Yin, F.; Zhong, Q.; Wang, B. Identification and Fine-Mapping of a New Bacterial Blight Resistance Gene, Xa47(t), in G252, an Introgression Line of Yuanjiang Common Wild Rice (Oryza rufipogon). Plant Dis. 2021, 105, 4106–4112. [Google Scholar] [CrossRef]
- Sun, X.L.; Cao, Y.L.; Yang, Z.F.; Xu, C.G.; Li, X.H.; Wang, S.P.; Zhang, Q.F. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J. 2004, 37, 517–527. [Google Scholar] [CrossRef]
- Wang, C.L.; Tan, M.P.; Xu, X.; Wen, G.S.; Zhang, D.P.; Lin, X.H. Localizing the Bacterial Blight Resistance Gene, Xa22(t), to a 100-Kilobase Bacterial Artifcial Chromosome. Phytopathology 2003, 93, 1258–1262. [Google Scholar] [CrossRef]
- Wang, C.L.; Wen, G.S.; Lin, X.H.; Liu, X.Q.; Zhang, D.P. Identification and fine mapping of the new bacterial blight resistance gene, Xa31(t), in rice. Eur. J. Plant Pathol. 2009, 123, 235–240. [Google Scholar] [CrossRef]
- Li, Z.; Sanchez, A.; Angeles, E.; Singh, S.; Domingo, J.; Huang, N.; Khush, G.S. Are the Dominant and Recessive Plant Disease Resistance Genes Similar?: A Case Study of Rice R Genes and Xanthomonas oryzae pv. oryzae Races. Genetics 2001, 159, 757–765. [Google Scholar] [CrossRef]
- Laine, A.L. Resistance variation within and among host populations in a plant-pathogen metapopulation: Implications for regional pathogen dynamics. J. Ecol. 2004, 92, 990–1000. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, N.; Koss, M.J.; Greco, C.; Nickles, G.; Wiemann, P.; Keller, N.P. Secreted Secondary Metabolites Reduce Bacterial Wilt Severity of Tomato in Bacterial–Fungal Co-Infections. Microorganisms 2021, 9, 2123. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.A.H. Bacterial leaf blight of rice: An overview of epidemiology and management with special reference to Indian sub-continent. Pak. J. Agri. Res. 2019, 32, 359–380. [Google Scholar] [CrossRef]
- Webb, K.M.; Oña, I.; Bai, J.; Garrett, K.A.; Mew, T.; Vera Cruz, C.M.; Leach, J.E. A benefit of high temperature: Increased effectiveness of a rice bacterial blight resistance gene. New Phytol. 2010, 185, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L. Bacterial blight. Rice Today 2013, 12, 12–13. [Google Scholar]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier-Academic Press: San Diego, CA, USA, 2005. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Cultivars | Mean Lesion Length (cm) | |||||
|---|---|---|---|---|---|---|
| Isolate NB7-8 | Isolate CM3-1 | Isolate CN2-1 | Isolate MS1-2 | Isolate NY1-1 | BSR (%) | |
| ULR024 | 4.97 def | 4.50 e–h | 5.79 efg | 5.73 e–h | 3.92 f–i | 100 |
| ULR029 | 3.58 ef | 2.05 fgh | 5.61 fg | 4.51 fgh | 8.92 c–g | 100 |
| ULR042 | 6.58 c–f | 8.00 a–f | 12.17 a–e | 6.83 d–h | 7.17 e–i | 80 |
| ULR048 | 7.28 c–f | 5.61 d–h | 9.45 b–f | 6.56 d–h | 9.33 b–g | 100 |
| ULR092 | 6.48 c–f | 7.67 a–g | 10.72 a–f | 8.79 b–g | 10.17 b–f | 60 |
| ULR119 | 4.71 def | 5.08 e–h | 6.78 d–g | 4.63 fgh | 8.11 d–h | 100 |
| ULR172 | 3.63 ef | 2.72 fgh | 7.00 d–g | 3.42 fgh | 5.89 f–i | 100 |
| ULR174 | 9.39 a–e | 7.61 a–g | 10.97 a–f | 8.09 b–h | 9.39 b–g | 80 |
| ULR181 | 1.51 f | 4.08 fgh | 4.56 fg | 5.25 fgh | 3.28 f–i | 100 |
| ULR183 | 6.42 c–f | 3.92 fgh | 6.59 d–g | 4.07 fgh | 4.48 f–i | 100 |
| ULR186 | 7.75 c–f | 5.06 e–h | 8.22 c–f | 8.67 b–g | 7.67 d–i | 100 |
| ULR207 | 2.82 ef | 1.75 fgh | 4.75 fg | 3.17 gh | 5.39 f–i | 100 |
| ULR222 | 9.97 a–e | 7.22 b–g | 10.53 a–f | 8.09 c–h | 9.39 b–g | 80 |
| ULR244 | 4.00 def | 5.78 c–h | 7.06 d–g | 6.32 d–h | 4.13 f–i | 100 |
| ULR292 | 4.42 def | 4.50 e–h | 8.00 c–f | 5.08 fgh | 7.00 e–i | 100 |
| ULR296 | 3.00 ef | 3.55 fgh | 7.00 d–g | 5.95 e–h | 6.94 e–i | 100 |
| ULR305 | 3.96 def | 3.67 fgh | 7.58 c–g | 6.11 d–h | 7.36 e–i | 100 |
| ULR337 | 6.65 c–f | 4.89 e–h | 6.83 d–g | 7.17 d–h | 5.61 f–i | 100 |
| ULR356 | 2.99 ef | 2.13 fgh | 6.36 d–g | 7.00 d–h | 7.06 e–i | 100 |
| ULR014 | 12.78 abc | 14.22 a | 16.91 a | 13.05 a–d | 17.11 a | 0 |
| ULR089 | 12.61 abc | 11.06 a–e | 13.89 abc | 16.89 a | 14.56 a–d | 0 |
| RD6 | 8.98 b–e | 12.50 abc | 9.93 b–f | 12.71 a–e | 13.34 a–e | 40 |
| SKN | 16.15 ab | 11.89 a–d | 15.61 ab | 15.12 ab | 15.03 abc | 0 |
| KDML105 | 16.22 a | 13.56 ab | 15.78 ab | 14.94 abc | 15.95 ab | 0 |
| RD23 | 11.03 a–d | 4.67 e–h | 12.50 a–d | 10.33 a–f | 5.08 f–i | 40 |
| IR62266 | 3.04 ef | 1.11 gh | 6.11 d–g | 6.58 d–h | 1.43 hi | 100 |
| IRBB21 | 8.87 cde | 6.22 c–h | 7.70 c–g | 6.67 d–h | 6.94 e–i | 100 |
| IRBB5 | 1.37 f | 0.42 h | 1.33 g | 1.15 h | 0.75 i | 100 |
| SR1 | 7.95 c–f | 3.06 fgh | 12.11 a–e | 9.69 b–g | 3.00 ghi | 80 |
| F-test | ** | ** | ** | ** | ** | - |
| CV (%) | 33.53 | 36.29 | 23.07 | 28.42 | 28.29 | - |
| Cultivars | Mean Score of Bacterial Blight Resistance | |||
|---|---|---|---|---|
| Mixed Isolate Inoculation | Reaction | Natural Infection | Reaction | |
| ULR024 | 3.33 ij | R | 0.00 d | R |
| ULR029 | 4.67 d–i | MR | 1.70 d | R |
| ULR042 | 8.67 ab | S | 8.33 ab | S |
| ULR048 | 8.67 ab | S | 6.67 ab | S |
| ULR092 | 8.83 a | S | 7.33 ab | S |
| ULR119 | 5.17 c–i | MS | 0.37 d | R |
| ULR172 | 4.00 g–j | MR | 0.00 d | R |
| ULR174 | 7.67 a–d | S | 8.17 ab | S |
| ULR181 | 5.67 b–h | MS | 0.27 d | R |
| ULR183 | 7.00 a–g | S | 2.50 cd | R |
| ULR186 | 7.67 a–d | S | 5.83 abc | MS |
| ULR207 | 4.50 e–i | MR | 0.00 d | R |
| ULR222 | 8.00 abc | S | 6.07 ab | MS |
| ULR244 | 6.17 a–i | MS | 2.10 d | R |
| ULR292 | 7.50 a–e | S | 0.80 d | R |
| ULR296 | 7.17 a–f | S | 0.00 d | R |
| ULR305 | 6.67 a–h | MS | 0.27 d | R |
| ULR337 | 4.17 f–i | MR | 1.03 d | R |
| ULR356 | 3.83 hij | MR | 0.67 d | R |
| ULR014 | 9.00 a | S | 8.33 ab | S |
| ULR089 | 8.83 a | S | 8.00 ab | S |
| KDML105 | 9.00 a | S | 7.83 ab | S |
| RD23 | 9.00 a | S | 8.67 ab | S |
| RD6 | 9.00 a | S | 8.50 ab | S |
| SKN | 9.00 a | S | 9.00 a | S |
| IR62266 | 6.00 a–i | MS | 6.17 ab | MS |
| IRBB21 | 8.83 a | S | 6.67 ab | MS |
| IRBB5 | 1.00 j | R | 0.00 d | R |
| SR1 | 8.67 ab | S | 5.50 bc | MS |
| F-test | ** | - | ** | - |
| CV (%) | 14.46 | - | 25.38 | - |
| r | 0.775 ** | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chumpol, A.; Monkham, T.; Saepaisan, S.; Sanitchon, J.; Falab, S.; Chankaew, S. Phenotypic Broad Spectrum of Bacterial Blight Disease Resistance from Thai Indigenous Upland Rice Germplasms Implies Novel Genetic Resource for Breeding Program. Agronomy 2022, 12, 1930. https://doi.org/10.3390/agronomy12081930
Chumpol A, Monkham T, Saepaisan S, Sanitchon J, Falab S, Chankaew S. Phenotypic Broad Spectrum of Bacterial Blight Disease Resistance from Thai Indigenous Upland Rice Germplasms Implies Novel Genetic Resource for Breeding Program. Agronomy. 2022; 12(8):1930. https://doi.org/10.3390/agronomy12081930
Chicago/Turabian StyleChumpol, Atitaya, Tidarat Monkham, Suwita Saepaisan, Jirawat Sanitchon, Shanerin Falab, and Sompong Chankaew. 2022. "Phenotypic Broad Spectrum of Bacterial Blight Disease Resistance from Thai Indigenous Upland Rice Germplasms Implies Novel Genetic Resource for Breeding Program" Agronomy 12, no. 8: 1930. https://doi.org/10.3390/agronomy12081930