
The Impact of Cell Wall Feruloylation on Plant Growth, Responses to Environmental Stress, Plant Pathogens and Cell Wall Degradability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Importance of Cell Walls to Plants
3. Cell Wall Structure
3.1. Cellulose
3.2. Hemicellulose
3.3. Pectin
3.4. Lignin
4. Plant Cell Walls as Food, Feed and a Source of Second-Generation Biofuels
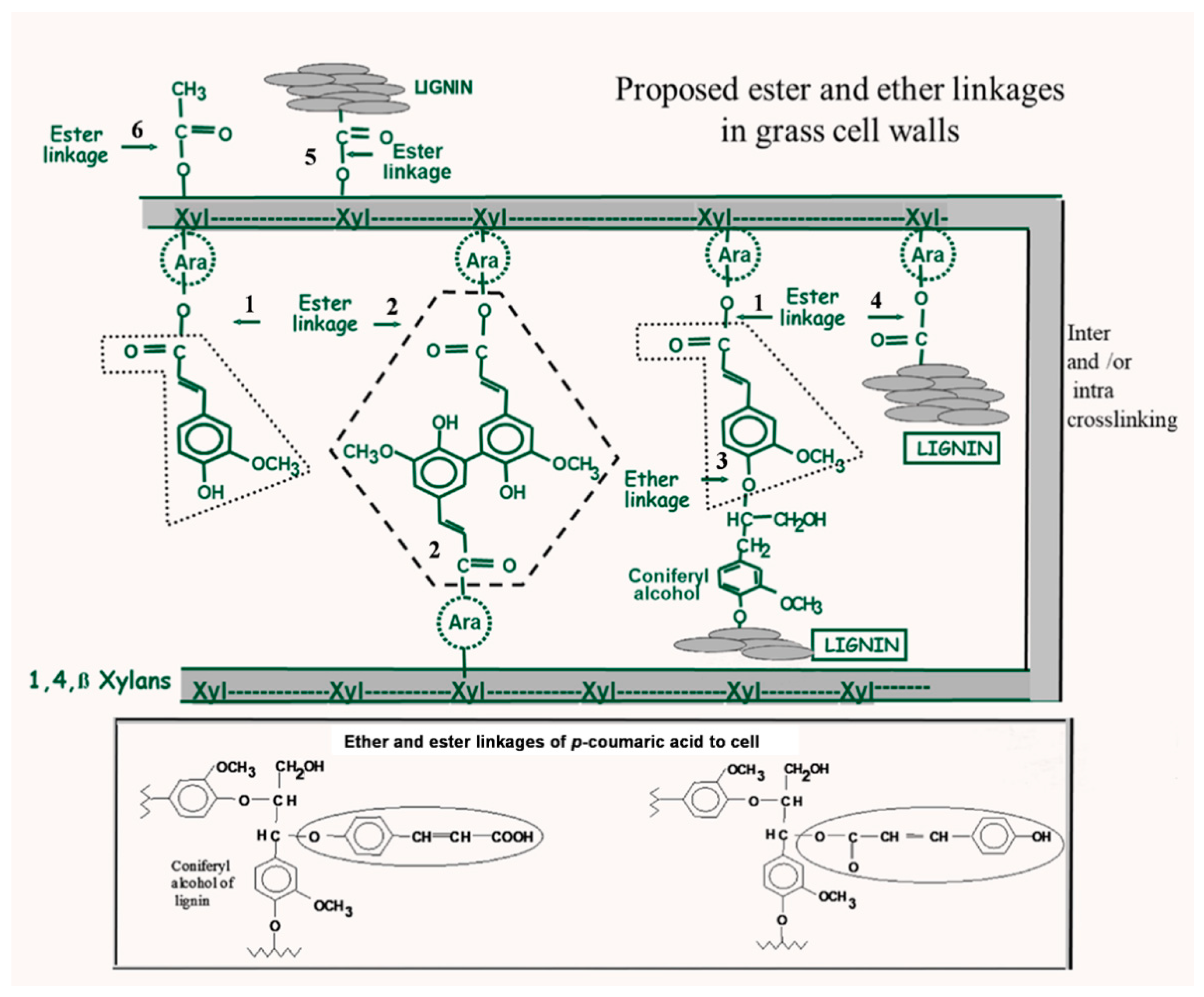
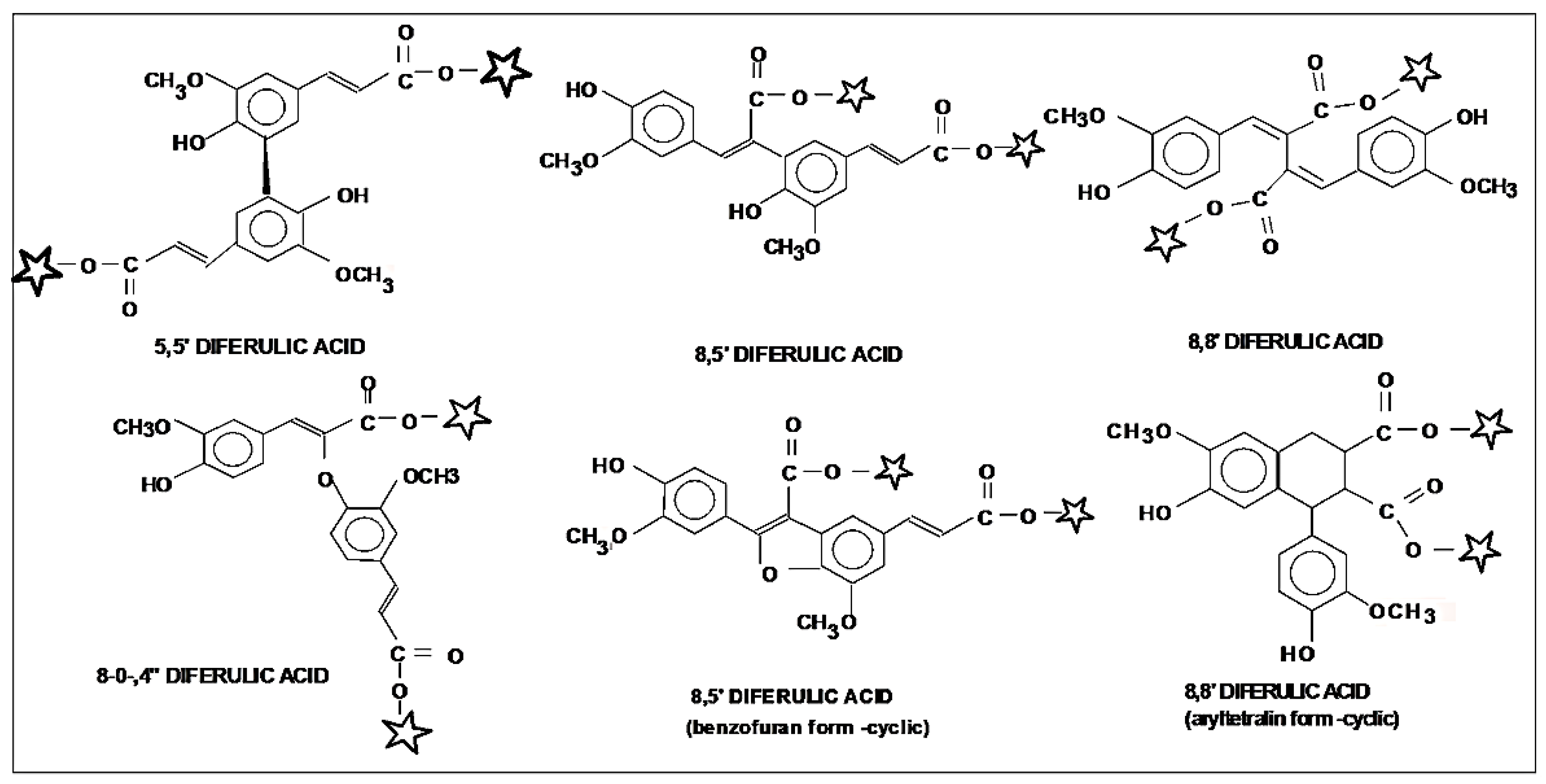
5. Cell Wall Feruloylation and Cross-Linking in Relation to Lignification
6. Strategies for Manipulation of Feruloylation
6.1. Conventional Plant Breeding
6.1.1. Divergent Selection
6.1.2. Association and QTL Mapping
6.1.3. Naturally Occurring and Induced Mutants
Ferulate Ester Mutants
Brown Mid-Rib Mutants
6.2. Genetic Modification of Structural or Regulatory Gene Expression
6.2.1. Pathway Genes
6.2.2. Transcription Factors
6.3. Expression of Transgenes That Alter Feruloylation
6.3.1. Microbial Ferulic Acid Esterases (FAE)
6.3.2. Other Cell-Wall-Degrading Enzymes Expressed in Plants Affecting Cell Wall Feruloylation
6.3.3. Genetic Manipulation of Lignin Genes
7. The Role of Cell Wall Feruloylation
7.1. Feruloylation Contributes to Cell-to-Cell Adhesion
7.2. Feruloylation Contributes to Cell Wall Strength and Growth Cessation
7.3. Feruloylation Contributes to Resistance to Pathogens and Herbivores
7.4. Feruloylation Is Modified by Abiotic Stress
7.5. Feruloylation Contributes to Cell Wall Degradability
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, M.M.; Hartley, R.D. Occurrence and nature of ferulic acid substitution of cell-wall polysaccharides in graminaceous plants. Carbohydr. Res. 1983, 118, 65–80. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Ralph, J.; Grabber, J.H. Cell wall cross-linking by ferulates and diferulates in grasses. J. Sci. Food Agric. 1999, 79, 403–407. [Google Scholar] [CrossRef]
- Bunzel, M.; Ralph, J.; Kim, H.; Lu, F.; Ralph, S.A.; Marita, J.M.; Hatfield, R.D.; Steinhart, H. Sinapate dehydrodimers and sinapate—Ferulate heterodimers in cereal dietary fiber. J. Agric. Food Chem. 2003, 51, 1427–1434. [Google Scholar] [CrossRef]
- Fry, S.C. Feruloylated pectins from the primary cell wall: Their structure and functions. Planta 1983, 157, 111–123. [Google Scholar] [CrossRef]
- Ishii, T.; Tobita, T. Structural characterization of feruloyl oligosaccharides from spinach-leaf cell walls. Carbohydr. Res. 1993, 248, 179–190. [Google Scholar] [CrossRef]
- Wallace, G.; Fry, S.C. In-vitro peroxidase-catalyzed oxidation of ferulic acid esters. Phytochemistry 1995, 39, 1293–1299. [Google Scholar] [CrossRef]
- Ishii, T. Structure and functions of feruloylated polysaccharides. Plant Sci. 1997, 127, 111–117. [Google Scholar] [CrossRef]
- Jacquet, G.; Pollet, B.; Lapierre, C.; Mhamdi, F.; Rolando, C. New ether-linked ferulic acid-coniferyl alcohol dimers identified in grass straws. J. Agric. Food Chem. 1995, 43, 2746–2751. [Google Scholar] [CrossRef]
- Williamson, G.; Kroon, P.A.; Faulds, C.B. Hairy plant polysaccharides: A close shave with microbial esterases. Microbiology 1998, 144, 2011–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, M.R.; Lou, H.; Aubert, M.K.; Wilkinson, L.G.; Little, A.; Houston, K.; Pinto, S.C.; Shirley, N.J. Exploring the role of cell wall-related genes and polysaccharides during plant development. Plants 2018, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamant, O.; Trass, J. The mechanics behind plant development. New Phytol. 2010, 185, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Bidhendi, A.J.; Geitmann, A. Relating the mechanics of the primary plant cell wall to morphogenesis. J. Exp. Bot. 2016, 67, 449–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamant, O.; Haswell, E.S. Life behind the wall: Sensing mechanical cues in plants. BMC Biol. 2017, 15, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldron, W.; Brett, C.T. The role of polymer cross-linking in intercellular adhesion. In Plant Cell Separation and Adhesion; Blackwell Publishing Oxford: Hoboken, NJ, USA, 2007; pp. 183–204. [Google Scholar]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacete, L.; Melida, H.; Miedes, E.; Molina, A. Plant cell wall mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; He, M.; Hou, Q.; Zou, L.; Wei, Y.; Chen, X. Cell wall associated immunity in plants. Stress Biol. 2021, 1, 3. [Google Scholar] [CrossRef]
- Houston, K.; Tucker, M.R.; Chowdhury, J.; Shirley, N.; Little, A. The plant cell wall: A complex and dynamic structure as revealed by the responses of genes under stress conditions. Front. Plant Sci. 2016, 7, 984. [Google Scholar] [CrossRef] [Green Version]
- Taylor, N.G. Cellulose biosynthesis and deposition in higher plants. New Phytol. 2008, 178, 239–252. [Google Scholar] [CrossRef]
- Harris, P.H. Primary and secondary plant cell walls: A comparative overview. N. Z. J. For. Sci. 2006, 36, 36–53. [Google Scholar]
- Krässig, H.A. Effect of structure and morphology on accessibility and reactivity. In Cellulose: Structure, Accessibility, and Reactivity, Polymer Monographs; Gordon and Breach Science Publishers CRS Press: Philadelphia, PA, USA, 1993; pp. 167–324. [Google Scholar]
- Harris, P.J.; Stone, B.A. Chemistry, and molecular organization of plant cell walls. In Biomass Recalcitrance: Deconstructing the Plant Cell Wall for Bioenergy; Himmel, M.E., Ed.; Blackwell Publishing: Oxford, UK, 2009; pp. 61–93. [Google Scholar]
- Delmer, D.P. Cellulose biosynthesis: Exciting times for a difficult field of study. Ann. Rev. Plant Biol. 1999, 50, 245–276. [Google Scholar] [CrossRef]
- Lampugnani, E.R.; Flores-Sandoval, E.; Tan, Q.W.; Mutwil, M.; Bowman, J.L.; Persson, S. Cellulose synthesis–central components and their evolutionary relationships. Trends Plant Sci. 2019, 24, 402–412. [Google Scholar] [CrossRef]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Ann. Rev. Plant Biol. 2010, 2, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C.; McCann, M.C. Redesigning plant cell walls for the biomass-based bioeconomy. J. Biol. Chem. 2020, 295, 15144–15157. [Google Scholar] [CrossRef] [PubMed]
- Gomez, L.D.; Steele-King, C.G.; McQueen-Mason, S.J. Sustainable liquid biofuels from biomass: The writing’s on the walls. New Phytol. 2008, 178, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Driouich, A.; Chevalier, L.; Vicré, M.; Lerouxel, O. Golgi-mediated synthesis and secretion of matrix polysaccharides of the primary cell wall of higher plants. Front. Plant Sci. 2012, 30, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grantham, N.J.; Wurman-Rodrich, J.; Terrett, O.M.; Lyczakowski, J.J.; Stott, K.; Iuga, D.; Simmons, T.J.; Durand-Tardif, M.; Brown, S.P.; Dupree, R.; et al. An even pattern of xylan substitution is critical for interaction with cellulose in plant cell walls. Nat. Plants 2017, 3, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Carpita, N.C. Structure and biogenesis of the cell walls of grasses. Ann. Rev. Plant Biol. 1996, 47, 445–476. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef]
- Ridley, B.L.; O’Neill, M.A.; Mohnen, D. Pectins: Structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 2001, 57, 929–967. [Google Scholar] [CrossRef]
- Liners, F.; Letesson, J.J.; Didembourg, C.; Van Cutsem, P. Monoclonal antibodies against pectin: Recognition of a conformation induced by calcium. Plant Physiol. 1989, 91, 1419–1424. [Google Scholar] [CrossRef]
- Ralph, J.; Lundquist, K.; Brunow, G.; Lu, F.; Kim, H.; Schatz, P.F.; Marita, J.M.; Hatfield, R.D.; Ralph, S.A.; Christensen, J.H.; et al. Lignins: Natural polymers from oxidative coupling of 4-hydroxyphenyl-propanoids. Phytochem. Rev. 2004, 3, 29–60. [Google Scholar] [CrossRef]
- Ralph, J.; Lapierre, C.; Boerjan, W. Lignin structure and its engineering. Curr. Opin. Biotech. 2019, 56, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.M.; Sederoff, R.R. Variation in lignin content and composition (mechanisms of control and implications for the genetic improvement of plants). Plant Physiol. 1996, 110, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Terashima, N. Formation, and structure of lignin in monocotyledons. IV. Deposition process and structural diversity of the lignin in the cell wall of sugarcane and rice plant studies by ultraviolet microscopic spectrometry. Holzforschung 1991, 45, 191–198. [Google Scholar] [CrossRef]
- Lu, F.; Ralph, J. Detection, and determination of p-coumaroylated units in lignin. J. Agric. Food Chem. 1999, 47, 1988–1992. [Google Scholar] [CrossRef]
- Alder, E. Lignin chemistry—Past, present, and future. Wood Sci. Technol. 1977, 11, 169–218. [Google Scholar] [CrossRef]
- Jones, L.; Ennos, A.R.; Turner, S.R. Cloning and characterization of irregular xylem4 (irx4): A severely lignin-deficient mutant of Arabidopsis. Plant J. 2001, 26, 205–216. [Google Scholar] [CrossRef]
- Sattler, S.; Funnell-Harris, D. Modifying lignin to improve bioenergy feedstocks: Strengthening the barrier against pathogens? Front. Plant Sci. 2013, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, N.H.; Selvaraj, G.; Wei, Y.; King, J. Gene expression profiling and silencing reveal that monolignol biosynthesis plays a critical role in penetration defense in wheat against powdery mildew invasion. J. Exper. Bot. 2009, 60, 509–521. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Jung, H.-J.G. Lignin, and fiber digestion. J. Range Manag. 2001, 54, 420–430. [Google Scholar] [CrossRef]
- Moore, K. J.; Hatfield, R. D. Carbohydrates, and forage quality. In Forage Quality, Evaluation, and Utilization; Fahey, G.C., Jr., Collins, M.C., Mertens, D.R., Moser, L.E., Eds.; ASA-CSSA-SSSA: Madison, WI, USA, 1994; pp. 229–280. [Google Scholar]
- Padayachee, A.; Day, L.; Howell, K.; Gidley, M.J. Complexity, and health functionality of plant cell wall fibers from fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2017, 57, 59–81. [Google Scholar] [CrossRef] [PubMed]
- Hoad, C.L.; Rayment, P.; Spiller, R.C.; Marciani, L.; Alonso, B.d.C.; Traynor, C.; Mela, D.J.; Peters, H.P.F.; Gowland, P.A. In vivo imaging of intragastric gelation and its effect on satiety in humans. J. Nutr. 2004, 134, 2293–2300. [Google Scholar] [CrossRef] [Green Version]
- Othman, R.A.; Moghadasian, M.H.; Jones, P.J. Cholesterol-lowering effects of oat β-glucan. Nutr. Rev. 2011, 69, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Grundy, M.M.; Edwards, C.H.; Mackie, A.R.; Gidley, M.J.; Butterworth, P.J.; Ellis, P.R. Re-evaluation of the mechanisms of dietary fiber and implications for macronutrient bio accessibility, digestion, and postprandial metabolism. Br. J. Nutr. 2016, 116, 816–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwood, A.D.; Salter, J.; Dettmar, P.W.; Chaplin, M.F. Dietary fiber, physicochemical properties, and their relationship to health. J. R. Soc. Promot. Health 2000, 120, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, C.B.; Cheng, K.-J. Molecular strategies to optimize forage and cereal digestion by ruminants. In Biotechnology and Nutrition; Bills, D.D., Kung, S.-D., Eds.; Butterworth Heinmann: Stoneham, UK, 1992; pp. 107–147. [Google Scholar]
- Akin, D.E.; Sethuraman, A.; Morrison, W.H.; Martin, S.A.; Eriksson, K.E. Microbial delignification with white rot fungi improves forage digestibility. Appl. Environ. Microbiol. 1993, 59, 4274–4282. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; McAllister, T.A. Rumen microbes, enzymes, and feed digestion-a review. Asian-Australas. J. Anim. Sci. 2002, 15, 1659–1676. [Google Scholar] [CrossRef]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell university press: Ithaca, NY, USA, 1994; pp. 1–121. [Google Scholar]
- Galyean, M.L.; Goetsch, A.L. Utilization of forage fiber by ruminants. In Forage Cell Wall Structure and Digestibility; Jung, H.G., Buxton, R.D., Hatfield, R.D., Ralph, J., Eds.; American Society of Agronomy, Inc.: Madison, WI, USA; Crop Science Society of America, Inc.: Madison, WI, USA; Soil Science Society of America, Inc.: Madison, WI, USA, 1993; pp. 33–71. [Google Scholar]
- Ko, J.K.; Lee, S.M. Advances in cellulosic conversion to fuels: Engineering yeasts for cellulosic bioethanol and biodiesel production. Curr. Opin. Biotech. 2018, 50, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Himmel, M.E.; Picataggio, S.K. Biomass recalcitrance: Deconstructing the plant cell wall for bioenergy. In Our Challenge Is to Acquire Deeper Understanding of Biomass Recalcitrance and Conversion; Himmel, M.E., Ed.; Wiley-Blackwell: Chichester, UK, 2009; pp. 1–6. [Google Scholar]
- McCann, M.C.; Carpita, N.C. Biomass recalcitrance: A multi-scale, multi-factor, and conversion-specific property. J. Exp. Bot. 2015, 66, 4109–4118. [Google Scholar] [CrossRef] [Green Version]
- La Grange, D.C.; den Haan, R.; van Zyl, W.H. Engineering cellulolytic ability into bioprocessing organisms. Appl. Microbiol. Biotechnol. 2010, 87, 1195–1208. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Reyes, L.H.; Michener, W.E.; Maness, P.C.; Chou, K.J. Engineering cellulolytic bacterium Clostridium thermocellum to co-ferment cellulose- and hemicellulose-derived sugars simultaneously. Biotechnol. Bioeng. 2018, 115, 1755–1763. [Google Scholar] [CrossRef]
- Ford, C.W.; Hartley, R.D. Cyclodimers of p-coumaric acid and ferulic acids in the cell-walls of tropical grasses. J. Sci. Food Agric. 1990, 50, 29–43. [Google Scholar] [CrossRef]
- Bunzel, M.; Ralph, J.; Brüning, P.; Steinhart, H. Structural identification of dehydrotriferulic and dehydrotetraferulic acids isolated from insoluble maize bran fiber. J. Agric. Food Chem. 2006, 54, 6409–6418. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.; Ralph, J.; Steinhart, H.; Bunzel, M. Isolation, and structural characterization of 8–O–4/8–O–4- and 8–8/8–O–4-coupled dehydrotriferulic acids from maize bran. Phytochemistry 2005, 66, 363–371. [Google Scholar] [CrossRef]
- Burr, S.J.; Fry, S.C. Extracellular cross-linking of maize arabinoxylans by oxidation of feruloyl esters to form oligoferuloyl esters and ether-like bonds. Plant J. 2009, 58, 554–567. [Google Scholar] [CrossRef] [PubMed]
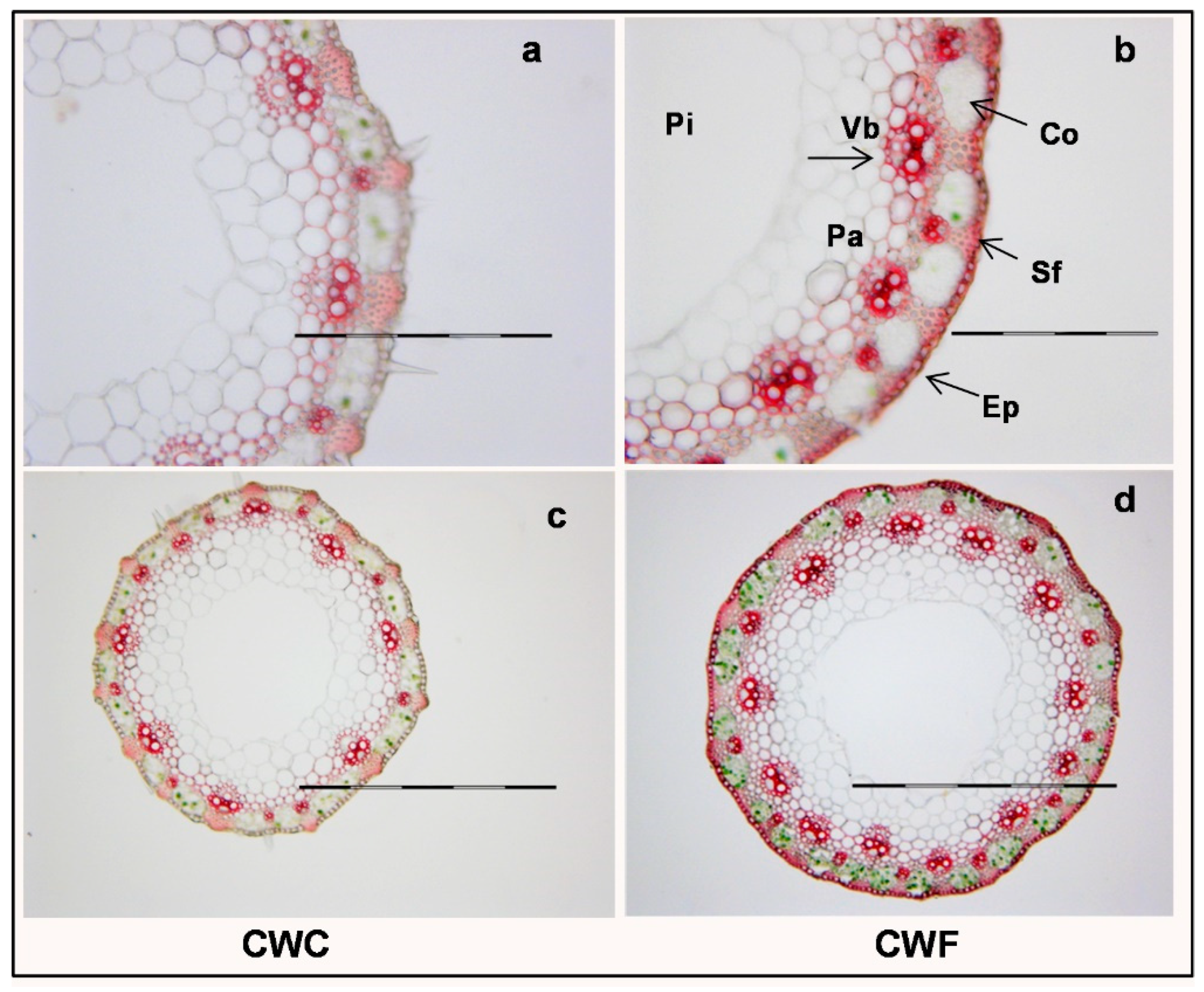
- Buanafina, M.M.d.O.; Buanafina, M.F.; Dalton, S.; Morris, P.; Kowalski, M.; Yadav, M.K.; Capper, L. Probing the role of cell wall feruloylation during maize development by differential expression of an apoplast targeted fungal ferulic acid esterase. PLoS ONE 2020, 15, e0240369. [Google Scholar] [CrossRef]
- MacAdam, J.W.; Grabber, J.H. Relationship of growth cessation with the formation of diferulate cross-links and p-coumaroylated lignins in tall fescue leaf blades. Planta 2002, 215, 785–793. [Google Scholar] [CrossRef]
- Buanafina, M.M.d.O.; Langdon, T.; Hauck, B.; Dalton, S.; Morris, P. Expression of a fungal ferulic acid esterase increases cell wall digestibility of tall fescue (festuca arundinacea). Plant Biotechnol. J. 2008, 6, 264–280. [Google Scholar] [CrossRef]
- Buanafina, M.M.d.O.; Langdon, T.; Hauck, B.; Dalton, S.J.; Morris, P. Manipulating the phenolic acid content and digestibility of italian ryegrass (Lolium multiflorum) by vacuolar-targeted expression of a fungal ferulic acid esterase. In Twenty-Seventh Symposium on Biotechnology for Fuels and Chemicals; McMillan, J.D., Adney, W.S., Mielenz, J.R., Klasson, K.T., Eds.; ABAB Symposium; Humana Press: Totowa, NJ, USA, 2006. [Google Scholar] [CrossRef]
- Waterstraat, M.; Bunzel, M. A multi-step chromatographic approach to purify radically generated ferulate oligomers reveals naturally occurring 5-5/8-8(Cyclic)-, 8-8(Noncyclic)/8-O-4-, and 5-5/8-8(Noncyclic)-coupled dehydrotriferulic acids. Front. Chem. 2018, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Grabber, J.H.; Hatfield, R.D.; Ralph, J.; Zoń, J.; Amrhein, N. Ferulate cross-linking in cell walls isolated from maize cell suspensions. Phytochemistry 1995, 40, 1077–1082. [Google Scholar] [CrossRef]
- Fry, S.C.; Willis, S.C.; Paterson, A.E. Intraprotoplasmic and wall-localised formation of arabinoxylan-bound diferulates and larger ferulate coupling-products in maize cell-suspension cultures. Planta 2000, 211, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, S.E.; Fry, S.C. Control of diferulate formation in dicotyledonous and gramineous cell-suspension cultures. Planta 2008, 227, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Encina, A.; Fry, S.C. Oxidative coupling of a feruloyl-arabinoxylan trisaccharide (FAXX) in the walls of living maize cells requires endogenous hydrogen peroxide and is controlled by a low-M r apoplastic inhibitor. Planta 2005, 223, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Obel, N.; Porchia, A.; Scheller, H. Intracellular feruloylation of arabinoxylan in wheat: Evidence for feruloyl-glucose as precursor. Planta 2003, 216, 620–629. [Google Scholar] [CrossRef]
- Ralph, J.; Grabber, J.H.; Hatfield, R.D. Lignin-ferulate cross-links in grasses: Active incorporation of ferulate polysaccharide esters into ryegrass lignins. Carbohyd. Res. 1995, 275, 167–178. [Google Scholar] [CrossRef]
- Grabber, J.H.; Ralph, J.; Hatfield, R.D. Cross-linking of maize walls by ferulate dimerization and incorporation into lignin. J. Agric. Food Chem. 2000, 48, 6106–6113. [Google Scholar] [CrossRef] [PubMed]
- Grabber, J.H.; Ralph, J.; Hatfield, R.D. Model studies of ferulate−coniferyl alcohol cross-product formation in primary maize walls: implications for lignification in grasses. J. Agric. Food Chem. 2002, 50, 6008–6016. [Google Scholar] [CrossRef] [PubMed]
- Barros-Rios, J.; Malvar, R.A.; Jung, H.-J.G.; Bunzel, M.; Santiago, R. Divergent selection for ester-linked diferulates in maize pith stalk tissues. Effects on cell wall composition and degradability. Phytochemistry 2012, 83, 43–50. [Google Scholar] [CrossRef]
- Casler, M.D.; Jung, H.G. Selection and evaluation of smooth bromegrass clones with divergent lignin or etherified ferulic acid concentration. Crop Sci. 1999, 39, 1866–1873. [Google Scholar] [CrossRef]
- Barrière, Y.; Thomas, J.; Denoue, D. QTL mapping for lignin content, lignin monomeric composition, p-hydroxycinnamate content, and cell wall digestibility in the maize recombinant inbred line progeny F838 × F286. Plant Sci. 2008, 175, 585–595. [Google Scholar] [CrossRef]
- Barrière, Y.; Méchin, V.; Lefevre, B.; Maltese, S. QTLs for agronomic and cell wall traits in a maize RIL progeny derived from a cross between an old Minnesota13 line and a modern Iodent line. Theor. Appl. Genet. 2012, 125, 531–549. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.A.C.; Dupree, P.; Shewry, P.R. A novel bioinformatics approach identifies candidate genes for the synthesis and feruloylation of arabinoxylan. Plant Physiol. 2007, 144, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riboulet, C.; Fabre, F.; Dénoue, D.; Martinantä, J.P.; Lefèvre, B.; Barrière, Y. QTL mapping and candidate gene research for lignin content and cell wall digestibility in a top-cross of a flint maize recombinant inbred line progeny harvested at silage stage. Maydica 2008, 53, 1–9. [Google Scholar]
- Courtial, A.; Méchin, V.; Reymond, M.; Grima-Pettenati, J.; Barrière, Y. Colocalizations between several QTLs for Cell Wall degradability and composition in the F288 × F271 early maize RIL progeny raise the question of the nature of the possible underlying determinants and breeding targets for biofuel capacity. Bioenergy Res. 2014, 7, 142–156. [Google Scholar] [CrossRef]
- Cogan, N.O.I.; Smith, K.F.; Yamada, T.; Francki, M.G.; Vecchies, A.C.; Jones, E.S.; Spangenberg, G.C.; Forster, J.W. QTL analysis and comparative genomics of herbage quality traits in perennial ryegrass (lolium perenne L.). Theor. Appl. Genet. 2005, 110, 364–380. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, K.; Hu, X.; Wu, Y.; Huang, C. Genome-wide association analysis of forage quality in maize mature stalk. BMC Plant Biol. 2016, 16, 227. [Google Scholar] [CrossRef] [Green Version]
- López-Malvar, A.; Butrón, A.; Samayoa, L.F.; Figueroa-Garrido, D.J.; Malvar, R.A.; Santiago, R. Genome-wide association analysis for maize stem cell wall-bound hydroxycinnamates. BMC Plant Biol. 2019, 19, 519. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.G.; Phillips, R.L. Putative seedling ferulate ester (sfe) maize mutant: Morphology, biomass yield, and stover cell wall composition and rumen degradability. Crop Sci. 2010, 50, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Jung, G.; Mertens, D.R.; Phillips, R.L. Effect of reduced ferulate-mediated lignin/arabinoxylan cross-linking in corn silage on feed intake, digestibility, and milk production. J. Dairy Sci. 2011, 94, 5124–5137. [Google Scholar] [CrossRef] [PubMed]
- Jorgenson, L.R. Brown midrib in maize and its linkage relations. J. Am. Soc. Agron. 1931, 23, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Cherney, J.H.; Cherney, D.J.R.; Akin, D.E.; Axtell, J.D. Potential of brown-midrib, low-lignin mutants for improving forage quality. Adv. Agron. 1991, 46, 157–198. [Google Scholar]
- Barrière, Y.; Argillier, O. Brown-midrib genes of maize: A review. Agronomie 1993, 13, 865–876. [Google Scholar] [CrossRef] [Green Version]
- Lam, T.B.T.; Iiyama, K.; Stone, B.A. Lignin and hydroxycinnamic acids in walls of brown midrib mutants of sorghum, pearl millet and maize stems. J. Sci. Food Agric. 1996, 71, 174–178. [Google Scholar] [CrossRef]
- Chabbert, B.; Tollier, M.T.; Monties, B.; Barrière, Y.; Argillier, O. Biological variability in lignification of maize: Expression of the brown midrib bm2 mutation. J. Sci. Food Agric. 1994, 64, 455–460. [Google Scholar] [CrossRef]
- Vermerris, W.; Boon, J.J. Tissue-specifi c patterns of lignifi cation are disturbed in the brown midrib2 mutant of maize (Zea mays L.). J. Agric. Food Chem. 2001, 49, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Marita, J.; Vermerris, W.; Ralph, J.; Hatfield, R.D. Variations in the cell wall composition of maize brown midrib mutants. J. Agric. Food Chem. 2003, 51, 1313–1321. [Google Scholar] [CrossRef]
- Barrière, Y.; Ralph, J.; Méchin, V.; Guillaumie, S.; Grabber, J.H.; Argillier, O.; Chabbert, B.; Lapierre, C. Genetic and molecular basis of grass cell wall biosynthesis and degradability. II. Lessons from brown midrib mutants. C. R. Biol. 2004, 327, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Vignols, F.; Rigau, J.; Torres, M.A.; Capellades, M.; Puigdomènech, P. The brown-midrib3 (bm3) mutation in maize occurs in the gene encoding caffeic acid O-methyl transferase. Plant Cell 1995, 7, 407–416. [Google Scholar] [PubMed] [Green Version]
- Yoshida-Shimokawa, T.; Yoshida, S.; Kakegawa, K.; Ishii, T. Enzymic feruloylation of arabinoxylan-trisaccharide by feruloyl-CoA: Arabinoxylan-trisaccharide O-hydroxycinnamoyl transferase from Oryza sativa. Planta 2001, 212, 470–474. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Bontpart, T.; Cheynier, V.; Ageorges, A.; Terrier, N. BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds. New Phytol. 2015, 208, 695–707. [Google Scholar] [CrossRef]
- Rautengarten, C.; Birdseye, D.; Pattathil, S.; McFarlane, H.E.; Saez-Aguayo, S.; Orellana, A.; Persson, S.; Hahn, M.G.; Scheller, H.V.; Heazlewood, J.L.; et al. The elaborate route for UDP-arabinose delivery into the Golgi of plants. Proc. Natl. Acad. Sci. USA 2017, 14, 4261–4266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buanafina, M.M.d.O. Feruloylation in grasses: Current and future perspectives. Mol. Plant 2009, 2, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Rennie, E.A.; Scheller, H.V. Xylan biosynthesis. Curr. Opin. Biotechnol. 2014, 26, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Chateigner-Boutin, A.L.; Ordaz-Ortiz, J.J.; Alvarado, C.; Bouchet, B.; Durand, S.; Verhertbruggen, Y.; Barrière, Y.; Saulnier, L. Developing pericarp of maize: A model to study arabinoxylan synthesis and feruloylation. Front. Plant Sci. 2016, 7, 1476. [Google Scholar] [CrossRef] [Green Version]
- Piston, F.; Uauy, C.; Fu, L.H.; Langston, J.; Labavitch, J.; Dubcovsky, J. Down-regulation of four putative arabinoxylan feruloyl transferase genes from family PF02458 reduces ester-linked ferulate content in rice cell walls. Planta 2010, 231, 677–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinari, H.B.C.; Pellny, T.K.; Freeman, J.; Shewry, P.R.; Mitchell, R.A.C. Grass cell wall feruloylation: Distribution of bound ferulate and candidate gene expression in Brachypodium distachyon. Front. Plant Sci. 2013, 4, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartley, L.E.; Peck, M.L.; Kim, S.R.; Ebert, B.; Maniseri, C.; Chiniquy, D.M.; Sykes, R.; Gao, L.; Rautengarten, C.; Vega-Sánchez, M.E.; et al. Overexpression of a BAHD acyltransferase, OsAt10, alters rice cell wall hydroxycinnamic acid content and saccharification. Plant Physiol. 2013, 161, 1615–1633. [Google Scholar] [CrossRef] [Green Version]
- Buanafina, M.M.d.O.; Fescemyer, H.W.; Sharma, M.; Shearer, E.S. Functional testing of a PF02458 homologue of putative rice arabinoxylan feruloyl transferase genes in Brachypodium distachyon. Planta 2016, 243, 659–674. [Google Scholar] [CrossRef]
- Li, G.; Jones, K.C.; Eudes, A.; Pidatala, V.R.; Sun, J.; Xu, F.; Ronald, P.C. Overexpression of a rice BAHD acyltransferase gene in switchgrass (Panicum virgatum L.) enhances saccharification. BMC Biotechnol. 2018, 18, 54. [Google Scholar] [CrossRef]
- Tian, Y.; Lin, C.Y.; Park, J.H.; Wu, C.Y.; Kakumanu, R.; Pidatala, V.R.; Eudes, A. Overexpression of the rice BAHD acyltransferase AT10 increases xylan-bound p-coumarate and reduces lignin in Sorghum bicolor. Biotechnol. Biofuels 2021, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, A.; Rancour, D.M.; Sullivan, M.; Karlen, S.D.; Ralph, J.; Riaño-Pachón, D.M.; Vicentini, R.; Silva, T.D.F.; Ferraz, A.; Hatfield, R.D.; et al. Overexpression of a sugarcane BAHD acyltransferase alters hydroxycinnamate content in maize cell wall. Front. Plant Sci. 2021, 12, 626168. [Google Scholar] [CrossRef] [PubMed]
- de Souza, W.R.; Martins, P.K.; Freeman, J.; Pellny, T.K.; Michaelson, L.V.; Sampaio, B.L.; Vinecky, F.; Ribeiro, A.P.; da Cunha, B.A.D.B.; Kobayashi, A.K.; et al. Suppression of a single BAHD gene in Setaria viridis causes large, stable decreases in cell wall feruloylation and increases biomass digestibility. New Phytolog. 2017, 18, 81–93. [Google Scholar]
- de Souza, W.R.; Pacheco, T.F.; Duarte, K.E.; Sampaio, B.L.; Molinari, P.A.D.O.; Martins, P.K.; Santiago, T.R.; Formighieri, E.F.; Vinecky, F.; Ribeiro, A.P.; et al. Silencing of a BAHD acyltransferase in sugarcane increases biomass digestibility. Biotechnol. Biofuels 2019, 12, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mota, T.R.; de Souza, W.R.; Oliveira, D.M.; Martins, P.K.; Bruno, L. Suppression of a BAHD acyltransferase decreases p-coumaroyl on arabinoxylan and improves biomass digestibility in the model grass Setaria viridis. Plant J. 2021, 105, 136–150. [Google Scholar] [CrossRef]
- Freeman, J.; Ward, J.L.; Kosik, O.; Lovegrove, A.; Wilkinson, M.D.; Shewry, P.R.; Mitchell, R.A.C. Feruloylation and structure of arabinoxylan in wheat endosperm cell walls from RNAi lines with suppression of genes responsible for backbone synthesis and decoration. Plant Biotechnol. J. 2017, 15, 1429–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, N.; Wilkinson, M.D.; Lovegrove, A.; Freeman, J.; Tryfona, T.; Pellny, T.K.; Weimar, T.; Mortimer, J.C.; Stott, K.; Baker, J.M.; et al. Glycosyl transferases in family 61 mediate arabinofuranosyl transfer onto xylan in grasses. Proc. Natl. Acad. Sci. USA 2012, 109, 989–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, X.; Dixon, R.A. Current models for transcriptional regulation of secondary cell wall biosynthesis in grasses. Front. Plant Sci. 2018, 9, 399. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, R.; Dalton, S.; Roberts, L.A.; Moron-Garcia, O.M.; Lacono, R.; Kosik, O.; Gallagher, J.A.; Bosch, M. Modified expression of ZmMYB167 in Brachypodium distachyon and Zea mays leads to increased cell wall lignin and phenolic content. Nat. Sci. Rep. 2019, 9, 8800. [Google Scholar] [CrossRef] [PubMed]
- Buanafina, M.M.d.O.; Langdon, T.; Hauck, B.; Dalton, S.; Timms-Taravella, E.; Morris, P. Targeting expression of a fungal ferulic acid esterase to the apoplast, endoplasmic reticulum or Golgi can disrupt feruloylation of the growing cell wall and increase the biodegradability of tall fescue (Festuca arundinacea). Plant Biotechn. J. 2010, 8, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Harholt, J.; Bach, I.C.; Lind-Bouquin, S.; Nunan, K.J.; Madrid, S.M.; Brinch-Pedersen, H.; Holm, P.B.; Scheller, H.V. Generation of transgenic wheat (Triticum aestivum L.) accumulating heterologous endo-xylanase or ferulic acid esterase in the endosperm. Plant Biotech. J. 2010, 8, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
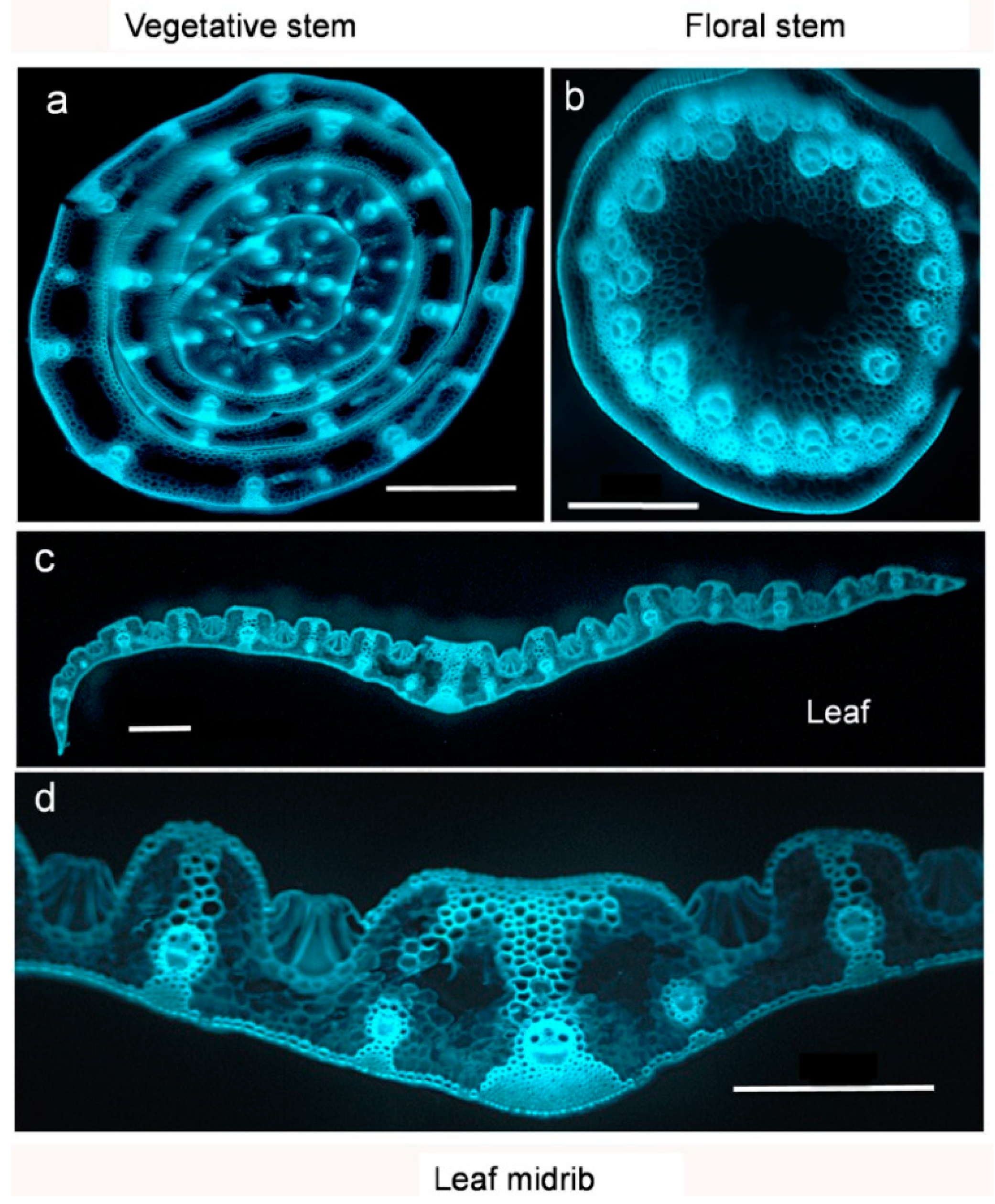
- Buanafina, M.M.d.O.; Iyer, P.R.; Buanafina, M.F.; Shearer, E.A. Reducing cell wall feruloylation by expression of a fungal ferulic acid esterase in Festuca arundinacea modifies plant growth, leaf morphology and the turnover of cell wall arabinoxylans. PLoS ONE 2017, 12, e0185312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogorelko, G.; Fursova, O.; Lin, M.; Pyle, E.; Jass, J.; Zabotina, O.A. Post-synthetic modification of plant cell walls by expression of microbial hydrolases in the apoplast. Plant Mol. Biol. 2011, 77, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Badhan, A.; Jin, L.; Wang, Y.; Han, S.; Kowalczys, K.; Brown, D.C.W.; Ayala, C.J.; Latoszek-Green, M.; Miki, B.; Tsang, A.; et al. Expression of a fungal ferulic acid esterase in alfalfa modifies cell wall digestibility. Biotechnol. Biofuels 2014, 7, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buanafina, M.M.d.O.; Langdon, T.; Dalton, S.; Morris, P. Expression of a Trichoderma reesei ß-1,4 endoxylanase in tall fescue modifies cell wall structure and digestibility and elicits pathogen defence responses. Planta 2012, 236, 1757–1774. [Google Scholar] [CrossRef] [PubMed]
- Buanafina, M.M.d.O.; Dalton, S.; Langdon, T.; Timms-Taravella, E.; Shearer, E.A.; Morris, P. Functional co-expression of a fungal ferulic acid esterase and a ß-1,4 endo-xylanase in Festuca arundinacea (tall fescue) modifies post-harvest cell wall deconstruction. Planta 2015, 242, 97–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier d’Yvoire, M.; Bouchabke-Coussa, O.; Voorend, W.; Antelme, S.; Cézard, L.; Legée, F.; Lebris, P.; Legay, S.; Whitehead, C.; McQueen-Mason, S.J.; et al. Disrupting the cinnamyl alcohol dehydrogenase 1 gene (BdCAD1) leads to altered lignification and improved saccharification in Brachypodium distachyon. Plant J. 2013, 73, 496–508. [Google Scholar] [CrossRef]
- Eudes, A.; Sathitsuksanoh, N.; Baidoo, E.E.; George, A.; Liang, Y.; Yang, F.; Singh, S.; Keasling, J.D.; Simmons, B.A.; Loqué, D. Expression of a bacterial 3-dehydroshikimate dehydratase reduces lignin content and improves biomass saccharification efficiency. Plant Biotechnol. J. 2015, 13, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.L.; Waldron, K.W. Texture of Chinese water chestnut: Involvement of cell wall phenolics. J. Sci. Food Agric. 1995, 68, 337–346. [Google Scholar] [CrossRef]
- Ng, A.; Harvey, A.J.; Parker, M.L.; Smith, A.C.; Waldron, K.W. Effect of oxidative coupling on the thermal stability of texture and cell wall chemistry of beet root (Beta vulgaris). J. Agric. Food Chem. 1998, 46, 3365–3370. [Google Scholar] [CrossRef]
- Fry, S.C. Cross-linking of matrix polymers in the growing cell walls of angiosperms. Ann. Rev. Plant Phys. 1986, 37, 165–186. [Google Scholar] [CrossRef]
- Liyama, K.; Lam, T.B.; Stone, B.A. Phenolic acid bridges between polysaccharides and lignin in wheat internodes. Phytochemistry 1990, 29, 733–737. [Google Scholar]
- Kamisaka, S.; Takeda, S.; Takahashi, K.; Shibata, K. Diferulic and ferulic acid in the cell wall of Avena coleoptiles—Their relationships to mechanical properties of the cell wall. Physiol. Plant. 1990, 78, 1–77. [Google Scholar] [CrossRef]
- Tan, K.-S.; Hoson, T.; Masuda, Y.; Kamisaka, S. Correlation between cell wall extensibility and the content of diferulic and ferulic acids in cell walls of Oryza sativa coleoptiles grown under water and in air. Physiol. Plant 1991, 83, 397–403. [Google Scholar] [CrossRef]
- Azuma, T.; Okita, N.; Nanmori, T.; Yasuda, T. Changes in cell wall-bound phenolic acids in the internodes of submerged floating rice. Plant Prod. Sci. 2005, 8, 441–446. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Soga, K.; Hoson, T. Phenylalanine ammonia-lyase and cell wall peroxidase are cooperatively involved in the extensive formation of ferulate network in cell walls of developing rice shoots. J. Plant Physiol. 2012, 168, 262–267. [Google Scholar] [CrossRef]
- Whetten, R.W.; MacKay, J.J.; Sederoff, R.R. Recent advances in understanding lignin biosynthesis. Ann. Rev. Plant Biol. 1998, 49, 585–609. [Google Scholar] [CrossRef]
- Fry, S.C. Gibberellin-controlled pectinic acid and protein secretion in growing cells. Phytochemistry 1980, 19, 735–740. [Google Scholar] [CrossRef]
- Clissold, F.J. The biomechanics of chewing and plant fracture: Mechanisms and implications. In Advances in Insect Physiology; Casas, J., Simpson, S.J., Eds.; Elsevier Science: London, UK, 2008; pp. 317–372. [Google Scholar]
- Read, J.; Stokes, A. Plant biomechanics in an ecological context. Am. J. Bot. 2006, 93, 1546–1565. [Google Scholar] [CrossRef]
- Faulds, C.; Williamson, G. The role of hydroxycinnamates in the plant cell wall. J. Sci. Food Agric. 1999, 79, 393–395. [Google Scholar] [CrossRef]
- García-Lara, S.; Bergvinson, D.J.; Burt, A.J.; Ramputh, A.I.I.; Diaz-Pontones, D.M.; Arnason, J.T. The role of pericarp cell wall components in maize weevil resistance. Crop Sci. 2004, 44, 1546–1552. [Google Scholar] [CrossRef]
- Santiago, R.; Butrón, A.; Reid, L.M.; Arnason, J.T.; Sandoya, G.; Souto, X.C.; Malvar, R.A. Diferulate content of maize sheaths is associated with resistance to the Mediterranean corn borer Sesamia nonagrioides (Lepidoptera: Noctuidae). J. Agric. Food Chem. 2006, 54, 9140–9144. [Google Scholar] [CrossRef] [PubMed]
- Lyons, P.C.; Hipskind, J.; Vincent, J.R.; Nicholson, R.L. Phenylpropanoid dissemination in maize resistant or susceptible to Helminthosporium maydis. Maydica 1993, 38, 175–181. [Google Scholar]
- Ikegawa, T.; Mayama, S.; Nakayashiki, H.; Kato, H. Accumulation of diferulic acid during the hypersensitive response of oat leaves to Puccinia coronata f.sp. avenae and its role in the resistance of oat tissues to cell wall degrading enzymes. Physiol. Mol. Plant Pathol. 1996, 48, 245–256. [Google Scholar]
- Bily, A.C.; Reid, L.M.; Taylor, J.H.; Johnston, D.; Malouin, C.; Burt, A.J.; Bakan, B.; Regnault-Roger, C.; Pauls, K.P.; Arnason, J.T.; et al. Dehydrodimers of ferulic acid in maize grain pericarp and aleurone: Resistance factors to Fusarium graminearum. Phytopathology 2003, 93, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, N.J.; Casler, M.D.; Grau, C.R.; Jung, H.G. Reactions of smooth bromegrass clones with divergent lignin or etherified ferulic acid concentration to three fungal pathogens. Crop Sci. 2002, 42, 1824–1831. [Google Scholar] [CrossRef] [Green Version]
- Barros-Rios, J.; Santiago, R.; Jung, H.J.G.; Malvar, R.A. Covalent cross-linking of cell wall polysaccharides through esterified diferulates as a maize resistance mechanism against corn borers. J. Agric. Food Chem. 2015, 63, 2206–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, R.; Sandoya, G.; Butrón, A.; Barros, J.; Malvar, R.A. Changes in phenolic concentrations during recurrent selection for resistance to the Mediterranean corn borer (Sesamia nonagrioides Lef.). J. Agric. Food Chem. 2008, 56, 8017–8022. [Google Scholar] [CrossRef] [PubMed]
- Buanafina, M.M.d.O.; Fescemyer, H. Modification of esterified cell wall phenolics increases vulnerability of tall fescue to herbivory by the fall armyworm. Planta 2012, 236, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Reem, N.T.; Pogorelko, G.; Lionetti, V.; Chambers, L.; Held, M.A.; Bellincampi, D.; Zabotina, O.A. Decreased polysaccharide feruloylation compromises plant cell wall integrity and increases susceptibility to necrotrophic fungal pathogens. Front. Plant Sci. 2016, 7, 630. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, S.; Reem, N.T.; Lionetti, V.; Zabotina, O.A. Coexpression of fungal cell wall-modifying enzymes reveals their additive impact on Arabidopsis resistance to the fungal pathogen, Botrytis cinerea. Biology 2021, 10, 1070. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, U. Cessation of cell elongation in rye coleoptiles is accompanied by a loss of cell-wall plasticity. J. Exper. Bot. 1996, 7, 1387–1394. [Google Scholar] [CrossRef] [Green Version]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2009, 9, 189–195. [Google Scholar] [CrossRef] [PubMed]
- De Costa, W.; Zörb, C.; Hartung, W.; Schubert, S. Salt resistance is determined by osmotic adjustment and abscisic acid in newly developed maize hybrids in the first phase of salt stress. Physiol. Plant. 2007, 131, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Z.; Neumann, P.M. Water-stressed maize, barley, and rice seedlings show species diversity in mechanisms of leaf growth inhibition. J. Exper. Bot. 1998, 49, 1945–1952. [Google Scholar] [CrossRef]
- Reuber, S.; Bornman, J.F.; Weissenböck, G. Phenylpropanoid compounds in primary leaf tissues of rye (Secale cereale). Light response of their metabolism and the possible role in UV-B protection. Physiolog. Plant. 1996, 97, 160–168. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Grzesiak, S. Possible contribution of cell-wall-bound ferulic acid in drought resistance and recovery in triticale seedlings. J. Plant Physiol. 2009, 166, 1720–1733. [Google Scholar] [CrossRef]
- Hura, T.; Hura, K.; Dziurka, K.; Ostrowska, A.; Bączek-Kwinta, R.; Grzesiak, M. An increase in the content of cell wall-bound phenolics correlates with the productivity of triticale under soil drought. J. Plant Physiol. 2012, 169, 1728–1736. [Google Scholar] [CrossRef]
- Uddin, N.; Stefan Hanstein, S.; Faust, F.; Eitenmüller, P.T.; Pitann, B.; Schubert, S. Diferulic acids in the cell wall may contribute to the suppression of shoot growth in the first phase of salt stress in maize. Phytochemistry 2014, 102, 126–136. [Google Scholar] [CrossRef]
- Fry, S.C. Phenolic components of the primary cell wall and their possible role in the hormonal regulation of growth. Planta 1979, 146, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, L. Transport barriers made of cutin, suberin and associated waxes. Trends Plant Sci. 2010, 15, 546–553. [Google Scholar] [CrossRef]
- Chen, F.; Dixon, R. Lignin modification improves fermentable sugar yields for biofuel production. Nat. Biotechnol. 2007, 25, 759–761. [Google Scholar] [CrossRef] [PubMed]
- Saleme, M.L.S.; Cesarino, I.; Vargas, L.; Kim, H.; Vanholme, R.; Goeminne, G.; Van Acker, R.; Fonseca, F.C.A.; Pallidis, A.; Voorend, W.; et al. Silencing Caffeoyl shikimate esterase affects lignification and improves saccharification in Poplar. Plant Physiol. 2017, 175, 1040–1057. [Google Scholar] [CrossRef] [Green Version]
- Pederson, J.F.; Vogel, K.P.; Funnel, D.L. Impact of reduced lignin on plant fitness. Crop Sci. 2005, 45, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.-J.G.; Samac, D.A.; Sarath, G. Modifying crops to increase cell wall digestibility. Plant Sci. 2012, 185, 65–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauly, M.; Keegstra, K. Plant cell wall polymers as precursors for biofuels. Curr. Opin. Plant Biol. 2010, 13, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Hartley, R.D. p-coumaric and ferulic acid components of cell walls of ryegrass and their relationships with lignin and digestibility. J. Sci. Food Agric. 1972, 23, 1347–1354. [Google Scholar] [CrossRef]
- Grabber, J.H.; Hatfield, R.D.; Ralph, J. Diferulate cross-links impede the enzymatic degradation of non-lignified maize walls. J. Sci. Food Agric. 1998, 77, 193–200. [Google Scholar] [CrossRef]
- Grabber, J.H.; Mertens, D.R.; Kim, H.; Funk, C.; Lu, F.; Ralph, J. Cell wall fermentation kinetics are impacted more by lignin content and ferulate cross-linking than by lignin composition. J. Sci. Food Agric. 2009, 89, 122–129. [Google Scholar] [CrossRef]
- Liyama, K.; Lam, T.B.T. Structural characteristics of cell walls of forage grasses—their nutritional evaluation for ruminants. Asian Australasian. J. Anim. Sci. 2001, 14, 862–879. [Google Scholar] [CrossRef]
- Lam, T.B.T.; Iiyama, K.; Stone, B.A. Hot alkali-labile linkages in the walls of the forage grass Phalaris aquatica and Lolium perenne and their relation to in vitro wall digestibility. Phytochemistry 2003, 64, 603–607. [Google Scholar] [CrossRef]
- Casler, M.D.; Jung, H.G.; Coblentz, W.K. Clonal selection for lignin and etherified ferulates in three perennial grasses. Crop Sci. 2008, 48, 424–433. [Google Scholar] [CrossRef]
- Morris, P.; Dalton, S.; Langdon, T.; Hauck, B.; Buanafina, M.M.d.O. Expression of a fungal ferulic acid esterase in suspension cultures of tall fescue (Festuca arundinacea) decreases cell wall feruloylation and increases rates of cell wall digestion. Plant Cell Tissue Organ Cult. (PCTOC) 2017, 129, 181–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buanafina, M.M.d.O.; Morris, P. The Impact of Cell Wall Feruloylation on Plant Growth, Responses to Environmental Stress, Plant Pathogens and Cell Wall Degradability. Agronomy 2022, 12, 1847. https://doi.org/10.3390/agronomy12081847
Buanafina MMdO, Morris P. The Impact of Cell Wall Feruloylation on Plant Growth, Responses to Environmental Stress, Plant Pathogens and Cell Wall Degradability. Agronomy. 2022; 12(8):1847. https://doi.org/10.3390/agronomy12081847
Chicago/Turabian StyleBuanafina, Marcia Maria de O., and Phillip Morris. 2022. "The Impact of Cell Wall Feruloylation on Plant Growth, Responses to Environmental Stress, Plant Pathogens and Cell Wall Degradability" Agronomy 12, no. 8: 1847. https://doi.org/10.3390/agronomy12081847