
Water Stress Impacts on Grapevines (Vitis vinifera L.) in Hot Environments: Physiological and Spectral Responses

 ,
,  , ,
, ,  , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
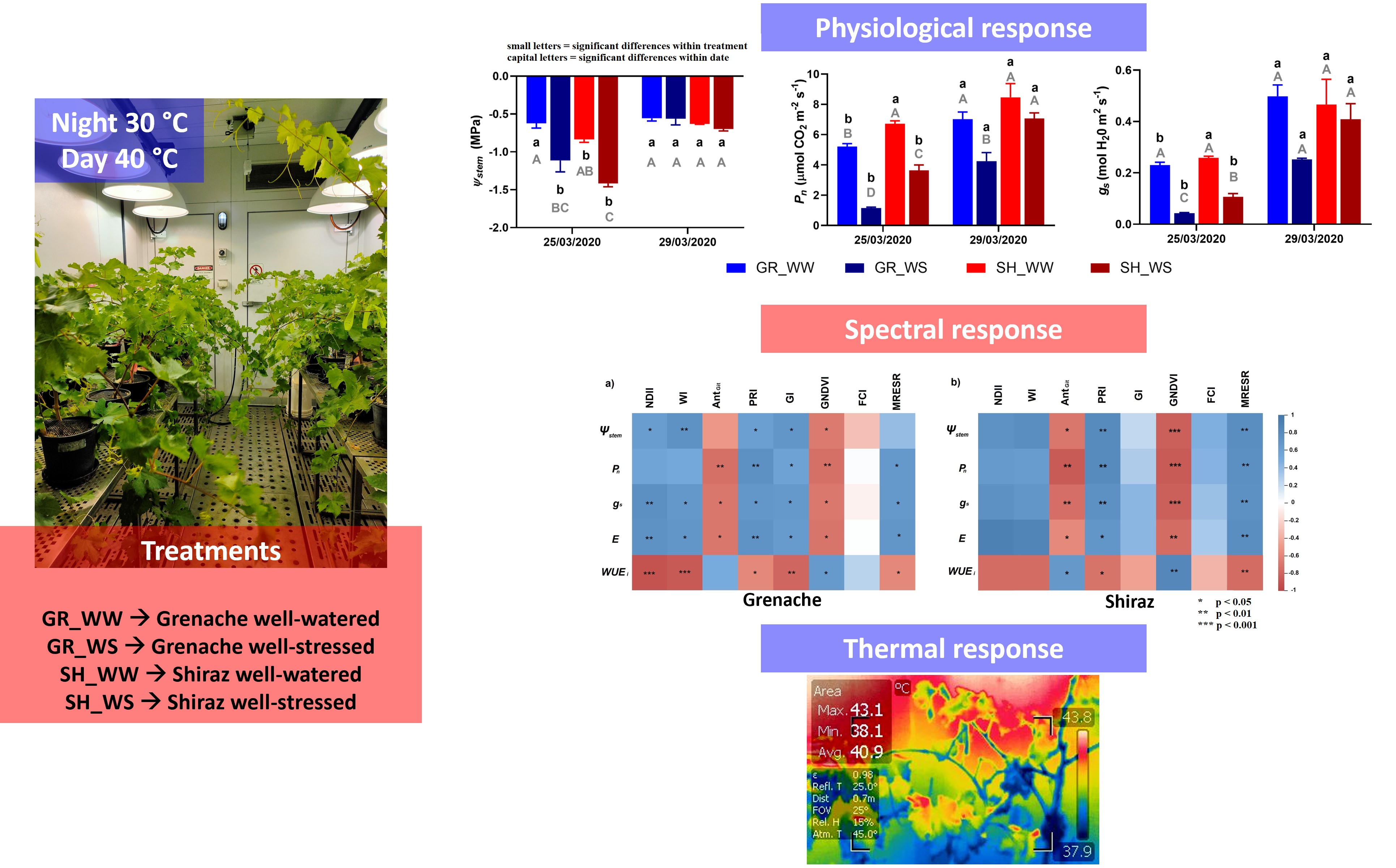
2.1. Treatments
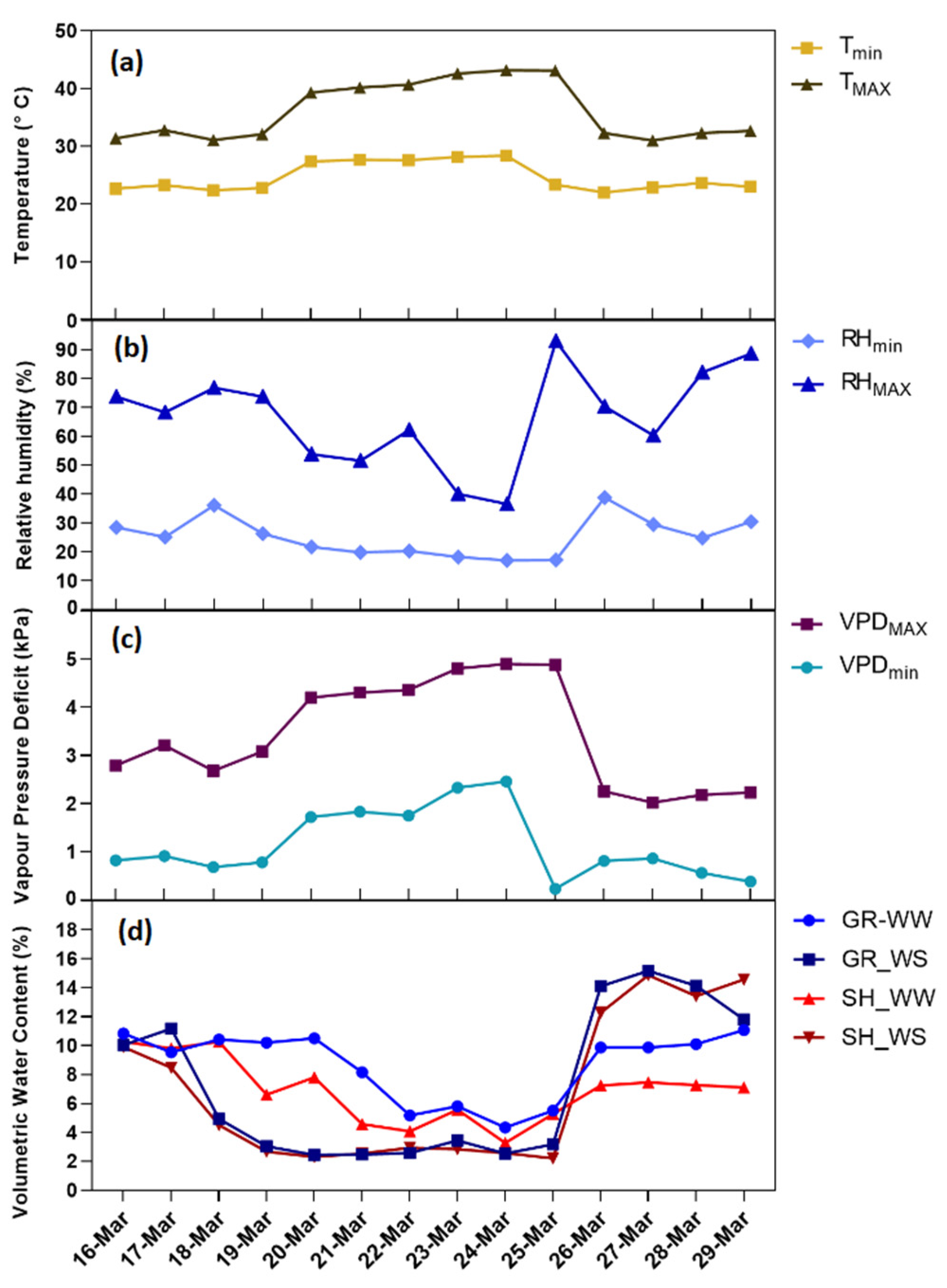
2.2. Environmental Conditions
2.3. Soil Moisture
2.4. Physiological Measurements
2.5. Hyperspectral Measurements
2.6. Thermal Measurements
2.7. Statistical Analyses
3. Results
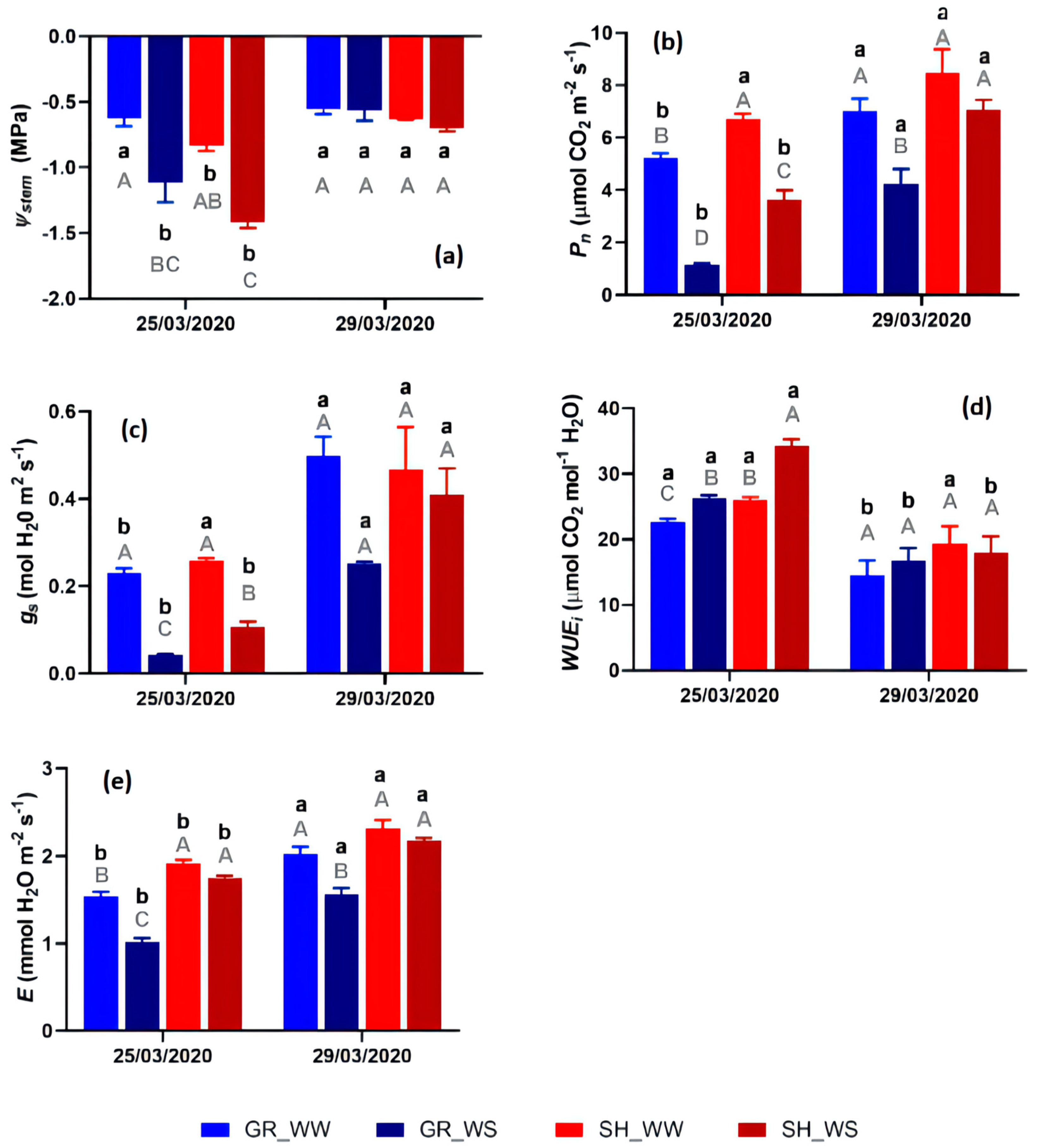
3.1. Grapevine Physiological Responses
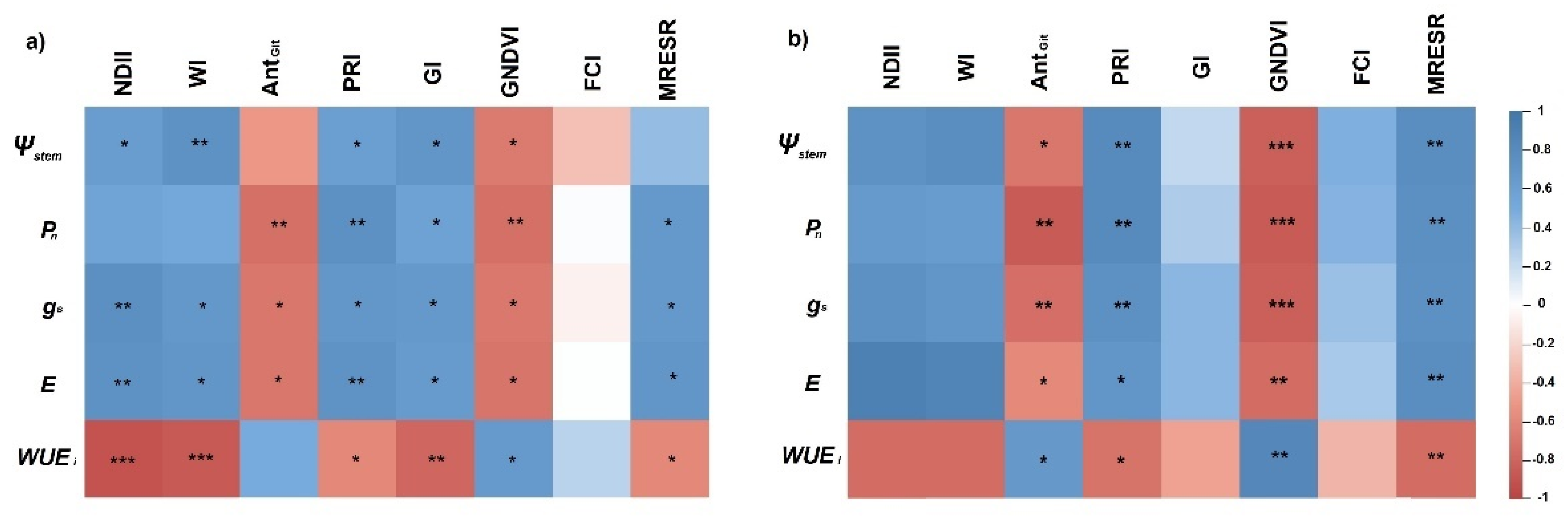
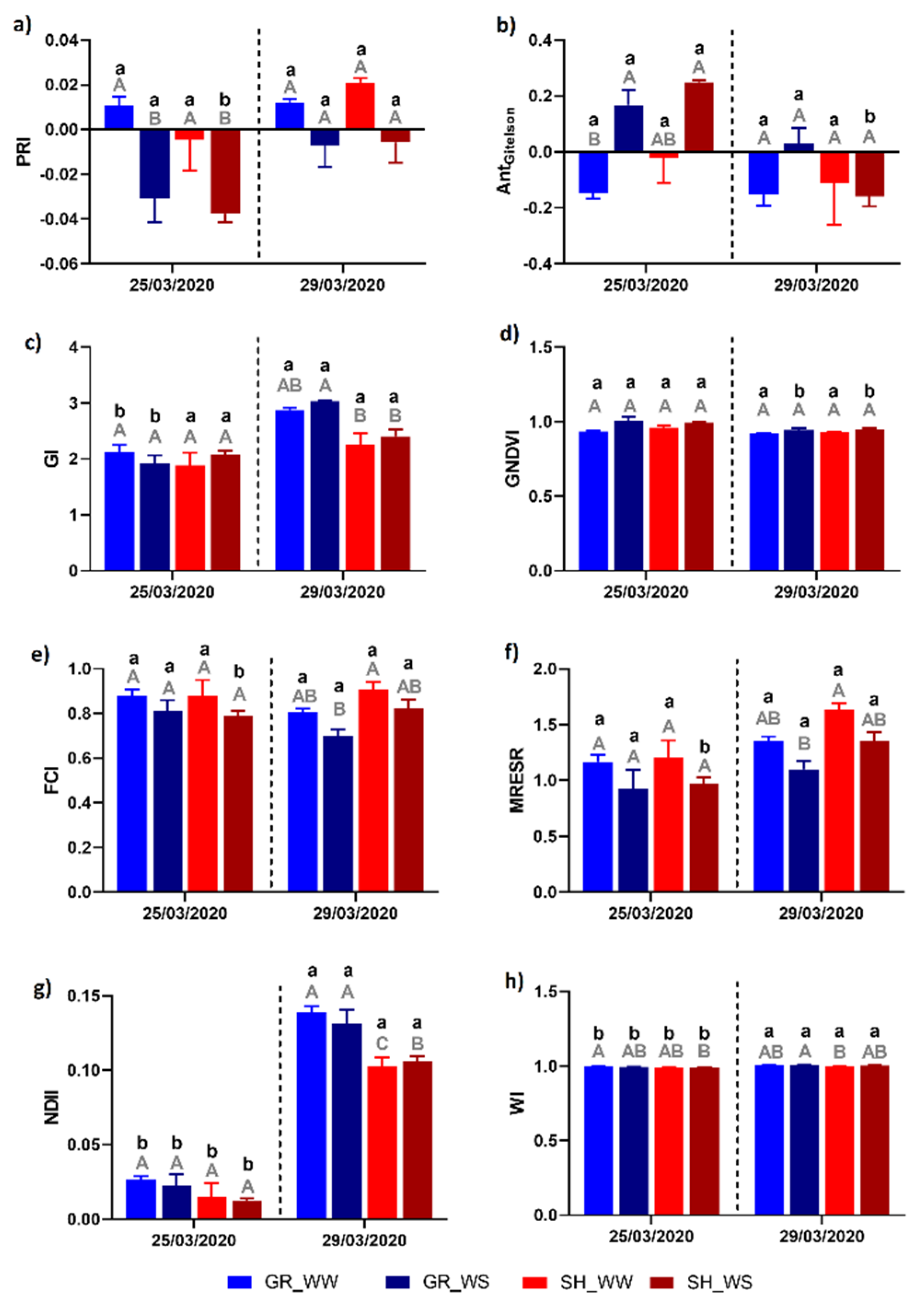
3.2. Hyperspectral Response
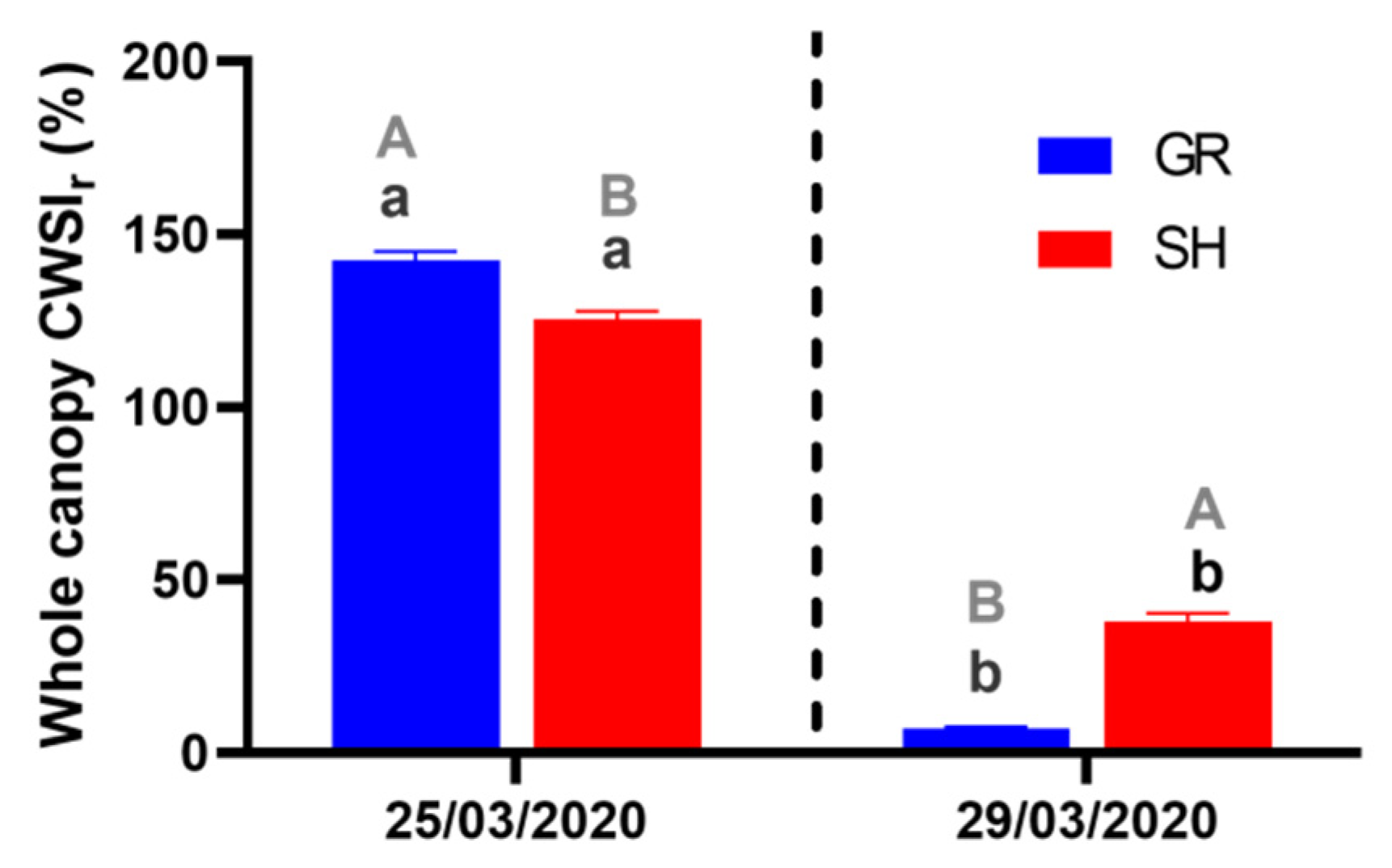
3.3. Canopy Temperature
3.4. Relationship between VIs and Physiological Parameters of Grenache and Shiraz
3.4.1. Physiological Parameters Estimation with VIs in Grenache
3.4.2. Physiological Parameters Estimation with VIs in Shiraz
4. Discussion
4.1. Physiological Monitoring of Grenache and Shiraz under High Temperature
4.2. Potential of the Thermal and Spectral Vegetation Indices to Detect Drought Stress in Grapevine
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change—The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Paris, France, 2021. [Google Scholar] [CrossRef]
- Bureau of Meteorology. Available online: http://www.bom.gov.au (accessed on 9 December 2018).
- Perkins-Kirkpatrick, S.E.; Lewis, S.C. Increasing trends in regional heatwaves. Nat. Commun. 2020, 11, 3357. [Google Scholar] [CrossRef] [PubMed]
- Trancoso, R.; Syktus, J.; Toombs, N.; Ahrens, D.; Wong, K.K.H.; Pozza, R.D. Heatwaves intensification in Australia: A consistent trajectory across past, present and future. Sci. Total Environ. 2020, 742, 140521. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.C.; Carvalho, D.; Rocha, A. Temperature and precipitation extremes over the iberian peninsula under climate change scenarios: A review. Climate 2021, 9, 139. [Google Scholar] [CrossRef]
- Ruosteenoja, K.; Markkanen, T.; Venäläinen, A.; Räisänen, P.; Peltola, H. Seasonal soil moisture and drought occurrence in Europe in CMIP5 projections for the 21st century. Clim. Dyn. 2018, 50, 1177–1192. [Google Scholar] [CrossRef] [Green Version]
- Calderan, A.; Sivilotti, P.; Braidotti, R.; Mihelčič, A.; Lisjak, K.; Vanzo, A. Managing moderate water deficit increased anthocyanin concentration and proanthocyanidin galloylation in “Refošk” grapes in Northeast Italy. Agric. Water Manag. 2021, 246, 106684. [Google Scholar] [CrossRef]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 2003, 1393–1405. [Google Scholar] [CrossRef]
- Jones, H.G. Interaction and integration of adaptive responses to water stress: The implications of an unpredictable environment. In Adaptation of Plants to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; Wiley: New York, NY, USA, 1980; pp. 353–365. [Google Scholar]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/Anisohydry: A Plant–Environment Interaction Rather Than a Simple Hydraulic Trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Morabito, C.; Orozco, J.; Tonel, G.; Cavalletto, S.; Meloni, G.R.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Do the ends justify the means? Impact of drought progression rate on stress response and recovery in Vitis vinifera. Physiol. Plant. 2022, 174, e13590. [Google Scholar] [CrossRef] [PubMed]
- Soar, C.J.; Speirs, J.; Maffei, S.M.; Penrose, A.B.; McCarthy, M.G.; Loveys, B.R. Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: Apparent links with ABA physiology and gene expression in leaf tissue. Aust. J. Grape Wine Res. 2006, 12, 2–12. [Google Scholar] [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- Dayer, S.; Scharwies, J.D.; Ramesh, S.A.; Sullivan, W.; Doerflinger, F.C.; Pagay, V.; Tyerman, S.D. Comparing Hydraulics between Two Grapevine Cultivars Reveals Differences in Stomatal Regulation under Water Stress and Exogenous ABA Applications. Front. Plant Sci. 2020, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Soar, C.J.; Collins, M.J.; Sadras, V.O. Irrigated Shiraz vines (Vitis vinifera) upregulate gas exchange and maintain berry growth in response to short spells of high maximum temperature in the field. Funct. Plant Biol. 2009, 36, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Faralli, M.; Bontempo, L.; Bianchedi, P.L.; Moser, C.; Bertamini, M.; Lawson, T.; Camin, F.; Stefanini, M.; Varotto, C. Natural variation in stomatal dynamics drives divergence in heat stress tolerance and contributes to seasonal intrinsic water-use efficiency in Vitis vinifera (subsp. sativa and sylvestris). J. Exp. Bot. 2022, 73, 3238–3250. [Google Scholar] [CrossRef] [PubMed]
- Lehr, P.P.; Hernández-Montes, E.; Ludwig-Müller, J.; Keller, M.; Zörb, C. Abscisic acid and proline are not equivalent markers for heat, drought and combined stress in grapevines. Aust. J. Grape Wine Res. 2022, 28, 119–130. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Rizhsky, L.; Liang, H.; Mittler, R. The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiol. 2002, 130, 1143–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poblete, T.; Ortega-Farías, S.; Moreno, M.A.; Bardeen, M. Artificial neural network to predict vine water status spatial variability using multispectral information obtained from an unmanned aerial vehicle (UAV). Sensors 2017, 17, 2488. [Google Scholar] [CrossRef] [Green Version]
- Diago, M.P.; Pou, A.; Millan, B.; Tardaguila, J.; Fernandes, A.M.; Melo-Pinto, P. Assessment of grapevine water status from hyperspectral imaging of leaves. Acta Hortic. 2014, 1038, 89–96. [Google Scholar] [CrossRef]
- Rapaport, T.; Hochberg, U.; Shoshany, M.; Karnieli, A.; Rachmilevitch, S. Combining leaf physiology, hyperspectral imaging and partial least squares-regression (PLS-R) for grapevine water status assessment. ISPRS J. Photogramm. Remote Sens. 2015, 109, 88–97. [Google Scholar] [CrossRef]
- Pagay, V.; Kidman, C.M. Evaluating Remotely-Sensed Grapevine (Vitis vinifera L.) Water Stress Responses across a Viticultural Region. Agronomy 2019, 9, 682. [Google Scholar] [CrossRef] [Green Version]
- Cogato, A.; Pagay, V.; Marinello, F.; Meggio, F.; Grace, P.; Migliorati, M.D.A. Assessing the feasibility of using sentinel-2 imagery to quantify the impact of heatwaves on irrigated vineyards. Remote Sens. 2019, 11, 2869. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.N.; Ferreira-Silva, S.L.; de Fontenele, A.V.; Ribeiro, R.V.; Viégas, R.A.; Silveira, J.A.G. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. J. Plant Physiol. 2010, 167, 1157–1164. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Gamon, J.A.; Penuelas, J.; Field, B. A Narrow-Waveband Spectral Index That Tracks Diurnal Changes in Photosynthetic Efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Courel, M.-F.; Chamard, P.; Guenegou, M.J.; Lerhun, J.; Levasseur, M.; Togola, M. Utilisation des bandes spectrales du vert et du rouge pour une meilleure évaluation des formations végétales actives. In Proceedings of the Congrès AUPELF-UREF, Sherbrooke, QC, Canada; 1991; pp. 203–210. [Google Scholar]
- Gitelson, A.A.; Merzlyak, M.N. Remote estimation of chlorophyll content in higher plant leaves. Int. J. Remote Sens. 1997, 18, 2691–2697. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L. Chlorophyll fluorescence effects on vegetation apparent reflectance: I. Leaf-level measurements and model simulation. Remote Sens. Environ. 2000, 74, 582–595. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Hunt, E.R.; Rock, B.N. Detection of changes in leaf water content using Near- and Middle-Infrared reflectances. Remote Sens. Environ. 1989, 30, 43–54. [Google Scholar]
- Penuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Save, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Idso, S.B. Non-water-stressed baselines: A key to measuring and interpreting plant water stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Hansen, P.M.; Jørgensen, J.R.; Thomsen, A. Predicting grain yield and protein content in winter wheat and spring barley using repeated canopy reflectance measurements and partial least squares regression. J. Agric. Sci. 2002, 139, 307–318. [Google Scholar] [CrossRef]
- Álvarez-Maldini, C.; Acevedo, M.; Pinto, M. Hydroscapes: A useful metric for distinguishing iso-/anisohydric behavior in almond cultivars. Plants 2021, 10, 1249. [Google Scholar] [CrossRef] [PubMed]
- Palliotti, A.; Poni, S.; Silvestroni, O.; Tombesi, S.; Bernizzoni, F. Morpho-structural and physiological performance of Sangiovese and Montepulciano cvv. (Vitis vinifera) under non-limiting water supply conditions. Funct. Plant Biol. 2011, 38, 888–898. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Cifre, J.; Escalona, J.M.; Galmés, J.; Gulías, J.; Lefi, E.K.; Martínez-Cañellas, S.F.; Moreno, M.T.; Ribas-Carbó, M.; et al. Understanding down-regulation of photosynthesis under water stress: Future prospects and searching for physiological tools for irrigation management. Ann. Appl. Biol. 2004, 144, 273–283. [Google Scholar] [CrossRef]
- Edwards, E.J.; Smithson, L.; Graham, D.C.; Clingeleffer, P.R. Grapevine canopy response to a high-temperature event during deficit irrigation. Aust. J. Grape Wine Res. 2011, 17, 153–161. [Google Scholar] [CrossRef]
- Greer, D.H.; Weston, C. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Funct. Plant Biol. 2010, 37, 206–214. [Google Scholar] [CrossRef]
- Prieto, J.A.; Lebon, É.; Ojeda, H. Stomatal behavior of different grapevine cultivars in response to soil water status and air water vapor pressure deficit. J. Int. Sci. Vigne du Vin 2010, 44, 9–20. [Google Scholar] [CrossRef]
- Carrasco-Benavides, M.; Antunez-Quilobrán, J.; Baffico-Hernández, A.; Ávila-Sánchez, C.; Ortega-Farías, S.; Espinoza, S.; Gajardo, J.; Mora, M.; Fuentes, S. Performance assessment of thermal infrared cameras of different resolutions to estimate tree water status from two cherry cultivars: An alternative to midday stem water potential and stomatal conductance. Sensors 2020, 20, 3596. [Google Scholar] [CrossRef] [PubMed]
- Maimaitiyiming, M.; Sagan, V.; Sidike, P.; Kwasniewski, M.T. Dual activation function-based Extreme Learning Machine (ELM) for estimating grapevine berry yield and quality. Remote Sens. 2019, 11, 740. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Keydan, G.P.; Merzlyak, M.N. Three-band model for noninvasive estimation of chlorophyll, carotenoids, and anthocyanin contents in higher plant leaves. Geophys. Res. Lett. 2006, 33, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Suárez, L.; Zarco-Tejada, P.J.; Sepulcre-Cantó, G.; Pérez-Priego, O.; Miller, J.R.; Jiménez-Muñoz, J.C.; Sobrino, J. Assessing canopy PRI for water stress detection with diurnal airborne imagery. Remote Sens. Environ. 2008, 112, 560–575. [Google Scholar] [CrossRef]
- Bhagat, V.; Kada, A.; Kumar, S. Analysis of Remote Sensing based Vegetation Indices (VIs) for Unmanned Aerial System (UAS): A Review. Remote Sens. Land 2020, 3, 58–73. [Google Scholar] [CrossRef]
- Nguyen, C.; Sagan, V.; Maimaitiyiming, M.; Maimaitijiang, M.; Bhadra, S.; Kwasniewski, M.T. Early detection of plant viral disease using hyperspectral imaging and deep learning. Sensors 2021, 21, 742. [Google Scholar] [CrossRef] [PubMed]
- Dobrowski, S.Z.; Pushnik, J.C.; Zarco-Tejada, P.J.; Ustin, S.L. Simple reflectance indices track heat and water stress-induced changes in steady-state chlorophyll fluorescence at the canopy scale. Remote Sens. Environ. 2005, 97, 403–414. [Google Scholar] [CrossRef]
- Pôças, I.; Gonçalves, J.; Costa, P.M.; Gonçalves, I.; Pereira, L.S.; Cunha, M. Hyperspectral-based predictive modelling of grapevine water status in the Portuguese Douro wine region. Int. J. Appl. Earth Obs. Geoinf. 2017, 58, 177–190. [Google Scholar] [CrossRef]
- Kováč, D.; Veselovská, P.; Klem, K.; Večeřová, K.; Ač, A.; Peñuelas, J.; Urban, O. Potential of photochemical reflectance index for indicating photochemistry and light use efficiency in leaves of European beech and Norway spruce trees. Remote Sens. 2018, 10, 1202. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Yao, X.; Liu, H.; Liu, B.; Cheng, T.; Tian, Y.; Cao, W.; Zhu, Y. Comparison of the abilities of vegetation indices and photosynthetic parameters to detect heat stress in wheat. Agric. For. Meteorol. 2019, 265, 121–136. [Google Scholar] [CrossRef]
- Ryu, J.H.; Jeong, H.; Cho, J. Performances of vegetation indices on paddy rice at elevated air temperature, heat stress, and herbicide damage. Remote Sens. 2020, 12, 2654. [Google Scholar] [CrossRef]
- Ripullone, F.; Rivelli, A.R.; Baraldi, R.; Guarini, R.; Guerrieri, R.; Magnani, F.; Peñuelas, J.; Raddi, S.; Borghetti, M. Effectiveness of the photochemical reflectance index to track photosynthetic activity over a range of forest tree species and plant water statuses. Funct. Plant Biol. 2011, 38, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarco-Tejada, P.J.; González-Dugo, V.; Williams, L.E.; Suárez, L.; Berni, J.A.J.; Goldhamer, D.; Fereres, E. A PRI-based water stress index combining structural and chlorophyll effects: Assessment using diurnal narrow-band airborne imagery and the CWSI thermal index. Remote Sens. Environ. 2013, 138, 38–50. [Google Scholar] [CrossRef]
- Król, A.; Amarowicz, R.; Weidner, S. Changes in the composition of phenolic compounds and antioxidant properties of grapevine roots and leaves (Vitis vinifera L.) under continuous of long-term drought stress. Acta Physiol. Plant. 2014, 36, 1491–1499. [Google Scholar] [CrossRef] [Green Version]
- Manley, P.V.; Sagan, V.; Fritschi, F.B.; Burken, J.G. Remote sensing of explosives-induced stress in plants: Hyperspectral imaging analysis for remote detection of unexploded threats. Remote Sens. 2019, 11, 1827. [Google Scholar] [CrossRef] [Green Version]
- Serrano, L.; González-Flor, C.; Gorchs, G. Assessing vineyard water status using the reflectance based Water Index. Agric. Ecosyst. Environ. 2010, 139, 490–499. [Google Scholar] [CrossRef]
- Peddinti, S.R.; Kambhammettu, B.V.N.P.; Rodda, S.R.; Thumaty, K.C.; Suradhaniwar, S. Dynamics of Ecosystem Water Use Efficiency in Citrus Orchards of Central India Using Eddy Covariance and Landsat Measurements. Ecosystems 2020, 23, 511–528. [Google Scholar] [CrossRef]
- Cogato, A.; Wu, L.; Jewan, S.Y.Y.; Meggio, F.; Marinello, F.; Sozzi, M.; Pagay, V. Evaluating the spectral and physiological responses of grapevines (Vitis vinifera L.) to heat and water stresses under different vineyard cooling and irrigation strategies. Agronomy 2021, 11, 1940. [Google Scholar] [CrossRef]
- Schillaci, C.; Perego, A.; Valkama, E.; Märker, M.; Saia, S.; Veronesi, F.; Lipani, A.; Lombardo, L.; Tadiello, T.; Gamper, H.A.; et al. New pedotransfer approaches to predict soil bulk density using WoSIS soil data and environmental covariates in Mediterranean agro-ecosystems. Sci. Total Environ. 2021, 780, 146609. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Variety | Average Volumetric Water Content (%) | No. of Vines (n) |
|---|---|---|---|
| GR_WW (control for Grenache) | Grenache | Plants well-watered (VWC = 10.5%) | 3 |
| SH_WW (control for Shiraz) | Shiraz | Plants well-watered (VWC = 7.8%) | 3 |
| GR_WS | Grenache | Plants subjected to WS (VWC = 2.4%) | 3 |
| SH_WS | Shiraz | Plants subjected to WS (VWC = 2.3%) | 3 |
| VI | Acronym | Equation | Class | Reference |
|---|---|---|---|---|
| Anthocyanin (Gitelson) | AntGitelson | AntGitelson = (1/R550 − 1/R700) × R780 | Pigment | [26] |
| Photochemical Reflectance Index | PRI | PRI = (R531 − R570)/(R531 + R570) | Pigment | [27] |
| Greenness Index | GI | GI = R554/R677 | Structure | [28] |
| Green Normalised Difference Vegetation Index | GNDVI | GNDVI = (R750 − R540 + R570)/(R750 + R540 − R570) | Structure | [29] |
| Fluorescence Curvature Index | FCI | FCI = R683/(R675 × R691) | Physiology | [30] |
| Modified Red-Edge Simple Ratio Index | MRESR | MRESR = (R750 − R445)/(R705 + R445) | Physiology | [31] |
| Normalised Difference Infrared Index | NDII | NDII = (R820 – R1650)/(R820 + R1650) | Water content | [32] |
| Water Index | WI | WI = R900/R970 | Water content | [33] |
| Grenache | Shiraz | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| stem | Pn | gs | E | WUEi | stem | Pn | gs | E | WUEi | |
| Whole canopy CWSI | −0.61 * | −0.32 ns | −0.31 ns | −0.38 ns | 0.60 * | −0.82 *** | −0.70 ** | −0.78 ** | −0.83 *** | 0.88 *** |
| Bottom canopy CWSI | −0.60 * | −0.32 ns | −0.33 ns | −0.39 ns | 0.62 * | −0.81 *** | −0.69 ** | −0.78 ** | −0.83 *** | 0.88 *** |
| Top canopy CWSI | −0.61 * | −0.31 ns | −0.28 ns | −0.35 ns | 0.56 ns | −0.80 ** | −0.68 ** | −0.77 ** | −0.81 *** | 0.87 *** |
| Physiological Parameter (Y) | Spectral Index (X) | Grenache | Shiraz | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| stem (MPa) | Model No. | Model type | Equations | R2 | RMSE | MAE | Significance | Model type | Equations | R2 | RMSE | MAE | Significance | |
| AntGitelson | 1 | Linear | Y = −6.95 − 11.49 * X | 0.36 | 2.21 | 1.49 | * | Linear | Y = −8.42 − 12.18 * X | 0.61 | 2.01 | 1.39 | ** | |
| PRI | 2 | Linear | Y = −6.35 + 89.85 * X | 0.45 | 2.04 | 1.46 | ** | Linear | Y = −7.91 + 116.6 * X | 0.69 | 1.70 | 1.23 | ** | |
| GI | 3 | Reciprocal-X | Y = 2.91 − 21.66/X | 0.44 | 2.06 | 1.52 | ** | ns | ns | |||||
| GNDVI | 4 | ns | ns | Square root-X | Y = −30.97 + 34.50 * sqrt(X) | 0.33 | 2.77 | 2.10 | ** | |||||
| FCI | 5 | ns | ns | Reciprocal-X | Y = 11.32 − 16.29/X | 0.28 | 2.88 | 2.17 | * | |||||
| MRESR | 6 | Reciprocal-X | Y = 0.46 − 7.42/X | 0.27 | 2.43 | 1.53 | * | Reciprocal-X | Y = 4.20 − 15.22/X | 0.55 | 2.28 | 1.63 | ** | |
| NDII | 7 | Reciprocal-X | Y = −4.33 − 0.078/X | 0.56 | 1.94 | 0.46 | ** | Linear | Y = −11.63 + 56.19 * X | 0.59 | 2.16 | 1.73 | ** | |
| WI | 8 | Reciprocal-X | Y = 278.2 − 285.3/X | 0.63 | 1.88 | 0.43 | ** | Reciprocal-X | Y = 376.1 − 383.3/X | 0.73 | 2.05 | 1.61 | ** | |
| Pn (μmol CO2 m−2 s−1) | AntGitelson | 1 | Exponential | Y = exp(1.29 − 3.78 * X) | 0.56 | 0.48 | 0.34 | ** | Linear | Y = 6.69 − 7.71 * X | 0.67 | 1.15 | 0.97 | ** |
| PRI | 2 | Exponential | Y = exp(1.49 + 27.74 * X) | 0.62 | 0.44 | 0.32 | ** | Exponential | Y = exp(1.90 + 11.31 * X) | 0.68 | 0.19 | 0.15 | ** | |
| GI | 3 | Double reciprocal | Y = 1/(−0.61 + 2.15/X) | 0.40 | 0.52 | 0.36 | ** | ns | ns | |||||
| GNDVI | 4 | S-curve | Y = exp(2.83 − 0.47/X) | 0.24 | 0.63 | 0.42 | * | Reciprocal-Y square root-X | Y = 1/(0.59 − 0.64 * sqrt(X)) | 0.32 | 0.05 | 0.04 | ** | |
| FCI | 5 | ns | ns | Double reciprocal | Y = 1/(−0.19 + 0.30/X) | 0.25 | 0.06 | 0.04 | * | |||||
| MRESR | 6 | S-curve | Y = exp(3.62 − 2.35/X) | 0.39 | 0.57 | 0.37 | ** | Double reciprocal | Y = 1/(−0.063 + 0.28/X) | 0.56 | 0.04 | 0.03 | ** | |
| NDII | 7 | Double reciprocal | Y = 1/(0.12 + 0.007/X) | 0.53 | 0.46 | 0.34 | ** | Exponential | Y = exp(1.56 + 5.11 * X) | 0.44 | 0.26 | 0.22 | ** | |
| WI | 8 | Double reciprocal | Y = 1/(−26.40 + 26.78/WI) | 0.46 | 0.51 | 0.36 | ** | Double reciprocal | Y = 1/(−6.07 + 6.22/X) | 0.46 | 0.05 | 0.04 | ** | |
| gs (mol H2O m−2 s−1) | AntGitelson | 1 | Exponential | Y = exp(−1.68 − 4.81 * X) | 0.56 | 0.61 | 0.47 | ** | Linear | Y = 0.35 − 0.72 * X | 0.70 | 0.36 | 0.28 | ** |
| PRI | 2 | Exponential | Y = exp(−1.43 + 36.13 * X) | 0.65 | 0.54 | 0.39 | ** | Exponential | Y = exp(−1.13 + 22.00 * X) | 0.72 | 0.34 | 0.27 | ** | |
| GI | 3 | Double reciprocal | Y = 1/(−20.27 + 63.38/X) | 0.44 | 6.04 | 4.62 | ** | ns | ns | |||||
| GNDVI | 4 | S-curve | Y = exp(0.39 − 0.63/X) | 0.27 | 0.79 | 0.54 | * | Reciprocal Y Logarithmic X | Y = 1/(−4.51 − 10.36 * ln(X)) | 0.36 | 2.40 | 1.83 | ** | |
| FCI | 5 | ns | ns | S-curve | Y = exp(2.54 − 3.11/X) | 0.27 | 0.56 | 0.48 | * | |||||
| MRESR | 6 | S-curve | Y = exp(1.41 − 3.14/X) | 0.45 | 0.70 | 0.44 | ** | Double reciprocal | Y = 1/(−7.04 + 13.78/X) | 0.61 | 1.87 | 1.32 | ** | |
| NDII | 7 | Multiplicative | Y = exp(0.81 + 0.80 * ln(X)) | 0.60 | 0.59 | 0.47 | ** | Exponential | Y = exp(−1.89 + 11.31 * X) | 0.60 | 0.41 | 0.33 | ** | |
| WI | 8 | Exponential | Y = exp(−95.97 + 94.26 * X) | 0.58 | 0.78 | 0.64 | ** | Linear | Y = −21.15 + 21.57 * WI | 0.61 | 0.12 | 0.09 | ** | |
| E (mmol H2O m−2 s−1) | AntGitelson | 1 | Exponential | Y = exp(0.43 − 1.60 * X) | 0.54 | 0.21 | 0.17 | ** | Reciprocal-Y | Y = 1/(0.48 + 0.22 * X) | 0.54 | 0.04 | 0.03 | ** |
| PRI | 2 | Reciprocal Y | Y = 1/(0.63− 7.91 * X) | 0.60 | 0.13 | 0.11 | ** | Reciprocal-Y | Y = 1/(0.47 − 2.06 * X) | 0.59 | 0.04 | 0.03 | ** | |
| GI | 3 | Double reciprocal | Y = 1/(0.081 + 1.26/X) | 0.27 | 0.18 | 0.13 | * | ns | ns | |||||
| GNDVI | 4 | Square root-Y reciprocal-X | Y = (1.69 − 0.13/X)^2 | 0.26 | 0.17 | 0.12 | * | Double reciprocal | Y = 1/(0.29 + 0.079/X) | 0.26 | 0.06 | 0.05 | * | |
| FCI | 5 | ns | ns | ns | ns | |||||||||
| MRESR | 6 | Double square root | Y = (−0.042 + 1.27 * sqrt(X))^2 | 0.40 | 0.15 | 0.11 | * | Double reciprocal | Y = 1/(0.25 + 0.28/X) | 0.52 | 0.05 | 0.04 | ** | |
| NDII | 7 | Reciprocal-Y Logarithmic-X | Y = 1/(0.13 − 0.18 * ln(X)) | 0.58 | 0.14 | 0.11 | ** | Reciprocal-Y | Y = 1/(0.56 − 1.32 * X) | 0.77 | 0.03 | 0.03 | ** | |
| WI | 8 | Double reciprocal | Y = 1/(−20.97 + 21.66/X) | 0.62 | 0.13 | 0.09 | ** | Double reciprocal | Y = 1/(−7.96 + 8.41/X) | 0.77 | 0.03 | 0.02 | ** | |
| WUEi (μmol CO2 mol−1 H2O−1) | AntGitelson | 1 | Squared-Y | Y= sqrt(428.6 + 825.8 * X) | 0.37 | 155.21 | 118.76 | * | Squared-Y | Y = sqrt(590.1 + 1265.5 * X) | 0.59 | 236.12 | 164.53 | ** |
| PRI | 2 | Squared-Y | Y = sqrt(386.1 – 6757 * X) | 0.51 | 136.74 | 107.51 | ** | Squared-Y | Y = sqrt(536.7 – 12,151 * X) | 0.69 | 206.70 | 164.42 | ns | |
| GI | 3 | Squared-Y reciprocal-X | Y = sqrt(−264.3 + 1519.1/X) | 0.44 | 146.56 | 116.88 | ** | ns | ns | |||||
| GNDVI | 4 | Squared-Y reciprocal-X | Y = sqrt(18.81 + 128.2/X) | 0.25 | 168.97 | 136.89 | * | Squared-Y reciprocal-X | Y = sqrt(−663.9 + 524.8/X) | 0.38 | 290.11 | 242.07 | * | |
| FCI | 5 | ns | ns | Squared-Y reciprocal-X | Y = sqrt(−1509 + 1733/X) | 0.26 | 316.56 | 270.36 | * | |||||
| MRESR | 6 | Squared-Y reciprocal-X | Y = sqrt(−184.5 + 634.5/X) | 0.42 | 149.39 | 121.55 | * | Squared-Y reciprocal-X | Y = sqrt(−819.0 + 1708/X) | 0.63 | 224.28 | 182.19 | ** | |
| NDII | 7 | Squared-Y square root-X | Y = sqrt(856.0 − 1722.9 * sqrt(X)) | 0.76 | 96.08 | 80.05 | ** | Linear | Y = 31.32 − 140.2 * X | 0.67 | 4.45 | 3.42 | ** | |
| WI | 8 | Squared-Y reciprocal-X | Y = sqrt(−22,275 + 22,733/X) | 0.79 | 90.71 | 70.96 | ** | Reciprocal-X | Y = −893.3 + 913.6/X | 0.70 | 4.21 | 3.23 | ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cogato, A.; Jewan, S.Y.Y.; Wu, L.; Marinello, F.; Meggio, F.; Sivilotti, P.; Sozzi, M.; Pagay, V. Water Stress Impacts on Grapevines (Vitis vinifera L.) in Hot Environments: Physiological and Spectral Responses. Agronomy 2022, 12, 1819. https://doi.org/10.3390/agronomy12081819
Cogato A, Jewan SYY, Wu L, Marinello F, Meggio F, Sivilotti P, Sozzi M, Pagay V. Water Stress Impacts on Grapevines (Vitis vinifera L.) in Hot Environments: Physiological and Spectral Responses. Agronomy. 2022; 12(8):1819. https://doi.org/10.3390/agronomy12081819
Chicago/Turabian StyleCogato, Alessia, Shaikh Yassir Yousouf Jewan, Lihua Wu, Francesco Marinello, Franco Meggio, Paolo Sivilotti, Marco Sozzi, and Vinay Pagay. 2022. "Water Stress Impacts on Grapevines (Vitis vinifera L.) in Hot Environments: Physiological and Spectral Responses" Agronomy 12, no. 8: 1819. https://doi.org/10.3390/agronomy12081819