
Investigation of Two QTL Conferring Seedling Resistance to Fusarium Crown Rot in Barley on Reducing Grain Yield Loss under Field Environments

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Inoculum Preparation
2.3. Conditions of Trial Sites
2.4. Trial Design
2.5. Data Collection
2.6. Statistical Analysis
3. Results
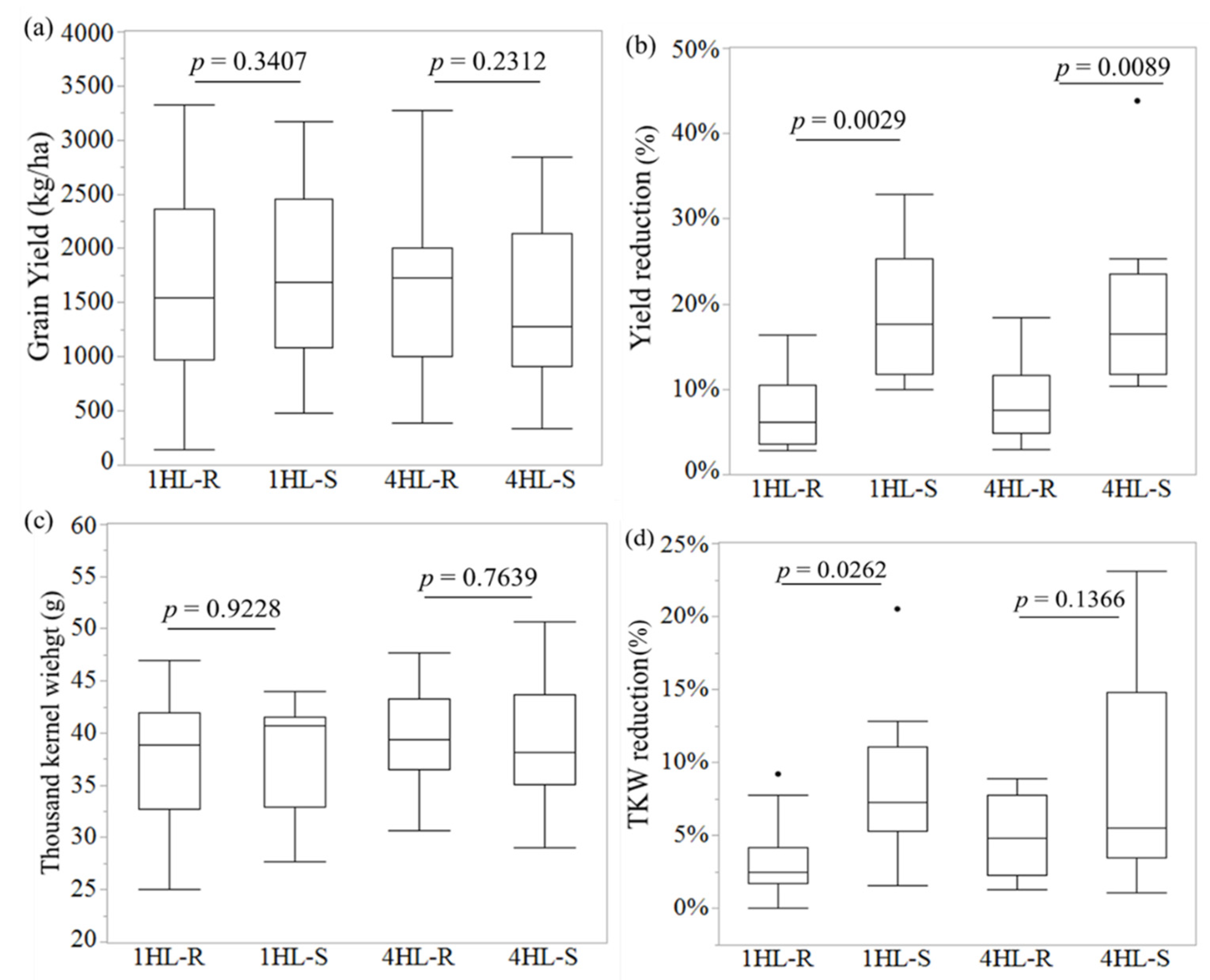
3.1. Field Performance of the near Isogenic Lines
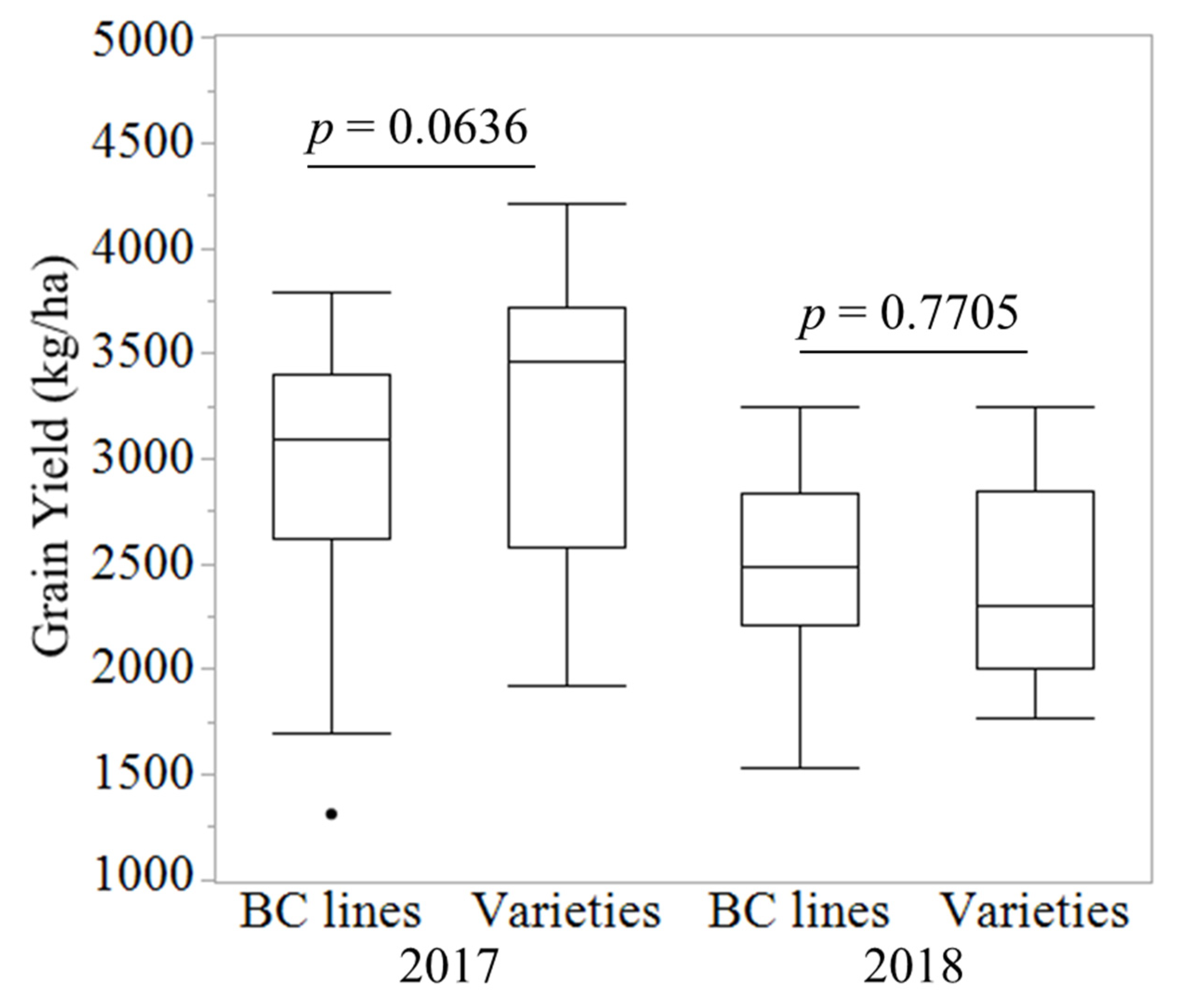
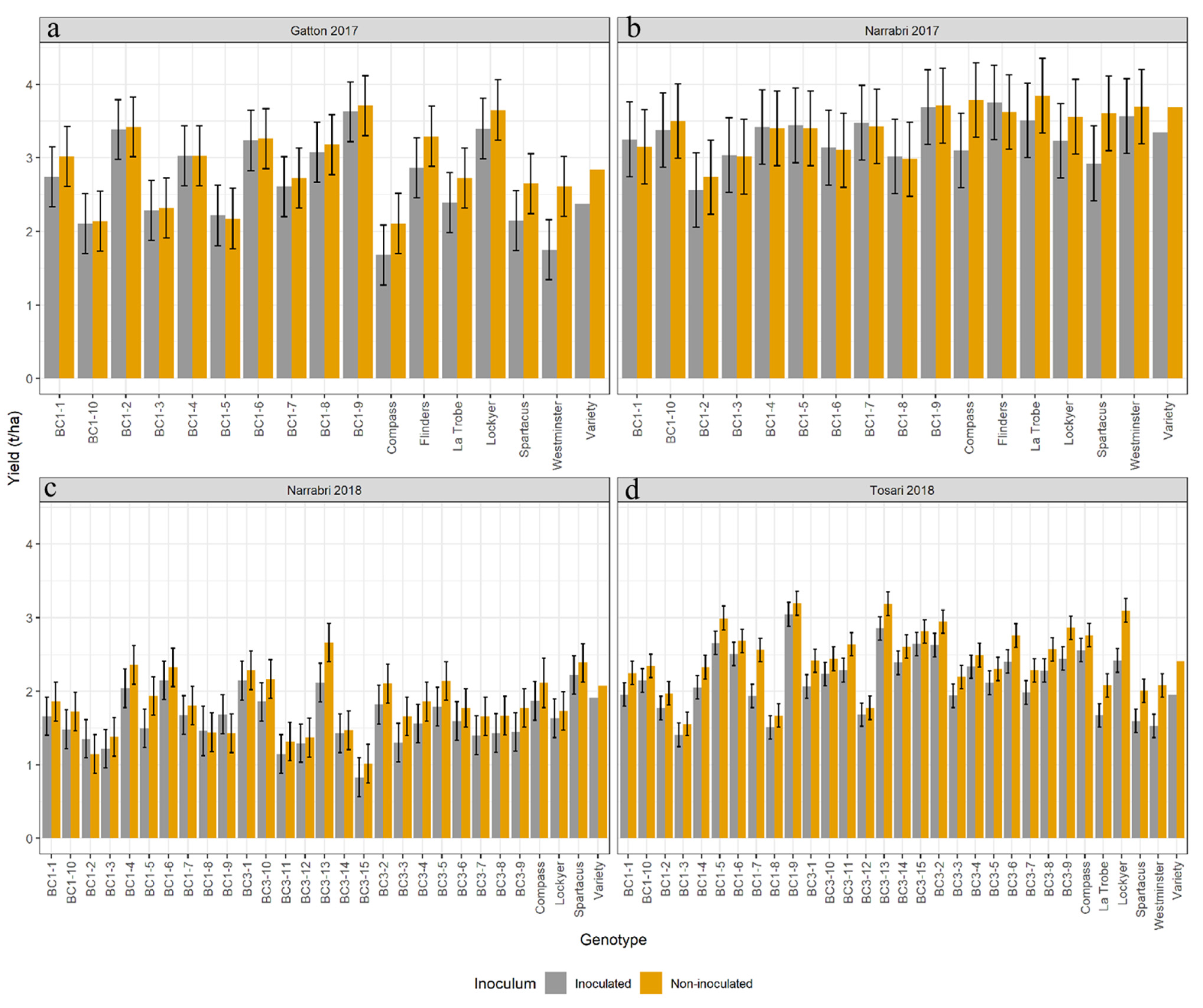
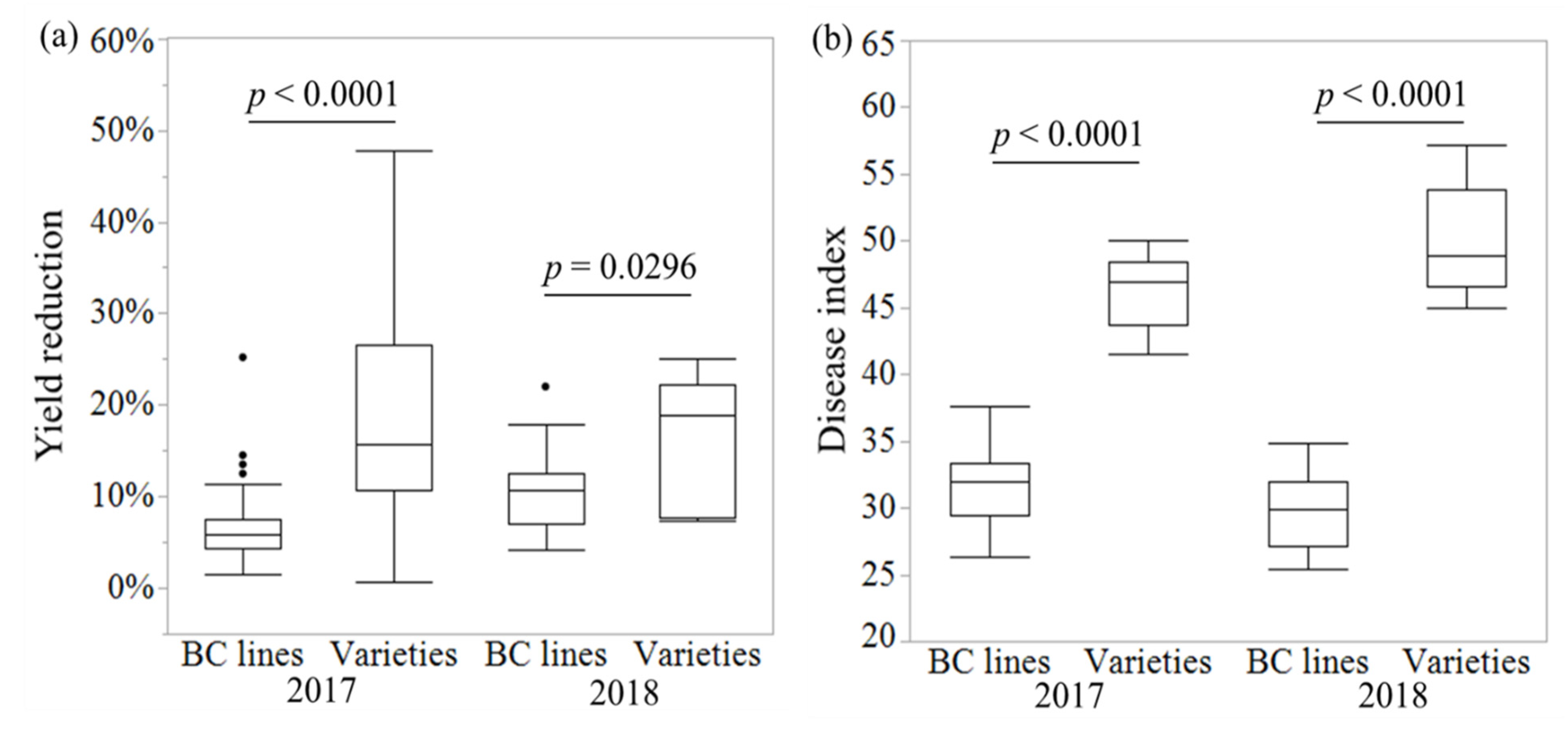
3.2. Field Performance of the BC Lines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Murray, G.M.; Brennan, J.P. Estimating disease losses to the Australian wheat industry. Australas. Plant Pathol. 2009, 38, 558–570. [Google Scholar] [CrossRef]
- Murray, G.M.; Brennan, J.P. Estimating disease losses to the Australian barley industry. Australas. Plant Pathol. 2010, 39, 85–96. [Google Scholar] [CrossRef]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.-M.; Whittaker, R.G. Crop Damage Estimates for Crown Rot of Wheat and Barley in the Pacific Northwest. Plant Dis. 2005, 89, 595–604. [Google Scholar] [CrossRef] [Green Version]
- Tunali, B.; Nicol, J.M.; Hodson, D.; Uçkun, Z.; Büyük, O.; Erdurmuş, D.; Hekimhan, H.; Aktaş, H.; Akbudak, M.A.; Bağci, S.A. Root and Crown Rot Fungi Associated with Spring, Facultative, and Winter Wheat in Turkey. Plant Dis. 2008, 92, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saremi, H.; Ammarellou, A.; Jafary, H. Incidence of crown rot disease caused by Fusarium pseudograminearum as a new soil born fungal species in North West Iran Pak. J. Biol. Sci. 2007, 20, 3606–3612. [Google Scholar]
- Chakraborty, S.; Liu, C.J.; Mitter, V.; Scott, J.B.; Akinsanmi, O.; Ali, S.; Dill-Macky, R.; Nicol, J.; Backhouse, D.; Simpfendorfer, S. Pathogen population structure and epidemiology are keys to wheat crown rot and Fusarium head blight management. Australas. Plant Pathol. 2006, 35, 643–655. [Google Scholar] [CrossRef]
- Matny, O.N.; Bates, S.T.; Song, Z. Original Article. Geographic distribution of Fusarium culmorum chemotypes associated with wheat crown rot in Iraq. J. Plant Prot. Res. 2017, 57, 43–49. [Google Scholar] [CrossRef]
- Minati, M.H. First record of nine Fusarium spp. causing root rot on wheat (Triticum aestivum L.) in Iraq. AIP Conf. Proc. 2020, 2290, 020009. [Google Scholar] [CrossRef]
- Chakraborty, S.; Obanor, F.; Westecott, R.; Abeywickrama, K. Wheat Crown Rot Pathogens Fusarium graminearum and F. pseudograminearum Lack Specialization. Phytopathology 2010, 100, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Hameed, M.; Rana, R.; Ali, Z. Identification and characterization of a novel Iraqi isolate of Fusarium pseudograminearum causing crown rot in wheat. Genet. Mol. Res. 2012, 11, 1341–1348. [Google Scholar] [CrossRef]
- Li, H.L.; He, X.L.; Ding, S.L.; Yuan, H.X.; Chen, L.L. First Report of Fusarium culmorum Causing Crown Rot of Wheat in China. Plant Dis. 2016, 100, 2532. [Google Scholar] [CrossRef]
- Xu, F.; Song, Y.L.; Wang, J.M.; Liu, L.L.; Zhao, K. Occurrence of Fusarium Crown Rot Caused by Fusarium pseudograminearum on Barley in China. Plant Dis. 2017, 101, 837. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, J.; Yan, W.; Yan, G.; Zhou, M.; Wei, Y.; Zheng, Y.; Liu, C. Different Tolerance in Bread Wheat, Durum Wheat and Barley to Fusarium Crown Rot Disease Caused by Fusarium pseudograminearum. J. Phytopathol. 2012, 160, 412–417. [Google Scholar] [CrossRef]
- Purss, G. Pathogenic specialization in Fusarium graminearum. Aust. J. Agric. Res. 1971, 22, 553–561. [Google Scholar] [CrossRef]
- Liu, C.; Ogbonnaya, F.C. Resistance to Fusarium crown rot in wheat and barley: A review. Plant Breed. 2015, 134, 365–372. [Google Scholar] [CrossRef]
- Akinsanmi, O.; Mitter, V.; Simpfendorfer, S.; Backhouse, D.; Chakraborty, S. Identity and pathogenicity of Fusarium spp. isolated from wheat fields in Queensland and northern New South Wales. Aust. J. Agric. Res. 2004, 55, 97–107. [Google Scholar] [CrossRef]
- Dodman, R.; Wildermuth, G. Inoculation methods for assessing resistance in wheat to crown rot caused by Fusarium graminearum Group 1. Crop Pasture Sci. 1987, 38, 473–486. [Google Scholar]
- Burgess, L.W.; Backhouse, D.; Summerell, B.A.; Swan, L.J. Crown Rot of Wheat, Fusarium: Paul E; Nelson Memorial Symposium; APS Press: St. Paul, MN, USA, 2001; pp. 1–294. [Google Scholar]
- Cook, R.J. Untold Stories: Forty Years of Field Research on Root Diseases of Wheat; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2017. [Google Scholar] [CrossRef]
- Smiley, R.W. Fusarium Crown Rot Whitehead Symptom as Influenced by Wheat Crop Management and Sampling Date. Plant Dis. 2019, 103, 2612–2623. [Google Scholar] [CrossRef]
- Summerell, B.A.; Leslie, J.F.; Backhouse, D.; Bryden, W.L.; Burgess, L.W. Fusarium: Paul E. Nelson Memorial Symposium; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2001. [Google Scholar]
- Smiley, R.W. Water and Temperature Parameters Associated with Winter Wheat Diseases Caused by Soilborne Pathogens. Plant Dis. 2009, 93, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.X.; Zheng, Y.L.; Wei, Y.; Zhou, M.X.; Liu, C.J. Genotypic differences to crown rot caused by Fusarium pseudograminearum in barley (Hordeum vulgare L.). Plant Breed. 2012, 131, 728–732. [Google Scholar] [CrossRef]
- Chen, G.D.; Liu, Y.X.; Wei, Y.M.; McIntyre, C.L.; Zhou, M.X.; Zheng, Y.-L.; Liu, C.J. Major QTL for Fusarium crown rot resistance in a barley landrace. Theor. Appl. Genet. 2013, 126, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Zhou, M.X.; Liu, C.J. A major QTL conferring crown rot resistance in barley and its association with plant height. Theor. Appl. Genet. 2009, 118, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, Y.; Ma, J.; Zheng, Z.; Wei, Y.; McIntyre, C.L.; Zheng, Y.-L.; Liu, C. A Novel and Major Quantitative Trait Locus for Fusarium Crown Rot Resistance in a Genotype of Wild Barley (Hordeum spontaneum L.). PLoS ONE 2013, 8, e58040. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Zheng, Z.; Hu, H.; Shi, H.; Ma, J.; Liu, Y.; Wei, Y.; Zheng, Y.-L.; Zhou, M.; Liu, C. A Novel QTL Conferring Fusarium Crown Rot Resistance Located on Chromosome Arm 6HL in Barley. Front. Plant Sci. 2019, 10, 1206. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Zheng, Z.; Powell, J.; Habib, A.; Stiller, J.; Zhou, M.; Liu, C. Validation and delineation of a locus conferring Fusarium crown rot resistance on 1HL in barley by analysing transcriptomes from multiple pairs of near isogenic lines. BMC Genom. 2019, 20, 650. [Google Scholar] [CrossRef] [Green Version]
- Habib, A.; Shabala, S.; Shabala, L.; Zhou, M.; Liu, C. Near-isogenic lines developed for a major QTL on chromosome arm 4HL conferring Fusarium crown rot resistance in barley. Euphytica 2015, 209, 555–563. [Google Scholar] [CrossRef]
- Chen, G.; Habib, A.; Wei, Y.; Zheng, Y.-L.; Shabala, S.; Zhou, M.; Liu, C. Enhancing Fusarium crown rot resistance by pyramiding large-effect QTL in barley. Mol. Breed. 2015, 35, 26. [Google Scholar] [CrossRef]
- Wildermuth, G.; McNamara, R. Testing wheat seedlings for resistance to crown rot caused by Fusarium graminearum group 1. Plant Dis. 1994, 10, 949–953. [Google Scholar] [CrossRef]
- Mitter, V.; Zhang, M.C.; Liu, C.J.; Ghosh, R.; Ghosh, M.; Chakraborty, S. A high-throughput glasshouse bioassay to detect crown rot resistance in wheat germplasm. Plant Pathol. 2006, 55, 433–441. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.; Chakraborty, S.; Manners, J.M.; Kazan, K. A Simple Method for the Assessment of Crown Rot Disease Severity in Wheat Seedlings Inoculated with Fusarium pseudograminearum. J. Phytopathol. 2008, 156, 751–754. [Google Scholar] [CrossRef]
- Yang, X.; Ma, J.; Li, H.; Ma, H.; Yao, J.; Liu, C. Different genes can be responsible for crown rot resistance at different developmental stages of wheat and barley. Eur. J. Plant Pathol. 2010, 128, 495–502. [Google Scholar] [CrossRef]
- Martin, A.; Bovill, W.D.; Percy, C.D.; Herde, D.; Fletcher, S.; Kelly, A.; Neate, S.M.; Sutherland, M.W. Markers for seedling and adult plant crown rot resistance in four partially resistant bread wheat sources. Theor. Appl. Genet. 2014, 128, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Tuinstra, M.R.; Ejeta, G.; Goldsbrough, P.B. Heterogeneous inbred family (HIF) analysis: A method for developing near-isogenic lines that differ at quantitative trait loci. Theor. Appl. Genet. 1997, 95, 1005–1011. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, H.B.; Chen, G.D.; Yan, G.; Liu, C.J. A procedure allowing up to eight generations of wheat and nine generations of barley per annum. Euphytica 2013, 191, 311–316. [Google Scholar] [CrossRef]
- Patterson, H.D.; Thompson, R. Recovery of inter-block information when block sizes are unequal. Biometrika 1971, 58, 545–554. [Google Scholar] [CrossRef]
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [Green Version]
- Butler, D.; Cullis, B.R.; Gilmour, A.; Gogel, B. ASReml-R Reference Manual The State of Queensland; Department of Primary Industries and Fisheries: Brisbane, Australia, 2009. [Google Scholar]
- R Development Core Team R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Ma, J.; Yan, G.; Liu, C.J. Development of near-isogenic lines for a major QTL on 3BL conferring Fusarium crown rot resistance in hexaploid wheat. Euphytica 2012, 183, 147–152. [Google Scholar] [CrossRef]
- Zheng, Z.; Gao, S.; Zhou, M.; Yan, G.; Liu, C. Enhancing Fusarium crown rot resistance by pyramiding large-effect QTL in common wheat (Triticum aestivum L.). Mol. Breed. 2017, 37, 107. [Google Scholar] [CrossRef]
- Gao, S.; Zheng, Z.; Hu, H.; Jiang, Y.; Liu, M.; Stiller, J.; Zhou, M.; Liu, C. Delineating a locus conferring Fusarium crown rot resistance on chromosome arm 1HL in barley by developing and analysing a large population derived from near isogenic lines. Crop J. 2020, 8, 1002–1010. [Google Scholar] [CrossRef]
- Jiang, Y.; Habib, A.; Zheng, Z.; Zhou, M.; Wei, Y.; Zheng, Y.-L.; Liu, C. Development of tightly linked markers and identification of candidate genes for Fusarium crown rot resistance in barley by exploiting a near-isogenic line-derived population. Theor. Appl. Genet. 2018, 132, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Xue, S.; Lin, F.; Yang, S.; Li, G.; Tang, M.; Kong, Z.; Cao, Y.; Zhao, D.; Jia, H.; et al. Mapping and validation of scab resistance QTLs in the Nanda2419 × Wangshuibai population. Cereal Res. Commun. 2008, 36, 245–251. [Google Scholar] [CrossRef]
- McCartney, C.A.; Somers, D.J.; Fedak, G.; Depauw, R.M.; Thomas, J.; Fox, S.L.; Humphreys, D.G.; Lukow, O.; Savard, M.E.; McCallum, B.D.; et al. The evaluation of FHB resistance QTLs introgressed into elite Canadian spring wheat germplasm. Mol. Breed. 2007, 20, 209–221. [Google Scholar] [CrossRef]
- Zheng, Z.; Kilian, A.; Yan, G.; Liu, C. QTL Conferring Fusarium Crown Rot Resistance in the Elite Bread Wheat Variety EGA Wylie. PLoS ONE 2014, 9, e96011. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.X.; Yang, X.M.; Ma, J.; Wei, Y.M.; Zheng, Y.L.; Ma, H.X.; Yao, J.B.; Yan, G.; Wang, Y.-G.; Manners, J.M.; et al. Plant Height Affects Fusarium Crown Rot Severity in Wheat. Phytopathology 2010, 100, 1276–1281. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Yan, W.; Liu, Y.; Wei, Y.; Zhou, M.; Zheng, Y.-L.; Manners, J.M.; Liu, C. The non-gibberellic acid-responsive semi-dwarfing gene uzu affects Fusarium crown rot resistance in barley. BMC Plant Biol. 2014, 14, 22. [Google Scholar] [CrossRef] [Green Version]
- Bai, Z.; Liu, C. Histological Evidence for Different Spread of Fusarium crown rot in Barley Genotypes with Different Heights. J. Phytopathol. 2015, 163, 91–97. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | d.f. | Grain Yield | TKW | DI | |||
|---|---|---|---|---|---|---|---|
| den. d.f. | p-Value | den. d.f. | p-Value | den. d.f. | p-Value | ||
| Trial (T) | 1 | 43.6 | 0.043 | 93.2 | <0.001 | 102.2 | 0.198 |
| Inoculation (I) | 1 | 98.3 | 0.042 | 90 | <0.001 | ||
| Locus (L) | 1 | 1.1 | 0.033 | 92.4 | 0.032 | 102.2 | 0.002 |
| T:I | 1 | 80.3 | 0.721 | 95.2 | 0.556 | ||
| L:Allele (A) | 2 | 39.6 | <0.001 | 90.7 | 0.008 | 102.2 | <0.001 |
| T:L | 1 | 69.1 | <0.001 | 90.9 | 0.494 | 102.2 | 0.68 |
| I:L | 1 | 86.5 | 0.779 | 84.4 | 0.719 | ||
| L:A:Genotype (G) | 16 | 15 | <0.001 | 90.1 | <0.001 | 102.2 | 0.826 |
| T:L:A | 2 | 84.6 | 0.115 | 92 | 0.081 | 102.2 | 0.616 |
| I:L:A | 2 | 85.8 | <0.001 | 81.3 | 0.001 | ||
| T:I:L | 1 | 86.1 | 0.61 | 81.6 | 0.973 | ||
| T:L:A:G | 16 | 85.1 | <0.001 | 90.1 | <0.001 | 102.2 | 1 |
| I:L:A:G | 16 | 85.6 | 0.951 | 81.8 | 0.034 | ||
| T:I:L:A | 2 | 86.3 | 0.41 | 81.5 | 0.1 | ||
| T:I:L:A:G | 16 | 85.3 | 0.584 | 80.9 | 0.33 | ||
| Genotype | Gatton | Narrabri | ||||
|---|---|---|---|---|---|---|
| DI | Difference (%) | p-Value | DI | Difference (%) | p-Value | |
| CR1H_1R | 33.2 | 28.0% | <0.01 | 29.3 | 24.5% | <0.01 |
| CR1H_1S | 46.1 | 38.8 | ||||
| CR1H_3R | 34.4 | 12.2% | <0.01 | 31.5 | 18.0% | <0.01 |
| CR1H_3S | 39.2 | 38.4 | ||||
| CR1H_7R | 29.3 | 27.4% | <0.01 | 29.3 | 16.8% | <0.01 |
| CR1H_7S | 40.4 | 35.3 | ||||
| CR1H_8R | 29.3 | 24.5% | <0.01 | 29.8 | 19.2% | <0.01 |
| CR1H_8S | 38.8 | 36.9 | ||||
| CR1H_9R | 28.9 | 24.9% | <0.01 | 29.3 | 22.9% | <0.01 |
| CR1H_9S | 38.4 | 38.1 | ||||
| CR4H_1R | 31.9 | 30.8% | <0.01 | 31.9 | 15.2% | <0.01 |
| CR4H_1S | 46.1 | 37.7 | ||||
| CR4H_3R | 31.9 | 27.1% | <0.01 | 31.9 | 23.1% | <0.01 |
| CR4H_3S | 43.9 | 41.6 | ||||
| CR4H_4R | 40.8 | 17.1% | <0.01 | 37.7 | 12.6% | <0.05 |
| CR4H_4S | 49.2 | 43.1 | ||||
| CR4H_6R | 39.2 | 15.7% | <0.01 | 37.3 | 11.1% | <0.05 |
| CR4H_6S | 46.5 | 41.9 | ||||
| CR4H_8R | 31.1 | 25.9% | <0.01 | 31.1 | 23.0% | <0.01 |
| CR4H_8S | 41.9 | 40.4 | ||||
| Term | d.f. | Grain Yield | TKW | DI | |||
|---|---|---|---|---|---|---|---|
| den. d.f. | p-Value | den. d.f. | p-Value | den. d.f. | p-Value | ||
| Trial (T) | 3 | 3.7 | 0.004 | 16.3 | <0.001 | 88.9 | 0.496 |
| Inoculation (I) | 1 | 2.8 | <0.001 | 154.5 | 0.132 | ||
| Genotype (G) | 30 | 43.5 | <0.001 | 56.5 | <0.001 | 47.6 | <0.001 |
| T:I | 3 | 11.2 | 0.003 | 83.6 | 0.235 | ||
| T:G | 56 | 73.4 | 0.001 | 98.3 | 0.438 | 89.6 | 0.074 |
| I:G | 30 | 28.8 | <0.001 | 105.3 | 0.198 | ||
| T:I:G | 56 | 52.6 | 0.002 | 73 | 0.722 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Z.; Powell, J.; Gao, S.; Percy, C.; Kelly, A.; Macdonald, B.; Zhou, M.; Davies, P.; Liu, C. Investigation of Two QTL Conferring Seedling Resistance to Fusarium Crown Rot in Barley on Reducing Grain Yield Loss under Field Environments. Agronomy 2022, 12, 1282. https://doi.org/10.3390/agronomy12061282
Zheng Z, Powell J, Gao S, Percy C, Kelly A, Macdonald B, Zhou M, Davies P, Liu C. Investigation of Two QTL Conferring Seedling Resistance to Fusarium Crown Rot in Barley on Reducing Grain Yield Loss under Field Environments. Agronomy. 2022; 12(6):1282. https://doi.org/10.3390/agronomy12061282
Chicago/Turabian StyleZheng, Zhi, Jonathan Powell, Shang Gao, Cassandra Percy, Alison Kelly, Bethany Macdonald, Meixue Zhou, Philip Davies, and Chunji Liu. 2022. "Investigation of Two QTL Conferring Seedling Resistance to Fusarium Crown Rot in Barley on Reducing Grain Yield Loss under Field Environments" Agronomy 12, no. 6: 1282. https://doi.org/10.3390/agronomy12061282