
Genetic Improvement of Heat Stress Tolerance in Cereal Crops



Abstract
:1. Introduction
2. Impact of Heat Stress on Cereal Crop Production
2.1. Cool-Season Cereal Crops: Wheat and Barley
2.2. Warm-Season Cereal Crops: Rice and Maize
3. Genetic Improvement of Heat Stress Tolerance—Lessons and Insights from Arabidopsis thaliana
3.1. Acquisition of Thermotolerance: Expression of Molecular Chaperones
3.2. Long-Term Adaptation to Heat Stress: Heat Stress Memory
4. Genetic and Physiological Basis of Agronomic Traits under Heat Stress
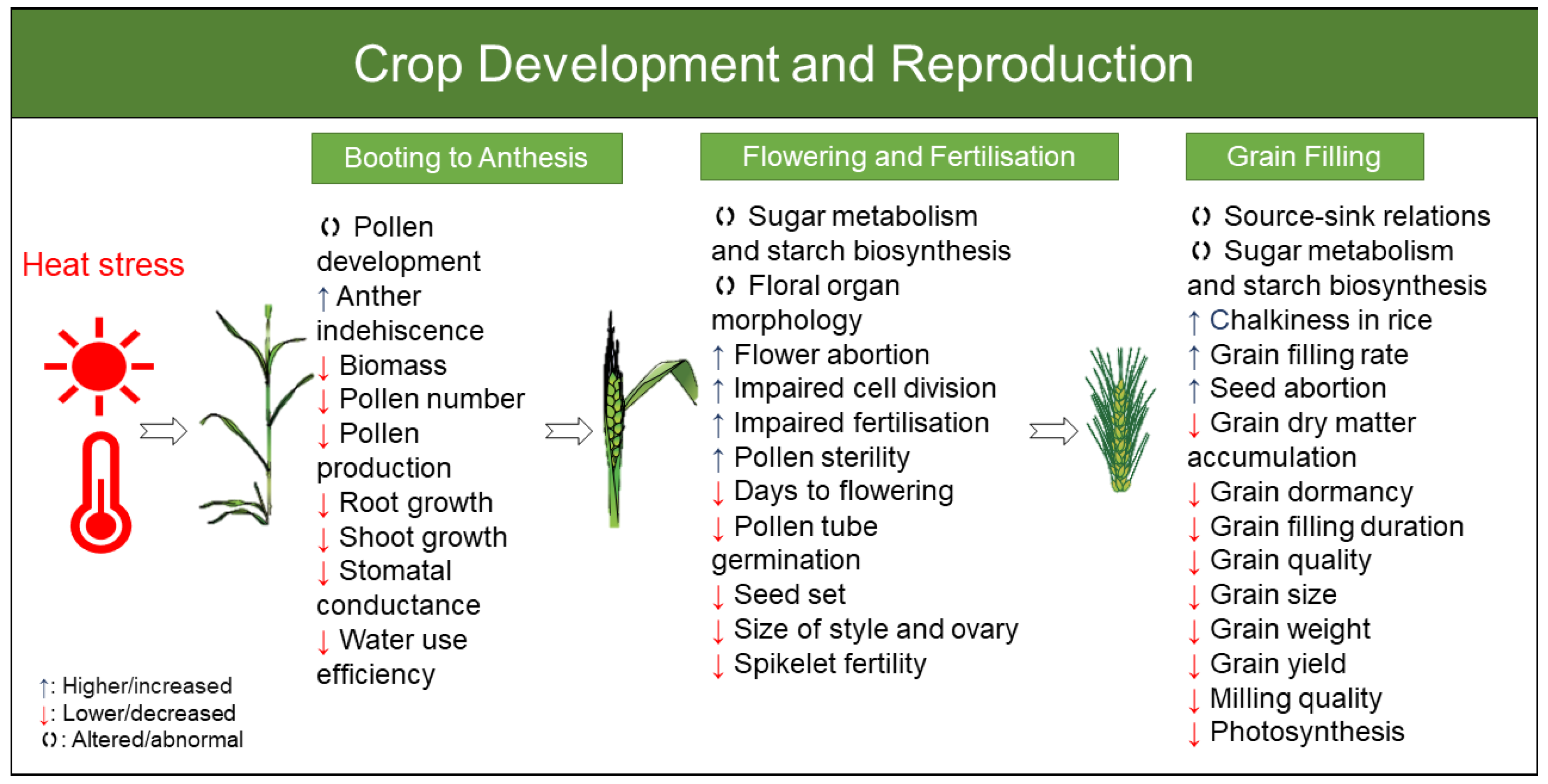
4.1. Crop Development and Reproduction
4.1.1. Wheat
4.1.2. Barley
4.1.3. Rice
4.1.4. Maize
4.2. Grain Yield and Quality
4.2.1. Wheat
4.2.2. Barley
4.2.3. Rice
4.2.4. Maize
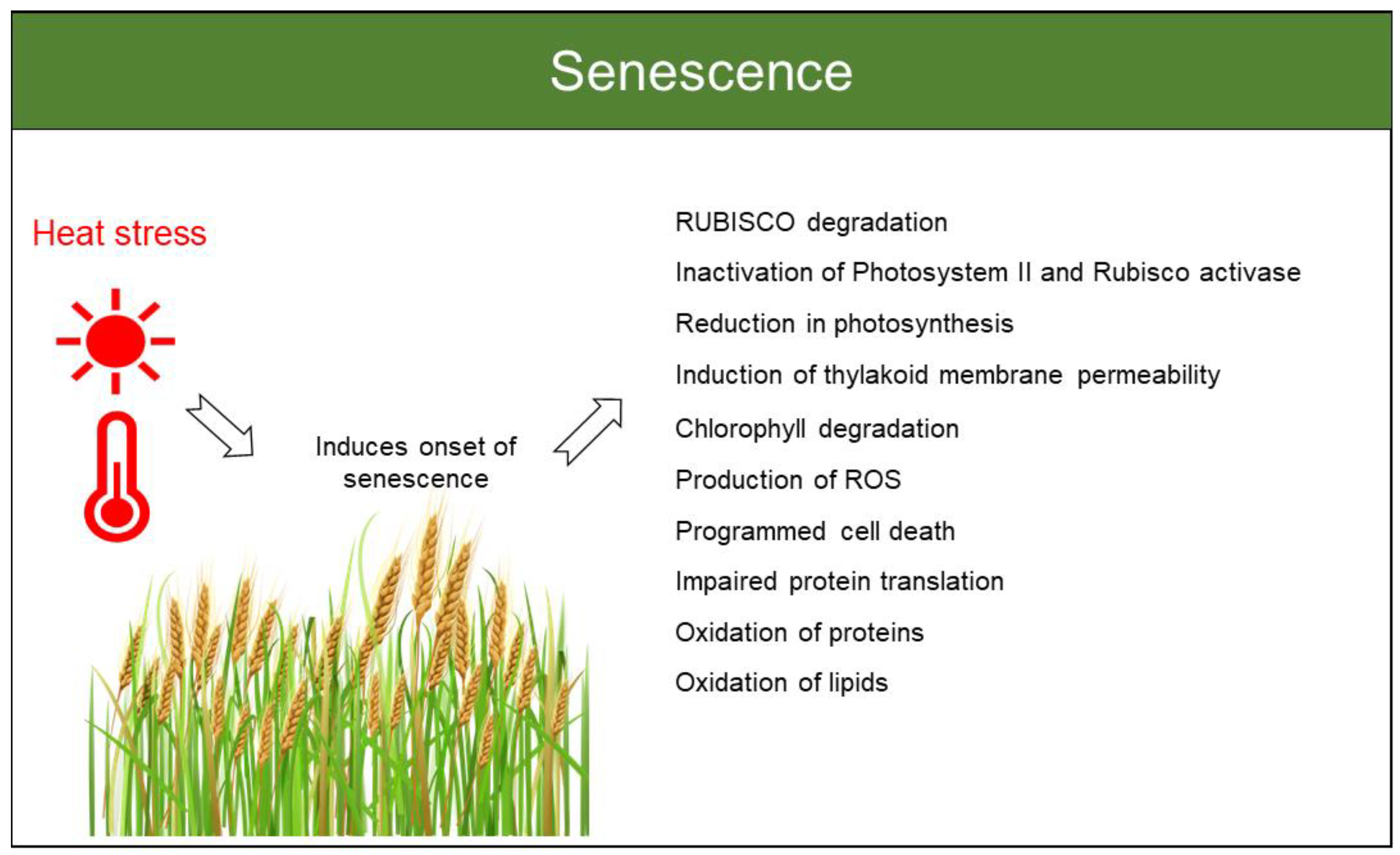
4.3. Senescence and Biomass
4.3.1. Wheat
4.3.2. Barley
4.3.3. Rice
4.3.4. Maize
5. Effective Methodology and Approaches for Genetic Improvement of Heat Stress Tolerance
5.1. Germplasm Screening in the Glasshouse and Field Trials
5.2. Genetic Mapping of Heat Stress Tolerance Genes
5.3. Targeted Manipulation of Heat Stress Tolerance Genes
6. Prospects and Challenges
- Heat tolerance in crops is a polygenic trait that is difficult to quantify, and thus different definitions of heat tolerance are currently used for field-based and laboratory evaluations. Until now, no direct method is available to select heat-tolerant plants, but some traits such as canopy temperature depression, membrane thermo-stability and chlorophyll fluorescence may be effective indicators of plant heat tolerance.
- Crops often experience several environmental stresses, such as drought, nutrient stress, and heat stress, simultaneously in the field. Therefore, many studies screen (deliberately or unintentionally) for the combined effect of different environmental stresses, and the interaction between stress responses, on the target traits. Past studies have used late sowing times [100,110] or in-field heat tents [174] to control the timing and incidence of heat stress. It remains unclear how comparable results obtained from these experiments are to heat events experienced in more typical production environments.
- Uncertainties in the knowledge of molecular crop responses to heat are limiting current research and breeding efforts. Despite considerable research to map QTLs and quantify their effects and interactions, most quantitative trait variation’s biological and molecular basis remains poorly understood. The information from QTL studies alone is often not sufficient to determine whether a genetic locus and a phenotype are connected directly, for example, via a specific gene or through multiple steps and pathways, such as transcription factors or other regulatory sequences. High-resolution mapping of specific genomic regions would help clarify whether coincident QTLs are due to linkage or pleiotropy.
- MAS and association genetics require long selection cycles and cannot capture significant marker–QTL associations with minor gene effects. Genomic selection is a technique that employs genome-wide markers to predict the phenotype of lines based on the genotype [175]. The marker effects are estimated in a fully genotyped and phenotyped training population and then used to obtain genomic estimated breeding values (GEBV) of the test population that has only been genotyped. The prediction’s reliability relies on the genetic diversity and size of the training population used to calibrate the prediction models, and the density of the molecular markers used to genotype lines. Genomic selection enables indirect selection for quantitative traits before phenotyping and has long been applied to animal breeding but has only a recent history in crop breeding. When used to enhance wheat’s heat and drought tolerance, genomic selection has shown that canopy temperature and normalized difference vegetation index (NDVI) measurements can increase genomic prediction model accuracies for grain yield [176]. A prediction accuracy of 0.62 using an item-based collaborative filtering approach was reported for grain yield in heat-stressed environments from NDVI measurements [5]. These results suggest a great potential for incorporating high-throughput phenotyping in genomic selection studies to improve the heat-stress tolerance of crops.
- Recently, regulatory connections between heat stress response pathways and the plant circadian clock have been revealed [177,178], opening new avenues for crop improvement if better understood. Adjustments in clock function may allow crops to re-adjust the balance between crop growth and stress tolerance in response to changes in the environment and ensure a sustained increase in heat stress tolerance when needed [179].
- Heatwave events have extremely high economic costs, but even with adjustments in sowing date, high temperatures are largely unavoidable during the reproductive phase of cereal crops in many parts of the world. Biotechnology, particularly genetic modifications and molecular markers, is key to speeding up plant breeding programs to deliver improved and better-adapted varieties sooner. Despite this, only small efforts have been made to identify genetic markers associated with heat tolerance that can be deployed in cereal crop breeding programs. There remains an urgent need to identify diagnostic markers for MAS breeding to maintain and improve the grain yield of cereal crops under heat-stressed environments.
- Regulatory hurdles hinder the development and adoption of new genetic variations with DNA technologies, mutational breeding, or genome editing. Added value contributed by the transgenes to enhancing stress tolerance must be demonstrated first before acceptance in breeding programs. This can only be achieved through costly large-scale extensive field evaluations of transgenic germplasm—a task difficult to accomplish in the current regulatory environment in many parts of the world. Given the regulatory complications and costs, the yield benefit under heat stress needs to be substantial without negative impacts in normal production environments. Argentina, one of the first countries with a regulatory framework for genetically modified crops, has recently approved the transgenic HB4 drought-tolerant wheat for growth and consumption [180]. Based on field trials conducted over ten years, transgenic HB4 wheat shows a 20% increased yield in seasons affected by drought with no yield penalty under normal growth conditions. HB4 technology is based on HaHB4, a sunflower transcription factor belonging to the homeodomain-leucine zipper I family whose ectopic expression in several plant species triggered drought tolerance. Given the recent advancements, it may only be a matter of time before the first heat-stress tolerant cereal crops will be commercially released.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhongming, Z.; Linong, L.; Xiaona, Y.; Wangqiang, Z.; Wei, L.; IPCC. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2021: The Physical Science Basis; Masson-Delmotte, V., Zhai, P.A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; in press. [Google Scholar]
- Ray, D.K.; Gerber, J.S.; MacDonald, G.K.; West, P.C. Climate Variation Explains a Third of Global Crop Yield Variability. Nat. Commun. 2015, 6, 5989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenov, M.A.; Shewry, P.R. Modelling Predicts That Heat Stress, Not Drought, Will Increase Vulnerability of Wheat in Europe. Sci. Rep. 2011, 1, 66. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature Increase Reduces Global Yields of Major Crops in Four Independent Estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Juliana, P.; Montesinos-López, O.A.; Crossa, J.; Mondal, S.; González Pérez, L.; Poland, J.; Huerta-Espino, J.; Crespo-Herrera, L.; Govindan, V.; Dreisigacker, S.; et al. Integrating Genomic-Enabled Prediction and High-Throughput Phenotyping in Breeding for Climate-Resilient Bread Wheat. Theor. Appl. Genet. 2019, 132, 177–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.; Tester, M.A.; Julkowska, M.M. The Use of High-Throughput Phenotyping for Assessment of Heat Stress-Induced Changes in Arabidopsis. Plant Phenomics 2020, 2020, 3723916. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, E.I.; Fischer, G.; van Velthuizen, H.; Walter, C.; Ewert, F. Global Hot-Spots of Heat Stress on Agricultural Crops Due to Climate Change. Agric. For. Meteorol. 2013, 170, 206–215. [Google Scholar] [CrossRef]
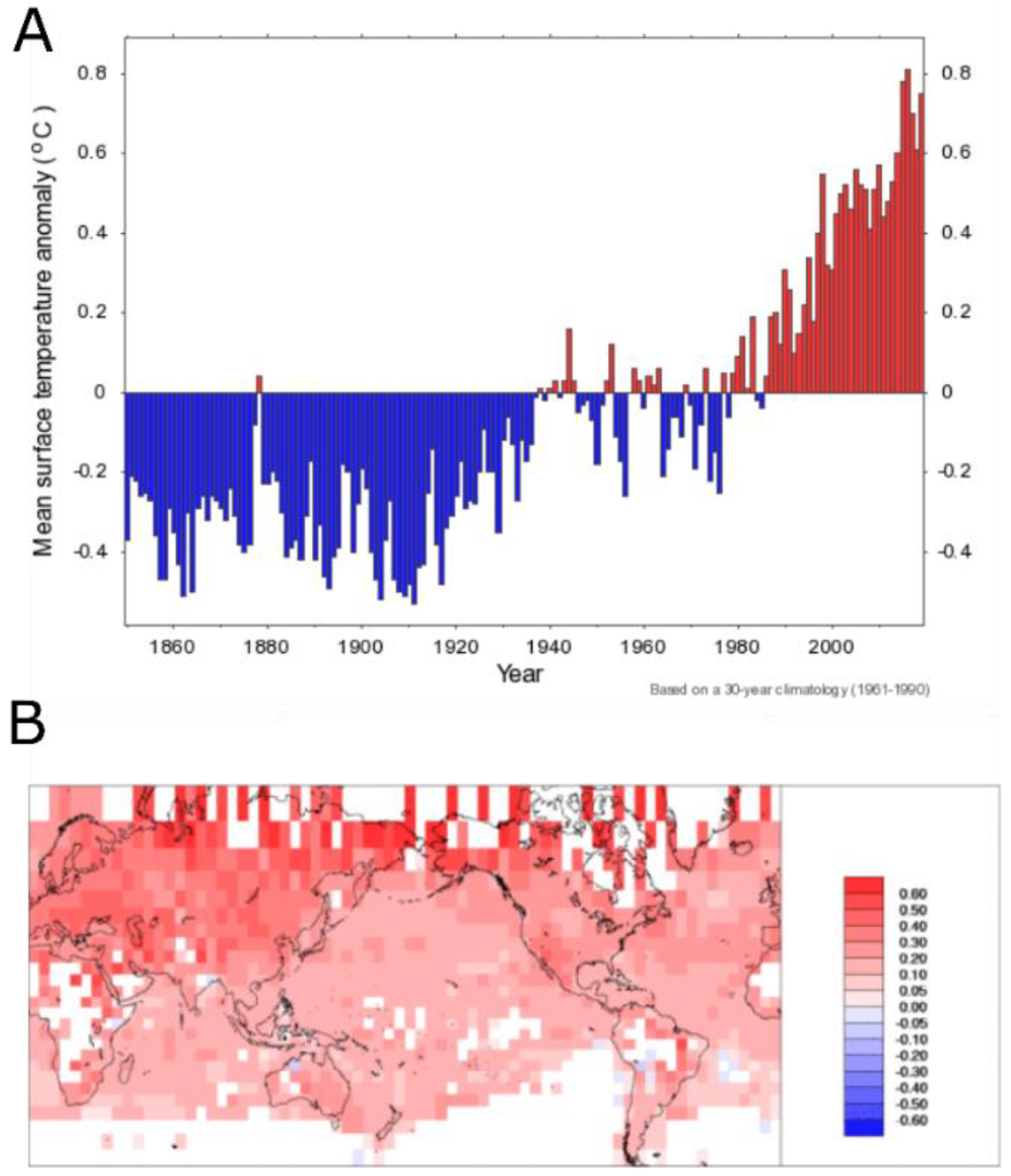
- Morice, C.P.; Kennedy, J.J.; Rayner, N.A.; Jones, P.D. Quantifying Uncertainties in Global and Regional Temperature Change Using an Ensemble of Observational Estimates: The HadCRUT4 Data Set. J. Geophys. Res. 2012, 117, D08101. [Google Scholar] [CrossRef]
- Jacott, C.N.; Boden, S.A. Feeling the Heat: Developmental and Molecular Responses of Wheat and Barley to High Ambient Temperatures. J. Exp. Bot. 2020, 71, 5740–5751. [Google Scholar] [CrossRef]
- Langridge, P.; Reynolds, M. Breeding for Drought and Heat Tolerance in Wheat. Theor. Appl. Genet. 2021, 134, 1753–1769. [Google Scholar] [CrossRef]
- Asseng, S.; Foster, I.; Turner, N.C. The Impact of Temperature Variability on Wheat Yields: Impact Of Temperature Variability On Wheat Yields. Glob. Chang. Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/ (accessed on 23 March 2022).
- Weichert, H.; Högy, P.; Mora-Ramirez, I.; Fuchs, J.; Eggert, K.; Koehler, P.; Weschke, W.; Fangmeier, A.; Weber, H. Grain Yield and Quality Responses of Wheat Expressing a Barley Sucrose Transporter to Combined Climate Change Factors. J. Exp. Bot. 2017, 68, 5511–5525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.; Yashveer, S.; Rehman, S.; Gyawali, S.; Kumar, Y.; Chao, S.; Sarker, A.; Verma, R.P.S. Genetic and Agro-Morphological Diversity in Global Barley (Hordeum vulgare L.) Collection at ICARDA. Genet. Resour. Crop. Evol. 2021, 68, 1315–1330. [Google Scholar] [CrossRef]
- Sánchez, B.; Rasmussen, A.; Porter, J.R. Temperatures and the Growth and Development of Maize and Rice: A Review. Glob. Chang. Biol. 2014, 20, 408–417. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S.; Maatougui, M.; Michael, M.; Slash, M.; Haghparast, R.; Rahmanian, M.; Taheri, A.; Al-Yassin, A.; Benbelkacem, A.; et al. Plant Breeding and Climate Changes. J. Agric. Sci. 2010, 148, 627–637. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B. On the Use of Statistical Models to Predict Crop Yield Responses to Climate Change. Agric. For. Meteorol. 2010, 150, 1443–1452. [Google Scholar] [CrossRef]
- Hill, C.B.; Li, C. Genetic Architecture of Flowering Phenology in Cereals and Opportunities for Crop Improvement. Front. Plant Sci. 2016, 7, 1906. [Google Scholar] [CrossRef] [Green Version]
- Woodward, A.W.; Bartel, B. Biology in Bloom: A Primer on the Arabidopsis Thaliana Model System. Genetics 2018, 208, 1337–1349. [Google Scholar] [CrossRef] [Green Version]
- Silva-Correia, J.; Freitas, S.; Tavares, R.M.; Lino-Neto, T.; Azevedo, H. Phenotypic Analysis of the Arabidopsis Heat Stress Response during Germination and Early Seedling Development. Plant Methods 2014, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Weng, M.; Yang, Y.; Feng, H.; Pan, Z.; Shen, W.-H.; Zhu, Y.; Dong, A. Histone Chaperone ASF1 Is Involved in Gene Transcription Activation in Response to Heat Stress in Arabidopsis Thaliana: AtASF1 in Heat Stress Response. Plant Cell. Environ. 2014, 37, 2128–2138. [Google Scholar] [CrossRef]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Heat Stress Response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Nover, L.; Bharti, K.; Döring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K.-D. Arabidopsis and the Heat Stress Transcription Factor World: How Many Heat Stress Transcription Factors Do We Need? Cell Stress Chaper. 2001, 6, 177. [Google Scholar] [CrossRef]
- Wiederrecht, G.; Seto, D.; Parker, C.S. Isolation of the Gene Encoding the S. Cerevisiae Heat Shock Transcription Factor. Cell 1988, 54, 841–853. [Google Scholar] [CrossRef]
- Charng, Y.; Liu, H.; Liu, N.; Chi, W.; Wang, C.; Chang, S.; Wang, T. A Heat-Inducible Transcription Factor, HsfA2, Is Required for Extension of Acquired Thermotolerance in Arabidopsis. Plant Physiol. 2007, 143, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Chen, S.; Liu, K.; Liu, Y.; Ni, L.; Zhang, K.; Zhang, L. Isolation of Heat Shock Factor HsfA1a-Binding Sites in Vivo Revealed Variations of Heat Shock Elements in Arabidopsis Thaliana. Plant Cell Physiol. 2008, 49, 1306–1315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.-C.; Liao, H.-T.; Charng, Y.-Y. The Role of Class A1 Heat Shock Factors (HSFA1s) in Response to Heat and Other Stresses in Arabidopsis: Master Heat Shock Factors in Arabidopsis. Plant Cell. Environ. 2011, 34, 738–751. [Google Scholar] [CrossRef]
- Wunderlich, M.; Werr, W.; Schoffl, F. Generation of Dominant-Negative Effects on the Heat Shock Response in Arabidopsis Thaliana by Transgenic Expression of a Chimaeric HSF1 Protein Fusion Construct. Plant J. 2003, 35, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Döring, P.; Vierling, E.; Scharf, K.-D. Complexity of the Heat Stress Response in Plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Schramm, F.; Larkindale, J.; Kiehlmann, E.; Ganguli, A.; Englich, G.; Vierling, E.; Von Koskull-Döring, P. A Cascade of Transcription Factor DREB2A and Heat Stress Transcription Factor HsfA3 Regulates the Heat Stress Response of Arabidopsis: Role of Arabidopsis HsfA3. Plant J. 2007, 53, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Qin, F.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional Analysis of an Arabidopsis Transcription Factor, DREB2A, Involved in Drought-Responsive Gene Expression. Plant Cell 2006, 18, 1292–1309. [Google Scholar] [CrossRef] [Green Version]
- Davletova, S.; Rizhsky, L.; Liang, H.; Shengqiang, Z.; Oliver, D.J.; Coutu, J.; Shulaev, V.; Schlauch, K.; Mittler, R. Cytosolic Ascorbate Peroxidase 1 Is a Central Component of the Reactive Oxygen Gene Network of Arabidopsis. Plant Cell 2005, 17, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Isono, K.; Sakata, Y.; Taji, T. Catalase2 Plays a Crucial Role in Long-Term Heat Tolerance of Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2021, 534, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, T.; Oberkofler, V.; Trindade, I.; Altmann, S.; Brzezinka, K.; Lämke, J.; Gorka, M.; Kappel, C.; Sokolowska, E.; Skirycz, A.; et al. Heteromeric HSFA2/HSFA3 Complexes Drive Transcriptional Memory after Heat Stress in Arabidopsis. Nat. Commun. 2021, 12, 3426. [Google Scholar] [CrossRef] [PubMed]
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A Hit-and-run Heat Shock Factor Governs Sustained Histone Methylation and Transcriptional Stress Memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, Q.; Lu, X.; Zeng, H.; Zhang, Y.; Zhu, J. Heat Stress Induction of MiR398 Triggers a Regulatory Loop That Is Critical for Thermotolerance in Arabidopsis. Plant J. 2013, 74, 840–851. [Google Scholar] [CrossRef]
- Sedaghatmehr, M.; Mueller-Roeber, B.; Balazadeh, S. The Plastid Metalloprotease FtsH6 and Small Heat Shock Protein HSP21 Jointly Regulate Thermomemory in Arabidopsis. Nat. Commun. 2016, 7, 12439. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A Chromosome Conformation Capture Ordered Sequence of the Barley Genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [Green Version]
- The International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; et al. Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Jayakodi, M.; Padmarasu, S.; Haberer, G.; Bonthala, V.S.; Gundlach, H.; Monat, C.; Lux, T.; Kamal, N.; Lang, D.; Himmelbach, A.; et al. The Barley Pan-Genome Reveals the Hidden Legacy of Mutation Breeding. Nature 2020, 588, 284–289. [Google Scholar] [CrossRef]
- Walkowiak, S.; Gao, L.; Monat, C.; Haberer, G.; Kassa, M.T.; Brinton, J.; Ramirez-Gonzalez, R.H.; Kolodziej, M.C.; Delorean, E.; Thambugala, D.; et al. Multiple Wheat Genomes Reveal Global Variation in Modern Breeding. Nature 2020, 588, 277–283. [Google Scholar] [CrossRef]
- Han, Y.; Broughton, S.; Liu, L.; Zhang, X.-Q.; Zeng, J.; He, X.; Li, C. Highly Efficient and Genotype-Independent Barley Gene Editing Based on Anther Culture. Plant Commun. 2021, 2, 100082. [Google Scholar] [CrossRef]
- Nishizawa-Yokoi, A.; Toki, S. A PiggyBac -mediated Transgenesis System for the Temporary Expression of CRISPR/Cas9 in Rice. Plant Biotechnol. J. 2021, 19, 1386–1395. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, J.; Ren, W.; Yang, Q.; Chai, Z.; Chen, R.; Wang, L.; Zhao, J.; Lang, Z.; Wang, H.; et al. Gene-Indexed Mutations in Maize. Mol. Plant 2018, 11, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Schreiber, M.; Barakate, A.; Uzrek, N.; Macaulay, M.; Sourdille, A.; Morris, J.; Hedley, P.E.; Ramsay, L.; Waugh, R. A Highly Mutagenised Barley (Cv. Golden Promise) TILLING Population Coupled with Strategies for Screening-by-Sequencing. Plant Methods 2019, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.-D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed Breeding Is a Powerful Tool to Accelerate Crop Research and Breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barmeier, G.; Schmidhalter, U. High-Throughput Field Phenotyping of Leaves, Leaf Sheaths, Culms and Ears of Spring Barley Cultivars at Anthesis and Dough Ripeness. Front. Plant Sci. 2017, 8, 1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madec, S.; Baret, F.; de Solan, B.; Thomas, S.; Dutartre, D.; Jezequel, S.; Hemmerlé, M.; Colombeau, G.; Comar, A. High-Throughput Phenotyping of Plant Height: Comparing Unmanned Aerial Vehicles and Ground LiDAR Estimates. Front. Plant Sci. 2017, 8, 2002. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Rahimi-Eichi, V.; Haefele, S.; Garnett, T.; Miklavcic, S.J. Estimation of Vegetation Indices for High-Throughput Phenotyping of Wheat Using Aerial Imaging. Plant Methods 2018, 14, 20. [Google Scholar] [CrossRef]
- Rawson, H.M.; Richards, R.A. Effects of High Temperature and Photoperiod on Floral Development in Wheat Isolines Differing in Vernalisation and Photoperiod Genes. Field Crop. Res. 1993, 32, 181–192. [Google Scholar] [CrossRef]
- Yang, J.; Sears, R.G.; Gill, B.S.; Paulsen, G.M. Quantitative and molecular characterization of heat tolerance in hexaploid wheat. Euphytica 2002, 126, 275–282. [Google Scholar] [CrossRef]
- Mason, R.E.; Mondal, S.; Beecher, F.W.; Pacheco, A.; Jampala, B.; Ibrahim, A.M.H.; Hays, D.B. QTL Associated with Heat Susceptibility Index in Wheat (Triticum aestivum L.) under Short-Term Reproductive Stage Heat Stress. Euphytica 2010, 174, 423–436. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, R.; Rane, J.; Gupta, V.K.; Mamrutha, H.M.; Tiwari, R. Mapping Quantitative Trait Loci Associated with Grain Filling Duration and Grain Number under Terminal Heat Stress in Bread Wheat (Triticum aestivum L.). Plant Breed. 2016, 135, 538–545. [Google Scholar] [CrossRef]
- Worland, A.J. The Influence of Flowering Time Genes on Environmental Adaptability in European Wheats. Euphytica 1996, 89, 49–57. [Google Scholar] [CrossRef]
- Deng, W.; Casao, M.C.; Wang, P.; Sato, K.; Hayes, P.M.; Finnegan, E.J.; Trevaskis, B. Direct Links between the Vernalization Response and Other Key Traits of Cereal Crops. Nat. Commun. 2015, 6, 5882. [Google Scholar] [CrossRef] [PubMed]
- Ochagavía, H.; Prieto, P.; Zikhali, M.; Griffiths, S.; Slafer, G.A. Earliness Per Se by Temperature Interaction on Wheat Development. Sci. Rep. 2019, 9, 2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.B.; Wong, D.; Tibbits, J.; Forrest, K.; Hayden, M.; Zhang, X.-Q.; Westcott, S.; Angessa, T.T.; Li, C. Targeted Enrichment by Solution-Based Hybrid Capture to Identify Genetic Sequence Variants in Barley. Sci. Data 2019, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.B.; Angessa, T.T.; McFawn, L.; Wong, D.; Tibbits, J.; Zhang, X.; Forrest, K.; Moody, D.; Telfer, P.; Westcott, S.; et al. Hybridisation-based Target Enrichment of Phenology Genes to Dissect the Genetic Basis of Yield and Adaptation in Barley. Plant Biotechnol. J. 2019, 17, 932–944. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.B.; Angessa, T.T.; Zhang, X.; Chen, K.; Zhou, G.; Tan, C.; Wang, P.; Westcott, S.; Li, C. A Global Barley Panel Revealing Genomic Signatures of Breeding in Modern Australian Cultivars. Plant J. 2021, 106, 419–434. [Google Scholar] [CrossRef]
- Laurie, D.A. Comparative Genetics of Flowering Time. Plant Mol. Biol. 1997, 35, 167–177. [Google Scholar] [CrossRef]
- Fernández-Calleja, M.; Casas, A.M.; Igartua, E. Major Flowering Time Genes of Barley: Allelic Diversity, Effects, and Comparison with Wheat. Theor. Appl. Genet. 2021, 134, 1867–1897. [Google Scholar] [CrossRef]
- Comadran, J.; Kilian, B.; Russell, J.; Ramsay, L.; Stein, N.; Ganal, M.; Shaw, P.; Bayer, M.; Thomas, W.; Marshall, D.; et al. Natural Variation in a Homolog of Antirrhinum Centroradialis Contributed to Spring Growth Habit and Environmental Adaptation in Cultivated Barley. Nat. Genet. 2012, 44, 1388–1392. [Google Scholar] [CrossRef]
- Casas, A.M.; Djemel, A.; Ciudad, F.J.; Yahiaoui, S.; Ponce, L.J.; Contreras-Moreira, B.; Gracia, M.P.; Lasa, J.M.; Igartua, E. HvFT1 (VrnH3) Drives Latitudinal Adaptation in Spanish Barleys. Theor. Appl. Genet. 2011, 122, 1293–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, B.; Deng, W.; Clausen, J.; Oliver, S.; Boden, S.; Hemming, M.; Trevaskis, B. Barley (Hordeum vulgare L.) Circadian Clock Genes Can Respond Rapidly to Temperature in an EARLY FLOWERING 3 -Dependent Manner. J. Exp. Bot. 2016, 67, 5517–5528. [Google Scholar] [CrossRef] [Green Version]
- Coast, O.; Šebela, D.; Quiñones, C.; Jagadish, S.V.K. Systematic Determination of the Reproductive Growth Stage Most Sensitive to High Night Temperature Stress in Rice (Oryza sativa). Crop Sci. 2020, 60, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Li, X.; Redoña, E.; Ishimaru, T.; Jagadish, K. Genetics and Breeding of Heat Tolerance in Rice. In Rice Improvement: Physiological, Molecular Breeding and Genetic Perspectives; Ali, J., Wani, S.H., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 203–220. ISBN 9783030665302. [Google Scholar]
- Zhao, Z.; Jiang, L.; Xiao, Y.; Zhang, W.; Zhai, H.; Wan, J. Identification of QTLs for heat tolerance at the booting stage in rice (Oryza sativa L.). Acta Agron. Sin. 2006, 32, 640–644. [Google Scholar]
- Ye, C.; Argayoso, M.A.; Redoña, E.D.; Sierra, S.N.; Laza, M.A.; Dilla, C.J.; Mo, Y.; Thomson, M.J.; Chin, J.; Delaviña, C.B.; et al. Mapping QTL for Heat Tolerance at Flowering Stage in Rice Using SNP Markers: Mapping QTL for Heat Tolerance at Flowering Stage in Rice. Plant Breed. 2012, 131, 33–41. [Google Scholar] [CrossRef]
- Ye, C.; Tenorio, F.A.; Argayoso, M.A.; Laza, M.A.; Koh, H.-J.; Redoña, E.D.; Jagadish, K.S.; Gregorio, G.B. Identifying and Confirming Quantitative Trait Loci Associated with Heat Tolerance at Flowering Stage in Different Rice Populations. BMC Genet. 2015, 16, 41. [Google Scholar] [CrossRef] [Green Version]
- Tazib, T.; Kobayashi, Y.; Koyama, H.; Matsui, T. QTL Analyses for Anther Length and Dehiscence at Flowering as Traits for the Tolerance of Extreme Temperatures in Rice (Oryza sativa L.). Euphytica 2015, 203, 629–642. [Google Scholar] [CrossRef]
- Xiao, Y.; Pan, Y.; Luo, L.; Zhang, G.; Deng, H.; Dai, L.; Liu, X.; Tang, W.; Chen, L.; Wang, G.-L. Quantitative Trait Loci Associated with Seed Set under High Temperature Stress at the Flowering Stage in Rice (Oryza sativa L.). Euphytica 2011, 178, 331–338. [Google Scholar] [CrossRef]
- Shanmugavadivel, P.S.; Sv, A.M.; Prakash, C.; Mk, R.; Tiwari, R.; Mohapatra, T.; Singh, N.K. High Resolution Mapping of QTLs for Heat Tolerance in Rice Using a 5K SNP Array. Rice 2017, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yuan, Z.; Vizcay-Barrena, G.; Yang, C.; Liang, W.; Zong, J.; Wilson, Z.A.; Zhang, D. PERSISTENT TAPETAL CELL1 Encodes a PHD-Finger Protein That Is Required for Tapetal Cell Death and Pollen Development in Rice. Plant Physiol. 2011, 156, 615–630. [Google Scholar] [CrossRef] [Green Version]
- Bheemanahalli, R.; Sathishraj, R.; Manoharan, M.; Sumanth, H.N.; Muthurajan, R.; Ishimaru, T.; Krishna, J.S.V. Is Early Morning Flowering an Effective Trait to Minimize Heat Stress Damage during Flowering in Rice? Field Crops Res. 2017, 203, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, H.; Sasaki, K.; Kambe, T.; Gannaban, R.B.; Miras, M.A.; Mendioro, M.S.; Simon, E.V.; Lumanglas, P.D.; Fujita, D.; Takemoto-Kuno, Y.; et al. QEMF3, a Novel QTL for the Early-Morning Flowering Trait from Wild Rice, Oryza Officinalis, to Mitigate Heat Stress Damage at Flowering in Rice, O. Sativa. J. Exp. Bot. 2015, 66, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, S.; Tabuchi, H.; Yatou, O.; Yoshida, H. Agronomic Traits and Gene Containment Capability of Cleistogamous Rice Lines with the Superwoman1-Cleistogamy Mutation. Breed. Sci. 2012, 62, 124–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, S.; Yamaguchi, T.; Ohmori, S.; Hayashi, T.; Yatou, O.; Yoshida, H. Cleistogamy Decreases the Effect of High Temperature Stress at Flowering in Rice. Plant Prod. Sci. 2015, 18, 111–117. [Google Scholar] [CrossRef]
- Zou, M.; Zhou, G.; Angessa, T.T.; Zhang, X.-Q.; Li, C. Polymorphism of Floral Type Gene Cly1 and Its Association with Thermal Stress in Barley. PLoS ONE 2018, 13, e0193390. [Google Scholar] [CrossRef]
- Tiwari, Y.K.; Yadav, S.K. High Temperature Stress Tolerance in Maize (Zea mays L.): Physiological and Molecular Mechanisms. J. Plant Biol. 2019, 62, 93–102. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature Extremes: Effect on Plant Growth and Development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Lizaso, J.I.; Ruiz-Ramos, M.; Rodríguez, L.; Gabaldon-Leal, C.; Oliveira, J.A.; Lorite, I.J.; Sánchez, D.; García, E.; Rodríguez, A. Impact of High Temperatures in Maize: Phenology and Yield Components. Field Crop. Res. 2018, 216, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering Dynamics, Pollen, and Pistil Contribution to Grain Yield in Response to High Temperature during Maize Flowering. Environ. Exp. Bot. 2019, 158, 80–88. [Google Scholar] [CrossRef]
- Liu, M.; Sheng, D.; Liu, X.; Wang, Y.; Hou, X.; Wang, Y.; Wang, P.; Guan, L.; Dong, X.; Huang, S. Dissecting heat tolerance and yield stability in maize from greenhouse and field experiments. J. Agron. Crop Sci. 2022, 208, 348–361. [Google Scholar] [CrossRef]
- Alam, M.A.; Seetharam, K.; Zaidi, P.H.; Dinesh, A.; Vinayan, M.T.; Nath, U.K. Dissecting Heat Stress Tolerance in Tropical Maize (Zea mays L.). Field Crop. Res. 2017, 204, 110–119. [Google Scholar] [CrossRef]
- Shah, F.; Huang, J.; Cui, K.; Nie, L.; Shah, T.; Chen, C.; Wang, K. Impact of High-Temperature Stress on Rice Plant and Its Traits Related to Tolerance. J. Agric. Sci. 2011, 149, 545–556. [Google Scholar] [CrossRef]
- Frey, F.P.; Urbany, C.; Hüttel, B.; Reinhardt, R.; Stich, B. Genome-Wide Expression Profiling and Phenotypic Evaluation of European Maize Inbreds at Seedling Stage in Response to Heat Stress. BMC Genom. 2015, 16, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemming, M.N.; Walford, S.A.; Fieg, S.; Dennis, E.S.; Trevaskis, B. Identification of High-Temperature-Responsive Genes in Cereals. Plant Physiol. 2012, 158, 1439–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravikiran, K.T.; Gopala Krishnan, S.; Vinod, K.K.; Dhawan, G.; Dwivedi, P.; Kumar, P.; Bansal, V.P.; Nagarajan, M.; Bhowmick, P.K.; Ellur, R.K.; et al. A Trait Specific QTL Survey Identifies NL44, a NERICA Cultivar as a Novel Source for Reproductive Stage Heat Stress Tolerance in Rice. Plant Physiol. Rep. 2020, 25, 664–676. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Pisipati, S.R.; Ristic, Z.; Bukovnik, U.; Fritz, A.K. Impact of Nighttime Temperature on Physiology and Growth of Spring Wheat. Crop Sci. 2008, 48, 2372–2380. [Google Scholar] [CrossRef]
- Yin, X.; Guo, W.; Spiertz, J.H. A Quantitative Approach to Characterize Sink–Source Relationships during Grain Filling in Contrasting Wheat Genotypes. Field Crop. Res. 2009, 114, 119–126. [Google Scholar] [CrossRef]
- Yang, H.; Gu, X.; Ding, M.; Lu, W.; Lu, D. Heat Stress during Grain Filling Affects Activities of Enzymes Involved in Grain Protein and Starch Synthesis in Waxy Maize. Sci. Rep. 2018, 8, 15665. [Google Scholar] [CrossRef] [Green Version]
- Spiertz, J.H.J.; Hamer, R.J.; Xu, H.; Primo-Martin, C.; Don, C.; van der Putten, P.E.L. Heat Stress in Wheat (Triticum aestivum L.): Effects on Grain Growth and Quality Traits. Eur. J. Agron. 2006, 25, 89–95. [Google Scholar] [CrossRef]
- Castro, M.; Peterson, C.J.; Rizza, M.D.; Dellavalle, P.D.; Vázquez, D.; IbáÑez, V.; Ross, A. Influence of Heat Stress on Wheat Grain Characteristics and Protein Molecular Weight Distribution. In Wheat Production in Stressed Environments; Buck, H.T., Nisi, J.E., Salomón, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 12, pp. 365–371. ISBN 9781402054969. [Google Scholar]
- Keeling, P.L.; Bacon, P.J.; Holt, D.C. Elevated Temperature Reduces Starch Deposition in Wheat Endosperm by Reducing the Activity of Soluble Starch Synthase. Planta 1993, 191, 342–348. [Google Scholar] [CrossRef]
- Shirdelmoghanloo, H.; Taylor, J.D.; Lohraseb, I.; Rabie, H.; Brien, C.; Timmins, A.; Martin, P.; Mather, D.E.; Emebiri, L.; Collins, N.C. A QTL on the Short Arm of Wheat (Triticum aestivum L.) Chromosome 3B Affects the Stability of Grain Weight in Plants Exposed to a Brief Heat Shock Early in Grain Filling. BMC Plant Biol. 2016, 16, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vetch, J.M.; Stougaard, R.N.; Martin, J.M.; Giroux, M.J. Review: Revealing the Genetic Mechanisms of Pre-Harvest Sprouting in Hexaploid Wheat (Triticum aestivum L.). Plant Sci. 2019, 281, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Porfirio, L.; Hughes, T.; Chen, J.; Dillon, S.; Gubler, F.; Ral, J.-P.F. Evaluation of the Impact of Heat on Wheat Dormancy, Late Maturity α-Amylase and Grain Size under Controlled Conditions in Diverse Germplasm. Sci. Rep. 2020, 10, 17800. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.-P.; Sadat, S.; Drenth, J.; McIntyre, C.L. The Heat Shock Factor Family from Triticum Aestivum in Response to Heat and Other Major Abiotic Stresses and Their Role in Regulation of Heat Shock Protein Genes. J. Exp. Bot. 2014, 65, 539–557. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.-J.; Chen, D.; Lynne Mclntyre, C.; Fernanda Dreccer, M.; Zhang, Z.-B.; Drenth, J.; Kalaipandian, S.; Chang, H.; Xue, G.-P. Heat Shock Factor C2a Serves as a Proactive Mechanism for Heat Protection in Developing Grains in Wheat via an ABA-Mediated Regulatory Pathway: TaHsfC2a Plays a Proactive Heat Protection Role. Plant Cell Environ. 2018, 41, 79–98. [Google Scholar] [CrossRef]
- Bhusal, N.; Sarial, A.K.; Sharma, P.; Sareen, S. Mapping QTLs for Grain Yield Components in Wheat under Heat Stress. PLoS ONE 2017, 12, e0189594. [Google Scholar] [CrossRef] [Green Version]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; McIntyre, C.L.; Olivares-Villegas, J.-J.; Chapman, S.C. Heat and Drought Adaptive QTL in a Wheat Population Designed to Minimize Confounding Agronomic Effects. Theor. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [Green Version]
- Paliwal, R.; Röder, M.S.; Kumar, U.; Srivastava, J.P.; Joshi, A.K. QTL Mapping of Terminal Heat Tolerance in Hexaploid Wheat (T. aestivum L.). Theor. Appl. Genet. 2012, 125, 561–575. [Google Scholar] [CrossRef]
- Pinto, R.S.; Lopes, M.S.; Collins, N.C.; Reynolds, M.P. Modelling and Genetic Dissection of Staygreen under Heat Stress. Theor. Appl. Genet. 2016, 129, 2055–2074. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, J.; Taylor, J.; Parent, B.; Bennett, D.; Reynolds, M.; Feuillet, C.; Langridge, P.; Mather, D. Multi-Environment Analysis and Improved Mapping of a Yield-Related QTL on Chromosome 3B of Wheat. Theor. Appl. Genet. 2013, 126, 747–761. [Google Scholar] [CrossRef]
- Parent, B.; Bonneau, J.; Maphosa, L.; Kovalchuk, A.; Langridge, P.; Fleury, D. Quantifying Wheat Sensitivities to Environmental Constraints to Dissect Genotype × Environment Interactions in the Field. Plant Physiol. 2017, 174, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Thomelin, P.; Bonneau, J.; Brien, C.; Suchecki, R.; Baumann, U.; Kalambettu, P.; Langridge, P.; Tricker, P.; Fleury, D. The Wheat Seven in Absentia Gene Is Associated with Increases in Biomass and Yield in Hot Climates. J. Exp. Bot. 2021, 72, 3774–3791. [Google Scholar] [CrossRef] [PubMed]
- Mangelsen, E.; Kilian, J.; Harter, K.; Jansson, C.; Wanke, D.; Sundberg, E. Transcriptome Analysis of High-Temperature Stress in Developing Barley Caryopses: Early Stress Responses and Effects on Storage Compound Biosynthesis. Mol. Plant 2011, 4, 97–115. [Google Scholar] [CrossRef]
- Mahalingam, R.; Bregitzer, P. Impact on Physiology and Malting Quality of Barley Exposed to Heat, Drought and Their Combination during Different Growth Stages under Controlled Environment. Physiol. Plantarum 2019, 165, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Sakata, T.; Takahashi, H.; Nishiyama, I.; Higashitani, A. Effects of High Temperature on the Development of Pollen Mother Cells and Microspores in Barley Hordeum vulgare L. J. Plant Res. 2000, 113, 395–402. [Google Scholar] [CrossRef]
- Shirdelmoghanloo, H.; Chen, K.; Paynter, B.; Angessa, T.T.; Westcott, S.; Khan, H.A.; Hill, C.B.; Li, C. Grain-filling rate improves physical grain quality in barley under heat stress conditions during the grain-filling period. Front. Plant Sci. 2022, 13, 858652. [Google Scholar] [CrossRef]
- Zhang, D.-P.; Zhou, Y.; Yin, J.-F.; Yan, X.-J.; Lin, S.; Xu, W.-F.; Baluška, F.; Wang, Y.-P.; Xia, Y.-J.; Liang, G.; et al. Rice G-Protein Subunits QPE9-1 and RGB1 Play Distinct Roles in Abscisic Acid Responses and Drought Adaptation. J. Exp. Bot. 2015, 66, 6371–6384. [Google Scholar] [CrossRef] [Green Version]
- Watt, C.; Zhou, G.; McFawn, L.-A.; Chalmers, K.J.; Li, C. Fine Mapping of QGL5H, a Major Grain Length Locus in Barley (Hordeum vulgare L.). Theor. Appl. Genet. 2019, 132, 883–893. [Google Scholar] [CrossRef]
- Watt, C.; Zhou, G.; Angessa, T.T.; Moody, D.; Li, C. A Novel Polymorphism in the 5′ UTR of HvDEP1 Is Associated with Grain Length and 1000-Grain Weight in Barley (Hordeum vulgare L.). Crop Pasture Sci. 2020, 71, 752. [Google Scholar] [CrossRef]
- Miura, S.; Crofts, N.; Saito, Y.; Hosaka, Y.; Oitome, N.F.; Watanabe, T.; Kumamaru, T.; Fujita, N. Starch Synthase IIa-Deficient Mutant Rice Line Produces Endosperm Starch With Lower Gelatinization Temperature Than Japonica Rice Cultivars. Front. Plant Sci. 2018, 9, 645. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Guo, T.; Wan, X.; Wang, H.; Zhu, M.; Li, A.; Su, N.; Shen, Y.; Mao, B.; Zhai, H.; et al. Transcriptome Analysis of Grain-Filling Caryopses Reveals Involvement of Multiple Regulatory Pathways in Chalky Grain Formation in Rice. BMC Genom. 2010, 11, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambardekar, A.A.; Siebenmorgen, T.J.; Counce, P.A.; Lanning, S.B.; Mauromoustakos, A. Impact of Field-Scale Nighttime Air Temperatures during Kernel Development on Rice Milling Quality. Field Crop. Res. 2011, 122, 179–185. [Google Scholar] [CrossRef]
- Gann, P.J.; Esguerra, M.; Counce, P.A.; Srivastava, V. Genotype-dependent and Heat-induced Grain Chalkiness in Rice Correlates with the Expression Patterns of Starch Biosynthesis Genes. Plant-Environ. Interact. 2021, 2, 165–176. [Google Scholar] [CrossRef]
- Tabassum, R.; Dosaka, T.; Ichida, H.; Morita, R.; Ding, Y.; Abe, T.; Katsube-Tanaka, T. FLOURY ENDOSPERM11-2 Encodes Plastid HSP70-2 Involved with the Temperature-dependent Chalkiness of Rice (Oryza sativa L.). Grains. Plant J. 2020, 103, 604–616. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive Expression Profiling of Rice Grain Filling-Related Genes under High Temperature Using DNA Microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef] [Green Version]
- Crafts-Brandner, S.J.; Salvucci, M.E. Sensitivity of Photosynthesis in a C4 Plant, Maize, to Heat Stress. Plant Physiol. 2002, 129, 1773–1780. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, E.P.; Mullen, R.E.; Keeling, P.L.; Singletary, G.W. Heat Stress during Grain Filling in Maize: Effects on Kernel Growth and Metabolism. Crop Sci. 1999, 39, 1733–1741. [Google Scholar] [CrossRef]
- Yang, H.; Lu, D.; Shen, X.; Cai, X.; Lu, W. Heat Stress at Different Grain Filling Stages Affects Fresh Waxy Maize Grain Yield and Quality. Cereal Chem. J. 2015, 92, 258–264. [Google Scholar] [CrossRef]
- Lu, D.; Lu, W. Effects of Heat Stress During Grain Filling on the Functional Properties of Flour from Fresh Waxy Maize. Cereal Chem. J. 2013, 90, 65–69. [Google Scholar] [CrossRef]
- Yuan, Y.; Cairns, J.E.; Babu, R.; Gowda, M.; Makumbi, D.; Magorokosho, C.; Zhang, A.; Liu, Y.; Wang, N.; Hao, Z.; et al. Genome-Wide Association Mapping and Genomic Prediction Analyses Reveal the Genetic Architecture of Grain Yield and Flowering Time Under Drought and Heat Stress Conditions in Maize. Front. Plant Sci. 2019, 9, 1919. [Google Scholar] [CrossRef] [Green Version]
- Longmei, N.; Gill, G.K.; Zaidi, P.H.; Kumar, R.; Nair, S.K.; Hindu, V.; Vinayan, M.T.; Vikal, Y. Genome Wide Association Mapping for Heat Tolerance in Sub-Tropical Maize. BMC Genom. 2021, 22, 154. [Google Scholar] [CrossRef] [PubMed]
- Seetharam, K.; Kuchanur, P.H.; Koirala, K.B.; Tripathi, M.P.; Patil, A.; Sudarsanam, V.; Das, R.R.; Chaurasia, R.; Pandey, K.; Vemuri, H.; et al. Genomic Regions Associated with Heat Stress Tolerance in Tropical Maize (Zea mays L.). Sci. Rep. 2021, 11, 13730. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.; Reynolds, M.; Mullan, D.; Izanloo, A.; Kuchel, H.; Langridge, P.; Schnurbusch, T. Detection of Two Major Grain Yield QTL in Bread Wheat (Triticum aestivum L.) under Heat, Drought and High Yield Potential Environments. Theor. Appl. Genet. 2012, 125, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, T.; Yu, T.; Zhang, S.; Mao, X.; Zhao, J.; Wang, X.; Dong, J.; Liu, B. Integrating Small RNA Sequencing with QTL Mapping for Identification of MiRNAs and Their Target Genes Associated with Heat Tolerance at the Flowering Stage in Rice. Front. Plant Sci. 2017, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Withanawasam, D.M.; Kommana, M.; Pulindala, S.; Eragam, A.; Moode, V.N.; Kolimigundla, A.; Puram, R.V.; Palagiri, S.; Balam, R.; Vemireddy, L.R. Improvement of Grain Yield under Moisture and Heat Stress Conditions through Marker-Assisted Pedigree Breeding in Rice. Crop Pasture Sci. 2022, 73, 356–369. [Google Scholar] [CrossRef]
- Sade, N.; del Mar Rubio-Wilhelmi, M.; Umnajkitikorn, K.; Blumwald, E. Stress-Induced Senescence and Plant Tolerance to Abiotic Stress. J. Exp. Bot. 2018, 69, 845–853. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Chai, M.; Guo, Z.; Shi, X.; Li, Y.; Tang, J.; Zhang, Z. Dissecting the Regulatory Network of Leaf Premature Senescence in Maize (Zea mays L.) Using Transcriptome Analysis of ZmELS5 Mutant. Genes 2019, 10, 944. [Google Scholar] [CrossRef] [Green Version]
- Breeze, E.; Harrison, E.; McHattie, S.; Hughes, L.; Hickman, R.; Hill, C.; Kiddle, S.; Kim, Y.; Penfold, C.A.; Jenkins, D.; et al. High-Resolution Temporal Profiling of Transcripts during Arabidopsis Leaf Senescence Reveals a Distinct Chronology of Processes and Regulation. Plant Cell 2011, 23, 873–894. [Google Scholar] [CrossRef] [Green Version]
- Zentgraf, U.; Doll, J.; Riester, L. Live and Let Die: The Core Circadian Oscillator Coordinates Plant Life History and Pilots Leaf Senescence. Mol. Plant 2018, 11, 351–353. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Y.; Zou, D.; Zhao, Y.; Wang, H.-L.; Zhang, Y.; Xia, X.; Luo, J.; Guo, H.; Zhang, Z. LSD 3.0: A Comprehensive Resource for the Leaf Senescence Research Community. Nucleic Acids Res. 2020, 48, D1069–D1075. [Google Scholar] [CrossRef]
- Farooq, M.; Bramley, H.; Palta, J.A.; Siddique, K.H.M. Heat Stress in Wheat during Reproductive and Grain-Filling Phases. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Park, S.-Y.; Kim, Y.-S.; Wang, S.-H.; Yoo, S.-C.; Hörtensteiner, S.; Paek, N.-C. Arabidopsis STAY-GREEN2 Is a Negative Regulator of Chlorophyll Degradation during Leaf Senescence. Mol. Plant 2014, 7, 1288–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, M.; Pudake, R.N.; Singh, V.P.; Joshi, A.K. Association of Staygreen Trait with Canopy Temperature Depression and Yield Traits under Terminal Heat Stress in Wheat (Triticum aestivum L.). Euphytica 2013, 190, 87–97. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Stay-Green in Spring Wheat Can Be Determined by Spectral Reflectance Measurements (Normalized Difference Vegetation Index) Independently from Phenology. J. Exp. Bot. 2012, 63, 3789–3798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talukder, S.K.; Babar, M.A.; Vijayalakshmi, K.; Poland, J.; Prasad, P.V.V.; Bowden, R.; Fritz, A. Mapping QTL for the Traits Associated with Heat Tolerance in Wheat (Triticum aestivum L.). BMC Genet. 2014, 15, 97. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Harikrishna; Barman, D.; Ghimire, O.P.; Gurumurthy, S.; Singh, P.K.; Chinnusamy, V.; Padaria, J.C.; Arora, A. Stay-Green Trait Serves as Yield Stability Attribute under Combined Heat and Drought Stress in Wheat (Triticum aestivum L.). Plant Growth Regul. 2022, 96, 67–78. [Google Scholar] [CrossRef]
- Hays, D.B.; Do, J.H.; Mason, R.E.; Morgan, G.; Finlayson, S.A. Heat Stress Induced Ethylene Production in Developing Wheat Grains Induces Kernel Abortion and Increased Maturation in a Susceptible Cultivar. Plant Sci. 2007, 172, 1113–1123. [Google Scholar] [CrossRef]
- Gous, P.W.; Hickey, L.; Christopher, J.T.; Franckowiak, J.; Fox, G.P. Discovery of QTL for Stay-Green and Heat-Stress in Barley (Hordeum vulgare L.) Grown under Simulated Abiotic Stress Conditions. Euphytica 2016, 207, 305–317. [Google Scholar] [CrossRef]
- Kilasi, N.L.; Singh, J.; Vallejos, C.E.; Ye, C.; Jagadish, S.V.K.; Kusolwa, P.; Rathinasabapathi, B. Heat Stress Tolerance in Rice (Oryza sativa L.): Identification of Quantitative Trait Loci and Candidate Genes for Seedling Growth Under Heat Stress. Front. Plant Sci. 2018, 9, 1578. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-M.; Chao, D.-Y.; Wu, Y.; Huang, X.; Chen, K.; Cui, L.-G.; Su, L.; Ye, W.-W.; Chen, H.; Chen, H.-C.; et al. Natural Alleles of a Proteasome A2 Subunit Gene Contribute to Thermotolerance and Adaptation of African Rice. Nat. Genet. 2015, 47, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Owttrim, G.W. RNA Helicases and Abiotic Stress. Nucleic Acids Res. 2006, 34, 3220–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Qin, B.; Li, X.; Tang, D.; Zhang, Y.; Cheng, Z.; Xue, Y. Nucleolar DEAD-Box RNA Helicase TOGR1 Regulates Thermotolerant Growth as a Pre-RRNA Chaperone in Rice. PLoS Genet. 2016, 12, e1005844. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, X.; Shi, Y.; Xu, X.; Li, L.; Wu, J.-L. Premature Senescence Leaf 50 Promotes Heat Stress Tolerance in Rice (Oryza sativa L.). Rice 2021, 14, 63, Erratum in Rice 2021, 14, 53. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Huang, T.; Ding, M.; Lu, D.; Lu, W. High Temperature during Grain Filling Impacts on Leaf Senescence in Waxy Maize. Agron. J. 2017, 109, 906–916. [Google Scholar] [CrossRef]
- Diaz-Mendoza, M.; Velasco-Arroyo, B.; Santamaria, M.E.; González-Melendi, P.; Martinez, M.; Diaz, I. Plant Senescence and Proteolysis: Two Processes with One Destiny. Genet. Mol. Biol. 2016, 39, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Prochazkova, D.; Sairam, R.K.; Srivastava, G.C.; Singh, D.V. Oxidative Stress and Antioxidant Activity as the Basis of Senescence in Maize Leaves. Plant Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Li, H.; Zhang, H.; Li, G.; Liu, Z.; Zhang, Y.; Zhang, H.; Guo, X. Expression of Maize Heat Shock Transcription Factor Gene ZmHsf06 Enhances the Thermotolerance and Drought-Stress Tolerance of Transgenic Arabidopsis. Funct. Plant Biol. 2015, 42, 1080. [Google Scholar] [CrossRef]
- Li, H.; Li, G.; Guo, X. Cloning, expression characteristics and subcellular location of heat shock transcription factor gene (ZmHsf06) in Zea mays. J. Agric. Biotech. 2015, 23, 41–51. [Google Scholar]
- Jiang, Y.; Zheng, Q.; Chen, L.; Liang, Y.; Wu, J. Ectopic Overexpression of Maize Heat Shock Transcription Factor Gene ZmHsf04 Confers Increased Thermo and Salt-Stress Tolerance in Transgenic Arabidopsis. Acta Physiol. Plant. 2018, 40, 9. [Google Scholar] [CrossRef]
- Shirdelmoghanloo, H.; Cozzolino, D.; Lohraseb, I.; Collins, N.C. Truncation of Grain Filling in Wheat (Triticum aestivum L.) Triggered by Brief Heat Stress during Early Grain Filling: Association with Senescence Responses and Reductions in Stem Reserves. Funct. Plant Biol. 2016, 43, 919. [Google Scholar] [CrossRef]
- Gaudin, A.C.M.; Henry, A.; Sparks, A.H.; Slamet-Loedin, I.H. Taking Transgenic Rice Drought Screening to the Field. J. Exp. Bot. 2013, 64, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.B.; Taylor, J.D.; Edwards, J.; Mather, D.; Bacic, A.; Langridge, P.; Roessner, U. Whole-Genome Mapping of Agronomic and Metabolic Traits to Identify Novel Quantitative Trait Loci in Bread Wheat Grown in a Water-Limited Environment. Plant Physiol. 2013, 162, 1266–1281. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.B.; Taylor, J.D.; Edwards, J.; Mather, D.; Langridge, P.; Bacic, A.; Roessner, U. Detection of QTL for Metabolic and Agronomic Traits in Wheat with Adjustments for Variation at Genetic Loci That Affect Plant Phenology. Plant Sci. 2015, 233, 143–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Sukumaran, S.; Claverie, E.; Sansaloni, C.; Dreisigacker, S.; Reynolds, M. Genetic Dissection of Heat and Drought Stress QTLs in Phenology-Controlled Synthetic-Derived Recombinant Inbred Lines in Spring Wheat. Mol. Breed. 2019, 39, 34. [Google Scholar] [CrossRef]
- Schmidt, J.; Tricker, P.J.; Eckermann, P.; Kalambettu, P.; Garcia, M.; Fleury, D. Novel Alleles for Combined Drought and Heat Stress Tolerance in Wheat. Front. Plant Sci. 2020, 10, 1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, P.; Lu, L.; Jia, L.; Kabir, M.R.; Zhang, J.; Lan, T.; Zhao, Y.; Xin, M.; Hu, Z.; Yao, Y.; et al. Global QTL Analysis Identifies Genomic Regions on Chromosomes 4A and 4B Harboring Stable Loci for Yield-Related Traits Across Different Environments in Wheat (Triticum aestivum L.). Front. Plant Sci. 2018, 9, 529. [Google Scholar] [CrossRef] [Green Version]
- Maulana, F.; Ayalew, H.; Anderson, J.D.; Kumssa, T.T.; Huang, W.; Ma, X.-F. Genome-Wide Association Mapping of Seedling Heat Tolerance in Winter Wheat. Front. Plant Sci. 2018, 9, 1272. [Google Scholar] [CrossRef]
- Templer, S.E.; Ammon, A.; Pscheidt, D.; Ciobotea, O.; Schuy, C.; McCollum, C.; Sonnewald, U.; Hanemann, A.; Förster, J.; Ordon, F.; et al. Metabolite Profiling of Barley Flag Leaves under Drought and Combined Heat and Drought Stress Reveals Metabolic QTLs for Metabolites Associated with Antioxidant Defense. J. Exp. Bot. 2017, 68, 1697–1713. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Yuan, Q.; Lin, H.; Li, X.; Zhang, C.; Gao, H.; Zhang, B.; He, H.; Liu, T.; Jie, Z.; et al. Linkage Analysis, GWAS, Transcriptome Analysis to Identify Candidate Genes for Rice Seedlings in Response to High Temperature Stress. BMC Plant Biol. 2021, 21, 85. [Google Scholar] [CrossRef]
- Wu, N.; Yao, Y.; Xiang, D.; Du, H.; Geng, Z.; Yang, W.; Li, X.; Xie, T.; Dong, F.; Xiong, L. A MITE Variation-associated Heat-inducible Isoform of a Heat-shock Factor Confers Heat Tolerance through Regulation of Jasmonate Zim-Domain Genes in Rice. New Phytol. 2022, 234, 1315–1331. [Google Scholar] [CrossRef] [PubMed]
- GM Approval Database. Available online: https://www.isaaa.org/gmapprovaldatabase/ (accessed on 23 March 2022).
- Tian, B.; Talukder, S.K.; Fu, J.; Fritz, A.K.; Trick, H.N. Expression of a Rice Soluble Starch Synthase Gene in Transgenic Wheat Improves the Grain Yield under Heat Stress Conditions. In Vitro Cell. Dev. Biol.-Plant 2018, 54, 216–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casaretto, J.A.; El-kereamy, A.; Zeng, B.; Stiegelmeyer, S.M.; Chen, X.; Bi, Y.-M.; Rothstein, S.J. Expression of OsMYB55 in Maize Activates Stress-Responsive Genes and Enhances Heat and Drought Tolerance. BMC Genom. 2016, 17, 312. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Kaur, A.; Pandey, A.; Mamrutha, H.M.; Singh, G.P. CRISPR-Based Genome Editing in Wheat: A Comprehensive Review and Future Prospects. Mol. Biol. Rep. 2019, 46, 3557–3569. [Google Scholar] [CrossRef] [PubMed]
- Nandy, S.; Pathak, B.; Zhao, S.; Srivastava, V. Heat-shock-inducible CRISPR/Cas9 System Generates Heritable Mutations in Rice. Plant Direct 2019, 3, e00145. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Kang, S.; He, L.; Zhao, J.; Zhang, S.; Hu, J.; Zeng, D.; Zhang, G.; Dong, G.; Gao, Z.; et al. The Newly Identified Heat-Stress Sensitive Albino 1 Gene Affects Chloroplast Development in Rice. Plant Sci. 2018, 267, 168–179. [Google Scholar] [CrossRef]
- Suh, S.; Johnson, J.A.; Tambjerg, L.; Sim, S.; Broeckx-Smith, S.; Reyes, W.; Chaplin-Kramer, R. Closing Yield Gap Is Crucial to Avoid Potential Surge in Global Carbon Emissions. Glob. Environ. Chang. 2020, 63, 102100. [Google Scholar] [CrossRef]
- Tian, X.; Engel, B.A.; Qian, H.; Hua, E.; Sun, S.; Wang, Y. Will Reaching the Maximum Achievable Yield Potential Meet Future Global Food Demand? J. Clean. Prod. 2021, 294, 126285. [Google Scholar] [CrossRef]
- Bergkamp, B.; Impa, S.M.; Asebedo, A.R.; Fritz, A.K.; Jagadish, S.V.K. Prominent Winter Wheat Varieties Response to Post-Flowering Heat Stress under Controlled Chambers and Field Based Heat Tents. Field Crop. Res. 2018, 222, 143–152. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef]
- Rutkoski, J.; Poland, J.; Mondal, S.; Autrique, E.; Pérez, L.G.; Crossa, J.; Reynolds, M.; Singh, R. Canopy Temperature and Vegetation Indices from High-Throughput Phenotyping Improve Accuracy of Pedigree and Genomic Selection for Grain Yield in Wheat. G3 Genes Genomes Genet. 2016, 6, 2799–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, K.; Park, C. Thermal Adaptation and Plasticity of the Plant Circadian Clock. New Phytol. 2019, 221, 1215–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mody, T.; Bonnot, T.; Nagel, D.H. Interaction between the Circadian Clock and Regulators of Heat Stress Responses in Plants. Genes 2020, 11, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnot, T.; Gillard, M.B.; Nagel, D.H. CAST-R: An Application to Visualize Circadian and Heat Stress-Responsive Genes in Plants. Plant Physiol. 2022, kiac121. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C. Argentina First to Market with Drought-Resistant GM Wheat. Nat. Biotechnol. 2021, 39, 652. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Function | Reference |
|---|---|---|---|
| DEHYDRATION-RESPONSIVE ELEMENT BINDING PROTEIN | DREB2A | Transcriptional regulators involved in plant responses to cold, drought, and salt stress | [30] |
| HEAT SHOCK PROTEIN | HSP | Molecular chaperones protecting the proteome against environmental stresses; thermomemory | [25] |
| HEAT SHOCK TRANSCRIPTION FACTOR | HSF | Part of signal transduction chains mediating the activation of genes responsive to both heat stress and other stresses | [23] |
| ASCORBATE PEROXIDASE | APX | Antioxidant system-related enzyme | [32] |
| CATALASE | CAT | Antioxidant system-related enzyme, long-term heat tolerance | [33] |
| Gene Name | Gene Symbol | Function | Reference |
|---|---|---|---|
| Filamentous temperature sensitive H | FtsH | Metalloprotease, regulation of heat-shock transcription factor σ32; thermomemory | [37] |
| HEAT SHOCK PROTEIN 21 | HSP21 | Molecular chaperones protecting the proteome against environmental stresses; thermomemory | [25,37] |
| ANTI-SILENCING FUNCTION 1 | ASF1 | Histone chaperone | [21] |
| FORGETTER3/ HEAT SHOCK TRANSCRIPTION FACTOR A3 | FGT3/HSFA3 | FGT3 encodes the HSFA3 gene; mediates heat stress memory by direct transcriptional activation of memory-related genes | [34] |
| Trait | Impact | QTL, Locus or Marker | Candidate Gene | Chromosome | Crop Species | Reference |
|---|---|---|---|---|---|---|
| Grain filling period duration | − | QHgfd.iiwbr-1B, QHgfd.iiwbr-5A | n.a. | 1B, 2B, 3B, 5A, 6B | bread wheat | [53] |
| Grain filling period duration | − | QHgfd.tam-1D (gwm337), QHgfd.tam-2A (wmc407), QHgfd.tam-6D (gwm325), QGfd.tam-1Bc (barc137), QGfd.tam-2A (wmc407), QGfd.tam-2D (cfd43) | n.a. | 1B, 1D, 2A, 2D 6D, | bread wheat | [52] |
| Grain filling period duration | − | Xgwm11, Xgwm293 | n.a. | 1B, 5A | bread wheat | [51] |
| Temperature sensitivity (late reproductive phase) | + | Eps | ELF3 | 1D | bread wheat | [56] |
| Early inflorescence development | + | n.a. | Ppd-1 | 2H | bread wheat | [50] |
| Life cycle duration/temperature sensitivity | −/+ | Mat.a8, eam8 | ELF3 | 1HL | barley | [64] |
| Reproductive phase duration | + | n.a. | ODDSOC2 | 3H | barley | [87] |
| Flower opening time | − | qEMF3 | n.a. | 3 | rice | [75] |
| Cleistogamy | + | spw1-cls | SPW1 | 6 | rice | [76,77] |
| Spikelet fertility at flowering | − | qHTSF1.1, qHTSF4.1 | n.a. | 1, 4 | rice | [68] |
| Spikelet fertility at flowering | − | 109 QTLs | n.a. | 1–12 | rice | [88] |
| Pollen fertility | − | qPF4 (RM5687—RM471), qPF6 (RM190—RM225) | n.a. | 4, 6 | rice | [71] |
| Trait | Impact | QTL, Locus or Marker | Candidate Gene | Chromosome | Crop Species | Reference |
|---|---|---|---|---|---|---|
| Grain number | − | QHknm.tam-1A (cfa2129), QHknm.tam-2B (barc200.2), QHknm.tam-3Bc (barc147), QHknm.tam-4A (wmc89), QHknm.tam-5B (gwm213) | n.a. | 1A, 2B, 3B, 4A, 5B | bread wheat | [52] |
| Grain weight | − | QHkwm.tam-1B (gwm268), QHkwm.tam-2B (gwm111.2), QHkwm.tam-3B (wmc527), QHkwm.tam-5A (gwm291), QHkwm.tam-6D (gwm325) | n.a. | 1B, 2B, 3B, 5A, 6D | bread wheat | [52] |
| Grain weight | − | QTL11, QTL27 | n.a. | 3B, 6B | bread wheat | [95,127] |
| Grain yield | − | Q.Yld.aww-3B-2, Q.Yld.aww-3B-1, Q.Yld.aww-3D, Q.Yld.aww-4D, Q.Yld.aww-5B, Q.Yld.aww-7A-1, Q.Yld.aww-7A-2 | n.a. | 3A, 3D, 4D 5B, 7A | bread wheat | [127] |
| Grain yield, grain number, grain weight | − | qYDH.3BL | SINA | 3B | bread wheat | [104,106] |
| Grain size, thousand-kernel weight | − | qGL5H | DEP1 | 5H | barley | [112,113] |
| Chalkiness | + | flo11-2 | cpHSP70-2 | 12 | rice | [118,128] |
| Grain yield | − | qSSPF10, qHT6 (RM190-RM587-RM510-RM225-RM217-RM50) | n.a. | 6 | rice | [128,129] |
| Grain yield | − | 1549 SNPs | 673 candidate genes including U-box domain-containing protein, GRAS family transcription factor, and protein kinase family protein | 1, 2, 4, 5, 8, 10 | maize | [124] |
| Grain yield | − | 269 SNPs | 140 candidate gene models, including bHLH (basic-Helix-Loop-Helix) transcription factors | 1–10 | maize | [126] |
| Grain yield | − | 12 SNPs | 9 candidate gene models | 1, 3, 6, 7, 10 | maize | [125] |
| Trait | Impact | QTL, Locus or Marker | Candidate Gene | Chromosome | Crop Species | Reference |
|---|---|---|---|---|---|---|
| Chlorophyll content | − | Q.Spad.aww-4D, Q.Spad.aww-6D | n.a. | 4D, 6D | bread wheat | [127] |
| Various senescence traits (incl. chlorophyll content) | − | QTL2, QTL13, QTL15, QTL18, QTL21, QTL29 | n.a. | 1A, 4A, 4B, 5A, 7B | bread wheat | [95,155] |
| Staygreen | − | 44 QTLs | n.a. | 2A, 4B, 4D, 6A, 7D | bread wheat | [103] |
| Staygreen | − | 6 QTLs, including HSPFLQ1 (bPb-5529) | lbi1 | 3H, 4H, 5H, 6H | barley | [143] |
| Seedling survival | − | TT1 | Os03g0387100 (PAB1) | 3 | rice | [145] |
| Biomass | − | togr1-1 | Os03g46610 (TOGR1) | 3 | rice | [147] |
| Leaf senescence | + | psl50 | Os01g50770 (PSL50) | 1 | rice | [148] |
| Seed germination rate, chlorophyll content | − | n.a. | ZmHsf06 | 1 | maize | [152] |
| Seedling survival | − | n.a. | ZmHsf04 | 1 | maize | [154] |
| Leaf senescence | + | ZmELS5 | n.a. | n.a. | maize | [132] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, C.B.; Li, C. Genetic Improvement of Heat Stress Tolerance in Cereal Crops. Agronomy 2022, 12, 1205. https://doi.org/10.3390/agronomy12051205
Hill CB, Li C. Genetic Improvement of Heat Stress Tolerance in Cereal Crops. Agronomy. 2022; 12(5):1205. https://doi.org/10.3390/agronomy12051205
Chicago/Turabian StyleHill, Camilla Beate, and Chengdao Li. 2022. "Genetic Improvement of Heat Stress Tolerance in Cereal Crops" Agronomy 12, no. 5: 1205. https://doi.org/10.3390/agronomy12051205