1. Introduction
Tamarillo (
Solanum betaceum Cav.), commonly called tree tomato or “tomate de la paz”, in Spanish [
1], is native to the Andean region of South America [
2,
3], where it grows wild or in orchards. It is cultivated for its juicy edible fruits, and has interesting nutritional properties due to low caloric levels [
4] and relatively high protein content, vitamins, minerals [
5], and chemicals with important antioxidant, therapeutic, and preventive properties [
6,
7,
8].
Tamarillo can be propagated by seeds or conventional methods of vegetative propagation, such as cutting and grafting [
4]. However, propagation through seeds does not assure the genetic uniformity of plants, besides being a slower process thus compromising productivity. Traditional cloning methods applied to this species have proven to be difficult given that the propagated plants are sometimes infected with diseases [
4]. Production of pathogen-free material is the first step of controlling diseases [
9]. In Vitro micropropagation techniques, as an alternative method of cloning, have been applied to many crops to propagate selected genotypes or hybrids and to scale-up cloning [
10].
Although in vitro cloning is nowadays a set of techniques of common use, the transfer of plants to the field is usually a crucial step of the micropropagation process that quite often limits the success of these techniques [
11,
12,
13]. A critical step in micropropagation, is the adaptation of in vitro produced plantlets to ex vitro environments, due to the shock resulting from the transfer from a high humidity environment to much drier conditions [
14,
15]. In general, plants need several weeks to adapt to the new ex vitro conditions and correct anomalies in their morphology, anatomy, and physiology, induced by the special conditions of in vitro culture [
15,
16,
17]. In other words, they need an acclimatization period.
Acclimatization refers to the climatic adaptation of an organism that has been placed in a new environment that is different from the previous one [
18]. Thus, an effective protocol for acclimatization of in vitro propagated plants must be developed to assure that losses during this process are residual.
The functionality of the adventitious root system is essential to plant survival during the acclimatization process, as it affects the absorption of water and nutrients, which were previously guaranteed by the in vitro medium [
17]. Therefore, factors such as substrate and mineral nutrition may also affect the success of the acclimatization process.
According to Moreira et al. [
19], choosing a suitable substrate reduces plant mortality during acclimatization. Several authors have witnessed different results in the performance of the cultures according to the type of substrate used. For example, the use of high amounts of vermiculite [
20], a mixture of peat and vermiculite [
21], or a combination of soil and vermiculite [
22] have resulted in better crop performance in terms of plant growth [
20,
22], root length, as well as root growth and penetration into the subsoil [
20]. Moreover, some substrates facilitate the functional link between soil/root microbiota and subsequent increase in crop yield [
21]. In the same way, according to Betancourt-Osorio et al. [
23] plant nutritional status may significantly influence plant acclimatization, and according to Oliet et al. [
24], the way nutrients are made available to plants can substantially influence seedling development.
The most commonly used commercial fertilizers are water soluble quick-release fertilizers (QRF) that are predictively readily available for plants [
25]. QRF, also called conventional fertilizers, are ideal for pre-plant applications and are the most used in nurseries in the fertigation form [
25]. However, the application of solid fertilizers to the substrate, particularly fertilizers that are not immediately available to plants, have proven to be a good alternative [
24,
26], because conventional fertilizers, due to their immediate release, do not follow the dynamic needs of crop growth [
25,
27].
An alternative are the controlled-release fertilizers (CRF) or slow-release fertilizers (SRF).The release rate of CRFs is designed in a way that allows for synchronized nutrient release that matches the needs of plants at different stages of development. In these kind of fertilizers, granules are coated or encapsulated by organic or inorganic materials that control the rate, pattern, and duration of the nutrient release [
25,
28]. On the contrary, in SRFs, nutrient release is not controlled because it depends on microbial activity, which, in turns, depends on environmental conditions such as temperature and humidity [
25,
29,
30,
31].
There are several protocols for tamarillo cloning using proliferation of axillary shoots [
32], organogenesis [
33,
34], and somatic embryogenesis [
35,
36]. However, a detailed study concerning the effects of substrate and mineral nutrition on the acclimatization of the micropropagated plants has not been performed yet. The objective of this work was the comparative analysis of four substrates and two types of mineral nutrition (QRF and CRF) as well as their absence, and their influence on the growth and development of tamarillo micropropagated plantlets during the acclimatization process. The evaluation of these factors was carried out through the study of photosynthetic parameters such as the chlorophyll content index and stomatal conductance, as well as through plant growth parameters such as the quantification of the biomass of the acclimatized plants.
3. Results
Ex vitro simultaneous acclimatization and rooting of tamarillo plants occurred with survival rates of 75.0, 81.3, and 93.8% for plants acclimatized with 1.6, 0.8, and 0.4 and 0 g/alveoli of CRF (second test), respectively. In the first test, there was a survival rate of 100% in all treatments.
It was not possible to perform measurements of g
s and the CCI in the plantlets that were not nutritionally supplemented (in both tests) because they were too small to correctly position the measuring equipment on leaves (
Figure 1). The results clearly show (
Figure 1) that the mineral nutrition has a considerable impact on plant growth during ex vitro acclimatization, making it possible to shorten the duration of this period and have plants ready to proceed to the greenhouse/field faster.
There were no significant differences at the level of g
s, regarding the type of substrate (
Table 1). However, in relation to the type of fertilizer, it was found that g
s was significantly (
p = 0.02) higher in plants supplemented with CRF comparatively to QRF (
Table 1). Similar results were obtained for the CCI, which was also significantly (
p = 0.00) higher in plants treated with CRF (
Table 1).
The CCI was also significantly higher in plants whose substrate consisted of a mixture with vermiculite when compared to those without vermiculite (
Table 1).
Table 2 shows how dry biomass is affected by fertilization. The results indicate that the amount of biomass was significantly higher in plants supplemented with mineral nutrients. Such as for CCI, the dry biomass of roots and shoots was significantly higher in plants whose substrate consisted of a mixture with vermiculite when compared to those without vermiculite, as shown in
Table 3.
The interaction between substrates (P, PP, PV, and PPV) and fertilizer (C, QRF, and CRF) did not have a significant effect on g
s, CCI, shoots’ dry biomass, and total dry biomass. Therefore, the factors act independently on these variables (
Table 4), but the interaction substrate/fertilizer had a significant effect for the roots dry biomass (
Table 4). Thus, root growth is promoted by a substrate containing a mixture with vermiculite (
Table 3) and supplemented with CRF (
Table 2).
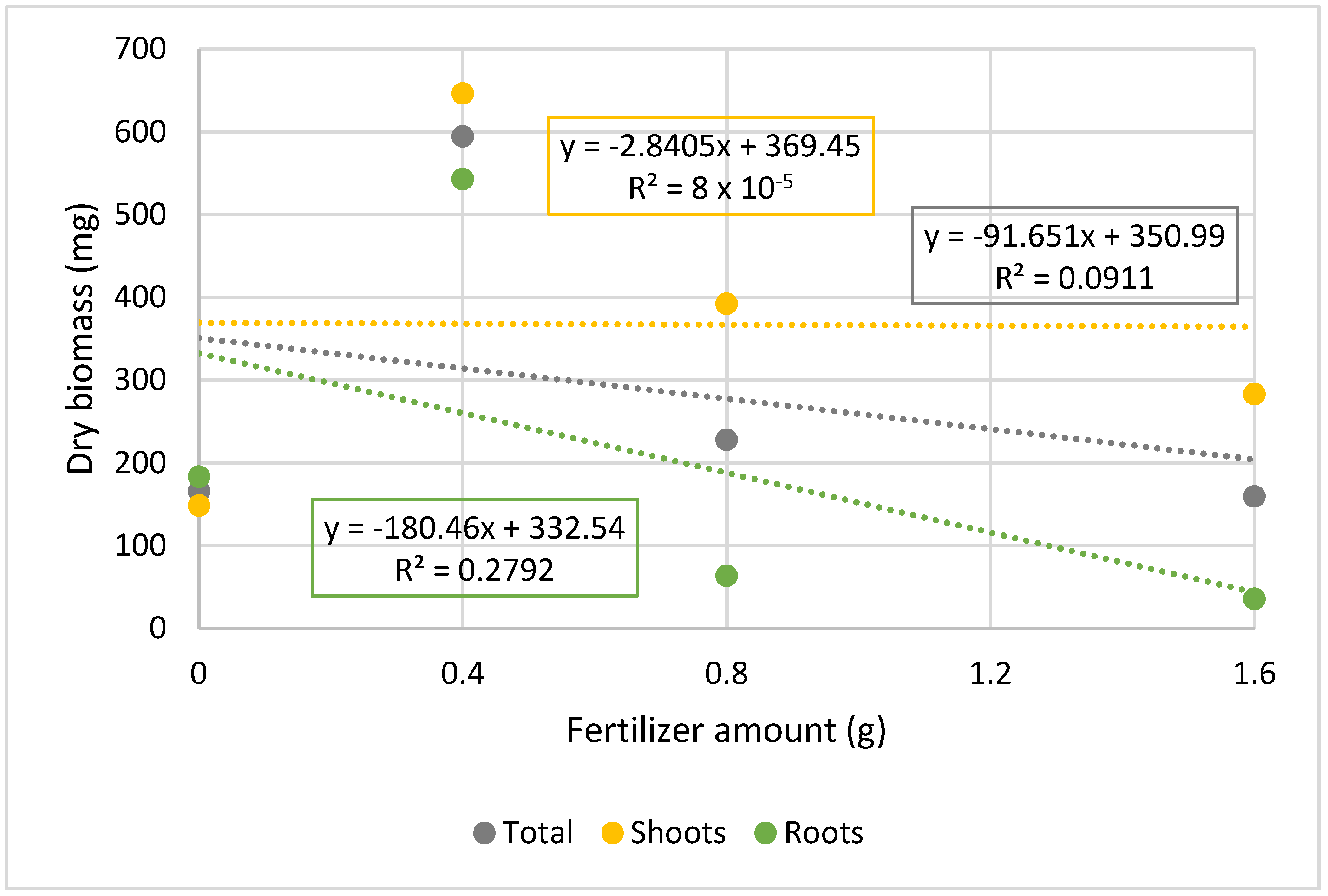
The second assay was established in a mixture of peat and vermiculite, to which different amounts of CRF were added. The results displayed in
Table 5, show that the total dry biomass (entire plantlets) is significantly higher in plants acclimatized in alveoli containing 0.4 g CRF. This means that the CRF has a positive impact on the growth and development of the tamarillo plantlets during the acclimatization process (low values of total dry biomass in not treated plantlets), but only when a certain amount of fertilizer is supplied. In fact, 0.8 and 1.6 g CRF/alveoli considerably reduce all the parameters analyzed. These results were corroborated by the evaluation of the physiological parameters (stomatal conductance and chlorophyll content) presented in
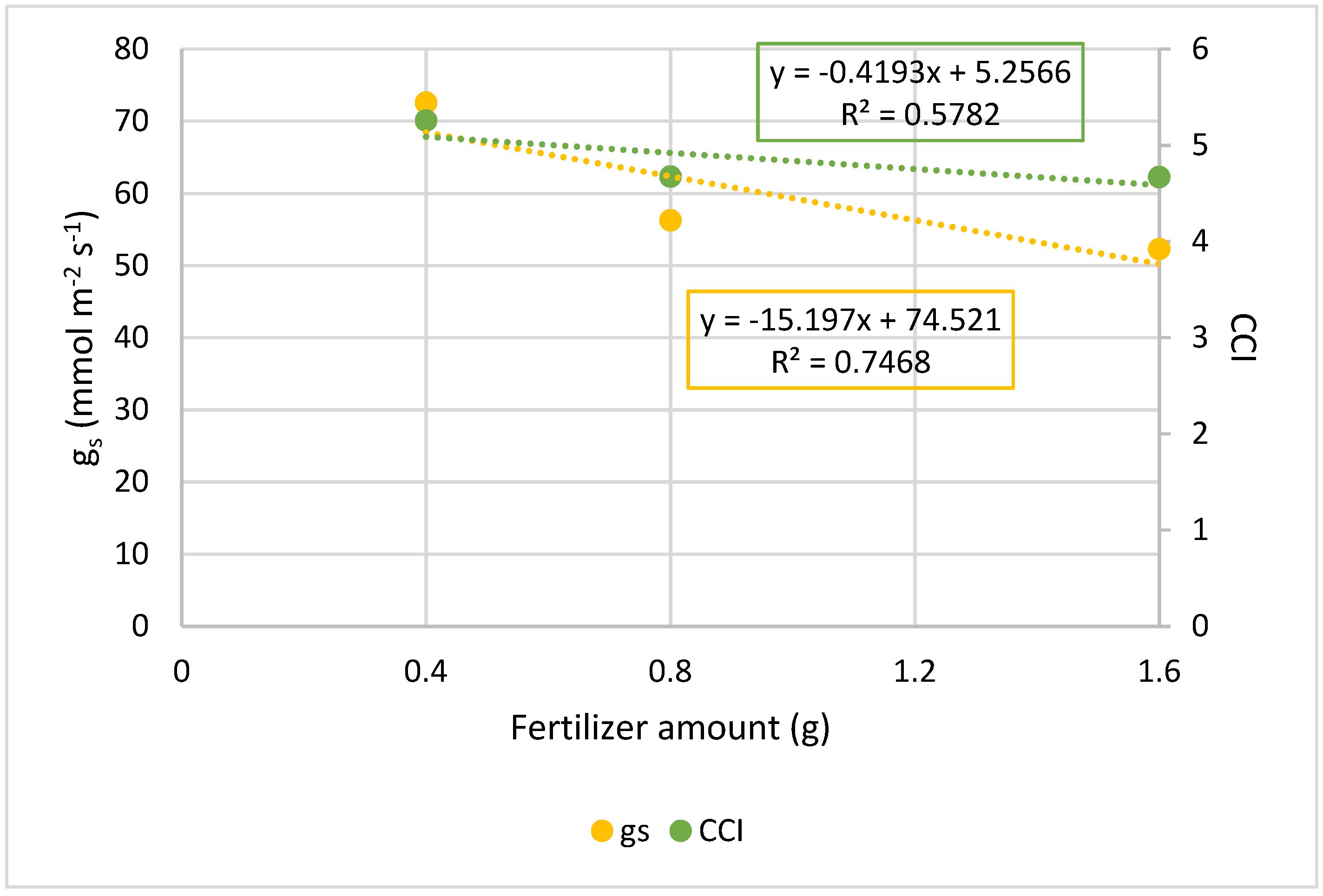
Table 6, and in
Figure 2 and
Figure 3, in which the regression lines slopes are negative for total biomass as well as for stomatal conductance and chlorophyll content.
4. Discussion
The results obtained revealed survival rates above 75%, which demonstrates the high capacity of this species to adapt to ex vitro conditions [
38,
39]. However, the survival rate results obtained in the first (100%) and second (between 75 and 93.8%) experiments are somewhat different. Even so, from the observation of
Figure 1, and
Table 1 and
Table 2, it is evident that CRF has a positive influence on tamarillo plant development during the acclimatization/rooting period, although there may be some variability in the results [
38]. As no bibliographic references concerning this aspect were found, the second test was carried out in an attempt to find a reference dose of CRF for tamarillo during the acclimatization/rooting phase.
In addition to the survival rate, the g
s and the chlorophyll content were evaluated since, according to Mielke and Schaffer [
40], the ability of a given species to acclimatize in different environments can be evidenced by the evaluation of the initial growth and the plasticity of the photosynthetic system.
A critical aspect for plant survival during the acclimatization process is the development of an effective stomatic regulation of transpiration [
15,
16,
41,
42], because the main problem after the transfer from the in vitro to the ex vitro environment, it is the high rate of water losses by the plants. The reason why it was decided to test the acclimatization of
S. betaceum on different substrates, which has no direct influence on the plants’ physiology, was because this could condition water availability until they achieved effective control of stomatal activity. The substrates were made with mixtures of perlite, a very porous material, and vermiculite, which has a high water-holding capacity. Thus, it was assured that tamarillo plantlets could overcome its known waterlogging problems [
23] and the moisture necessary for plants adjusting to the ex vitro environment, leading to it being considerably dryer than in vitro conditions. However, no significant differences in the values of g
s, in relation to the type of substrate were found. Perhaps this is because the mechanisms that regulate stomatal activity are similar at this stage of plant development, since stomatal activity tends to be less variable in very young or older plants [
43]. However, when the effect analyzed was the type of fertilizer, it was found that the g
s was significantly higher in plants supplemented with CRF comparatively to QRF. In the same way, the CCI was also significantly higher in plants that received CRF.
Between 50 to 70% of the nitrogen (N) in the leaves is associated with enzymes present in chloroplasts, pointing to a direct relationship between the N and chlorophyll contents [
44,
45,
46]. However, our results showed that the plants treated with the fertilizer that provides 10% nitrogen (CRF) had a significantly higher CCI when compared to the fertilizer that provides 12% nitrogen (QRF). Apparently, these results are not due to the N content of the fertilizers (which is still similar), but to their release characteristics.
According to Oliet et al. [
24] the form of nutrient delivery can substantially influence the development of the plants. Thus, the application of gradual release fertilizers has been a promising alternative to conventional water soluble quick-release fertilizers, commonly used in nurseries and for pre-plant applications [
24,
25], because as CRF release nutrients over time they replenish nutrients in the soil solution as nutrients are released from the fertilizer granules [
47].
Considering that chlorophyll content is crucial for photosynthetic efficiency and, consequently, to growth and adaptability to different environments, the use of gradual release fertilizers can contribute to obtain plants of better potential quality, such as those observed by Oliet et al. [
24], Almeida et al. [
30], Rossa et al. [
31], and Neto et al. [
48].
The CCI was also significantly higher in plants whose substrate consisted of a mixture with vermiculite, compared to those without vermiculite, but according to Afonso et al. [
49], it is possible the photosynthetic pigments show both qualitative and quantitative changes according to the substrate composition, as found by these researchers in
Albizia niopoides. Thus, in our experiments, it seems plausible to assume that, due to the greater water retention capacity of vermiculite [
20], which in this case would be a nitrogen solution (QRF and CRF), and given the relationship between N and chloroplasts, chlorophyll was superior in plants developing in substrates with this element.
According to Costa [
50], the nutritional status of plants can be determined by analyzing their composition, and the best way to do this is, in a first approach, is to evaluate dry matter. From
Table 2 it is evident that the amount of biomass was significantly higher in plants supplemented with mineral nutrients. These results are in agreement with observations [
51] showing that plants from nutrient-rich sites tend to produce more biomass per unit nutrient in the plant. Similar results were observed by other researchers. Thus, Ramírez-Soler et al. [
52], reported that unfertilized tamarillo plants showed a decrease in the dry matter of roots, shoots, and total, in relation to well-nourished plants. Clark and Richardson [
53] recorded that there is a relationship between the accumulation of biomass in the tamarillo plants and the mineral nutrients, and Betancourt-Osorio et al. [
23] found that well-nourished tamarillo plants showed a significantly higher shoot length than poorly nourished plants.
Our data showed that the dry biomass of roots and shoots was higher in plants acclimatized in substrates made up of mixtures with vermiculite (PV; PPV). Similar results were obtained by Jacobs et al. [
20] in
Pseudotsuga menziesii, wherein peat and vermiculite facilitated the penetration of roots into the substrate, resulting in greater root volume and length, and greater average height. Thus, considering the first test performed, the substrates were made up of mixtures with vermiculite and supplemented with mineral nutrients through the application of CRF, which seems to favor the ex vitro acclimatization of tamarillo plants, being evident that it is mineral nutrition that has the greatest impact in this process.
In this framework, the second test was established in a mixture of peat and vermiculite and applying different concentrations of CRF, with the objective of determining which are the best amounts that should be used in tamarillo. The results showed that CRF has a positive impact on tamarillo growth during the acclimation process, given the low values of total dry biomass of non-treated plants, which reinforces the results obtained in the first test. However, this positive effect was only observed when relative low amounts (0.4 g/alveoli) were used. According to Larcher [
44], having satisfied the needs of the plants, an increase in nutrients can confer advantages in terms of competition for increasing resistance to pathogens or adverse climatic conditions. However, this can also not promote any improvement, just luxury consumption [
51]. Additionally, when the concentrations are too high, they can even become toxic [
51]. In our experiments, it seems that luxury consumptions may have occurred for the two highest concentrations of fertilizer (0.8 and 1.6 per alveoli). Because of that, these plants might not be stimulated to expand the root system in search of nutrients. Supporting this assumption are the data in
Table 5, showing the difference between the dry biomass of the roots and shoots of the non-fertilized plants when compared with those treated with 0.4 g/alveoli of CRF; the last ones display more proportional values, also comparatively with the plants treated with 0.8 and 1.6 g/alveoli of CRF. The development and architecture of the root system can interfere with the greater or lesser success of plant establishment in the field, since the root biomass is an indicative of the better performance of the plants when transferred to the field, because they present greater capacity of support and absorption of water and nutrients, increasing the productive potential and the ability to adapt to adverse environmental conditions [
54,
55].
The regression analyses of these results show that the root system is the most sensitive to variations in concentration of CRF. It can also be seen (
Figure 2) that the amount of dry biomass (total, roots, and shoots) is greater for a smaller concentration of CRF (0.4 g/alveoli), and that all slopes of the regression lines are negative, which suggests that with this concentration of CRF, toxicity phenomena may also have occurred. Similar results were obtained by Silva et al. [
56] who found that CRF had a significant influence on the growth of
Acacia mangium; however, there was a reduction in seedling gain after a specific dose, which varied according to the parameters evaluated.
The results showed in
Figure 3 also indicate that, similar to dry biomass, the slopes of the regression lines established between the amount of CRF and the physiological parameters of g
s and CCI are negative, and therefore, the greater the amount of CRF, the lower the g
s and the chlorophyll content of the plants. Moreover, g
s and CCI were significantly higher in plants treated with 0.4 g/alveoli of CRF, compared to those treated with the highest amounts, thus supporting the idea that the 0.8 and 1.6 g/alveoli fertilizer’s concentrations would be too high for these plants in these conditions, becoming toxic.
Taken together, the results indicate that in tamarillo, after the ex vitro transplantation, supplementation with nutrients promotes plant development and reduces the acclimatization period, allowing plants to be ready to go to the greenhouse/field more quickly, and giving them competitive advantages due to their more developed root systems and stem growth, and therefore, greater photosynthetic capacity. It was also found that the amount of CRF used in the first test would have been too high, given the results of the second test (under the same conditions), but as the comparison term was the application of fertilizer or its absence, and as it was found that supplementation with mineral nutrients has a positive impact on the acclimatization process; that specific dose of CRF was always better than nothing. However, this test revealed that these plants are very small (seedlings) and fragile, their nutritional needs are low, and toxicity phenomena can easily arise at the same time that resources are being consumed unnecessarily.
Several researchers have focused their research on the crucial issue of acclimatization of micropropagated plants, and there are several works describing methodologies that allow improving the performance of these plants in vitro, giving them greater resistance to later ex vitro conditions, such as through manipulation of the culture medium composition [
57], the use of bioreactors [
58], or selenium nanoparticles [
59]. Perez-Jiménez et al. [
60] went further and studied the effect of different CO
2 concentrations on the ex vitro acclimatization process in
Cynara scolymus plants. Our study is a further contribution, advancing the use of mineral nutrients that were usually only used by nurseries, when the plants are already perfectly adapted to the ex vitro environment, and showed potential for the future development of a protocol for ex vitro acclimatization, in the sense to make the process faster and losses during this phase minimal.
Thus, CRFs are interesting alternatives, as they can reduce the time to obtain suitable plantlets for the market and shorten the production cycle, reducing labor and mainly offering effective measures to be implemented to improve the quality of plantlets and optimize production costs [
26], when testing/knowing the specific needs of each plant species at each stage of its state of growth and development.







{kind=link}
{kind=link}
{kind=link}