
Species-Enriched Grass-Clover Mixtures Can Promote Bumblebee Abundance Compared with Intensively Managed Conventional Pastures


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Region and Design
2.2. Pollinator Monitoring
2.3. Statistical Analysis
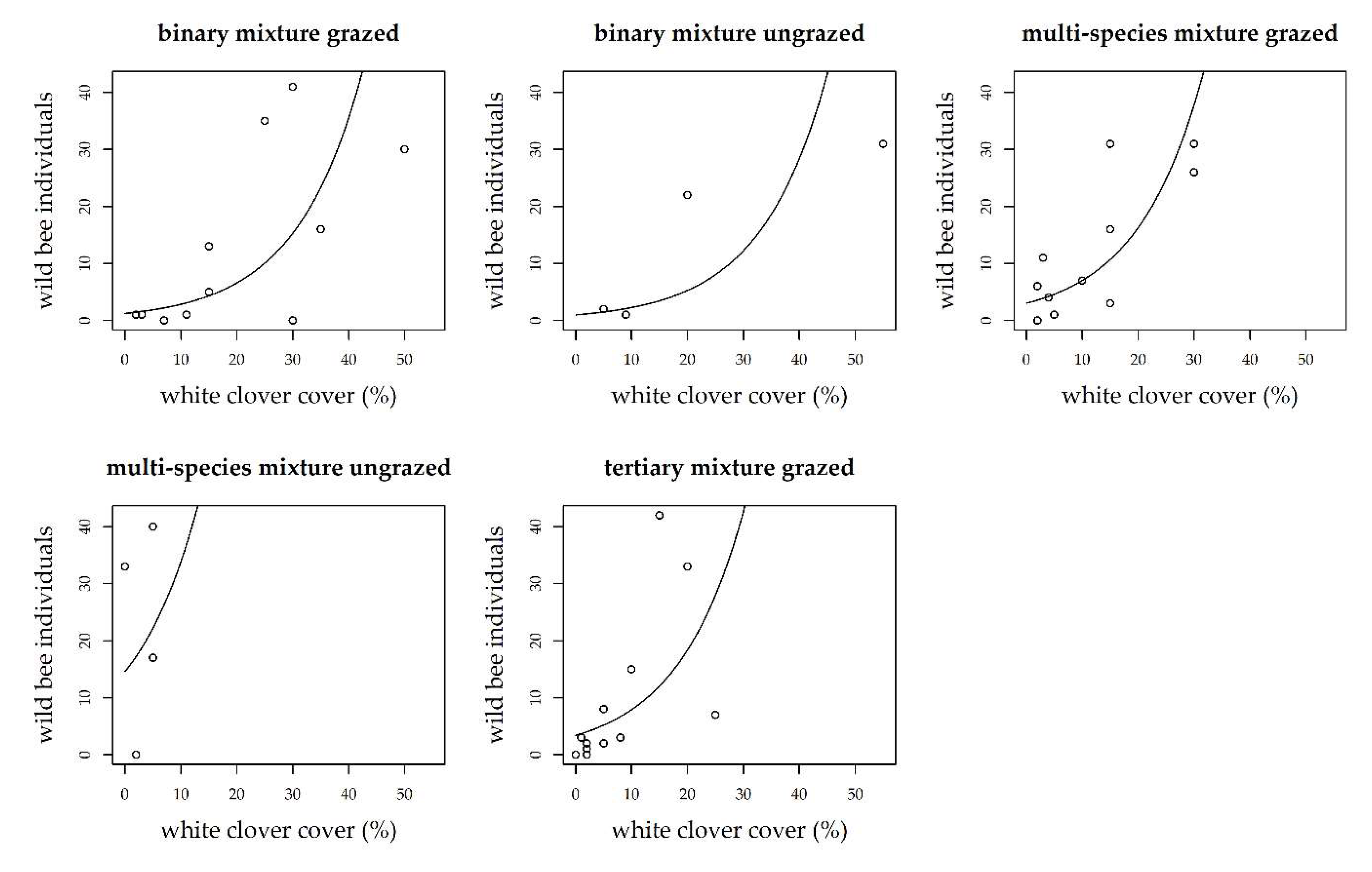
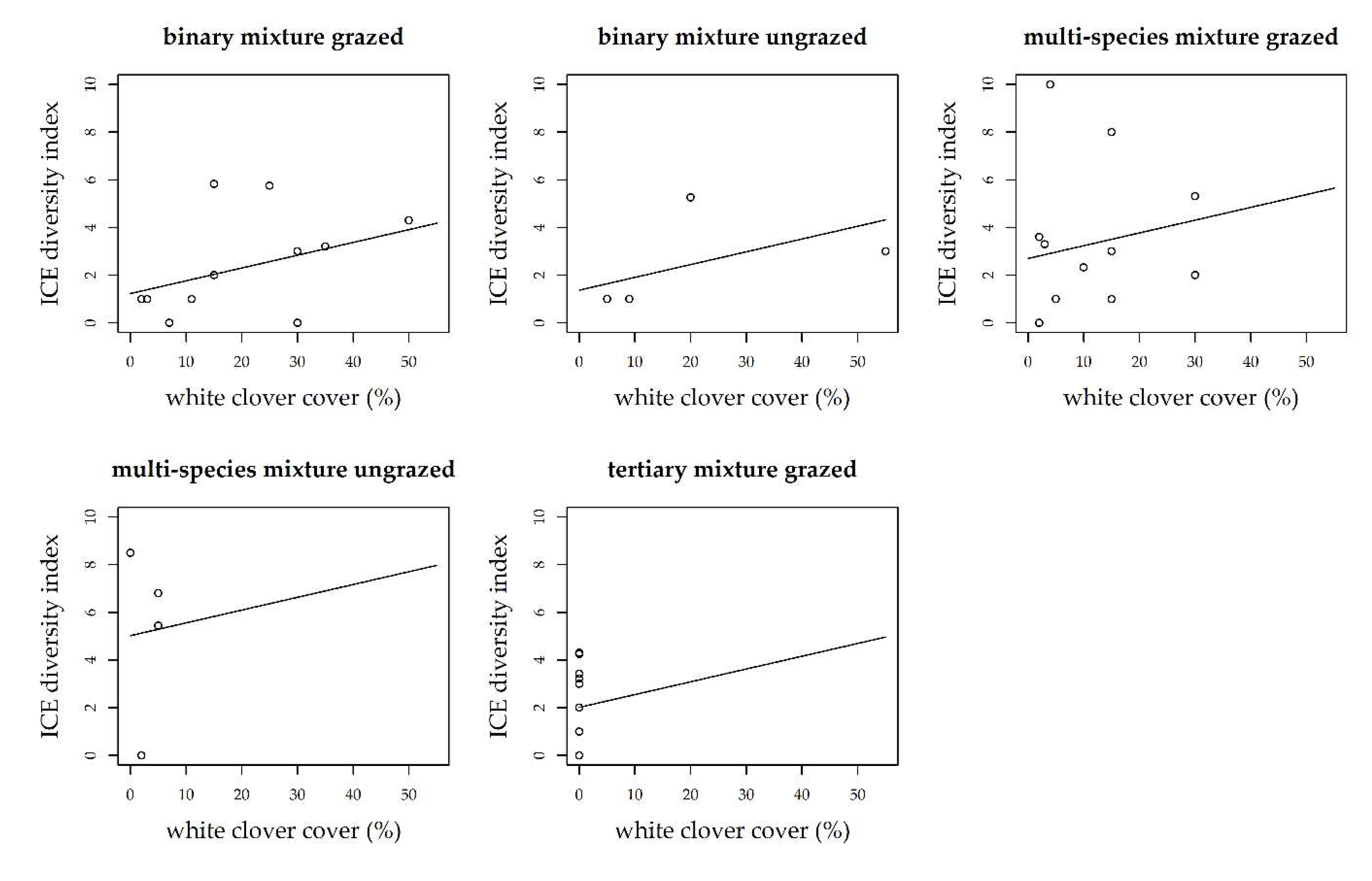
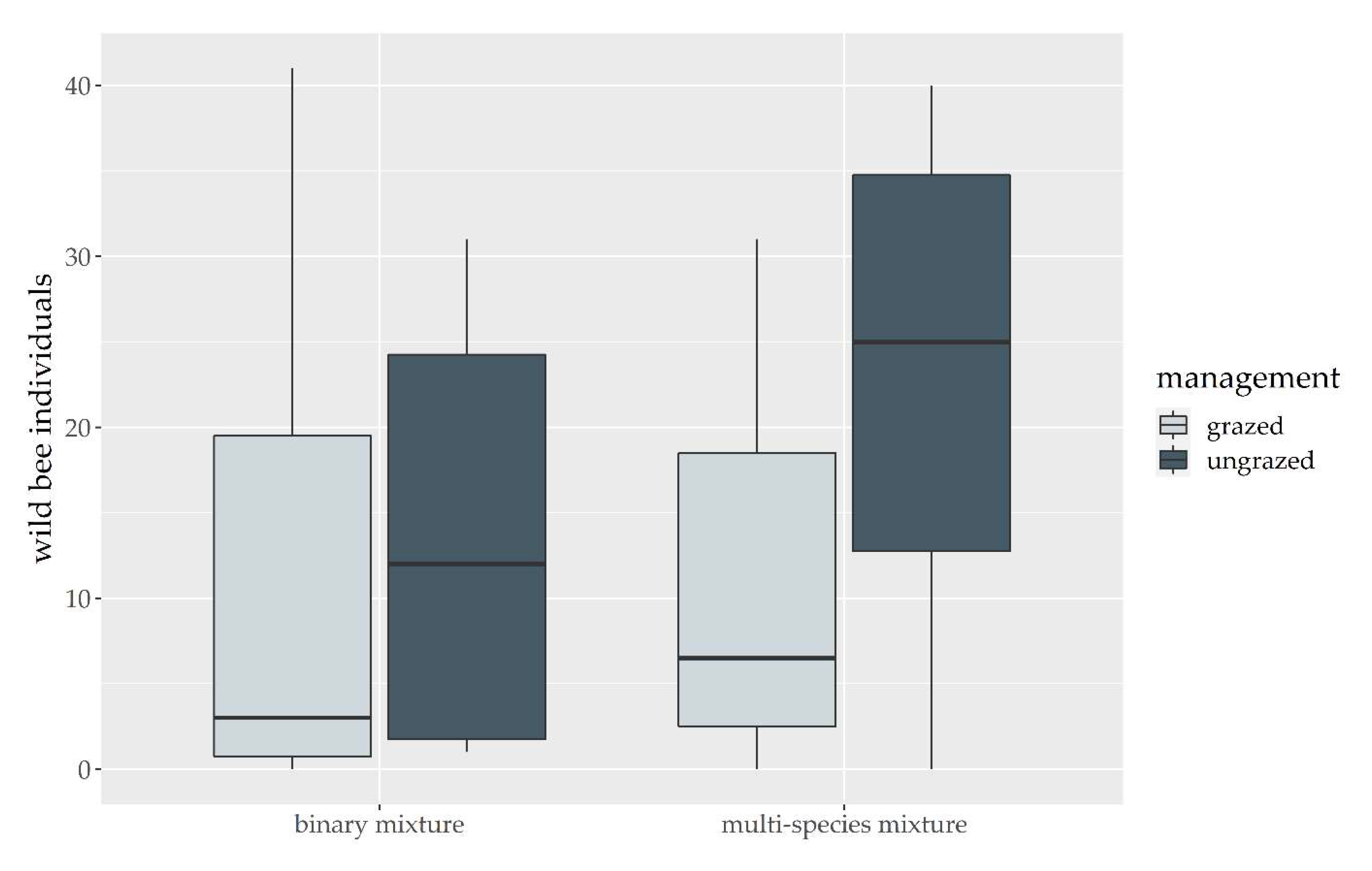
3. Results
3.1. Grass-Clover Mixtures
3.2. Management
3.3. Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Species | Variety | Binary Mixture | Tertiary Mixture | Multispecies Mixture |
|---|---|---|---|---|
| Perennial ryegrass (Lolium perenne) | Discuss | 6 kg | 6 kg | 4 kg |
| Calvano1 | 6 kg | 6 kg | 4 kg | |
| Astonenergy | 6 kg | 6 kg | 4 kg | |
| Astonhockey | 6 kg | 6 kg | 4 kg | |
| Red clover (Trifolium repens) | Vysocan | 2 kg | 1 kg | 0.65 kg |
| Liflex | 2 kg | 1 kg | 0.65 kg | |
| White clover (Trifolium pratense) | Harmonie | 3 kg | 1.5 kg | |
| Larus | 3 kg | 1.5 kg | ||
| Plantain (Plantago lanceolata) | 4n’Herculese | 1 kg | ||
| Chicory (Cichorium intybus) | Spadona | 2 kg | ||
| Sheep’s burnet (Sanguisorba minor) | Burnet | 2 kg | ||
| Caraway (Carum carvi) | Volhouden | 2 kg | ||
| Birdsfoot trefoil (Lotus corniculatus) | Lotanova | 3 kg |
| Estimate | SE | z-Value | p-Value | |
|---|---|---|---|---|
| binary mix.—tertiary mixture | 1.0248 | 0.5895 | 1.738 | 0.191 |
| binary mix.—multispecies mix. | 0.9044 | 0.5662 | 1.597 | 0.247 |
| tertiary mix.—multispecies mix. | −0.1205 | 0.5602 | −0.215 | 0.975 |
| Estimate | SE | z-Value | p-Value | |
|---|---|---|---|---|
| tertiary mix.—binary mix. | 0.7870 | 1.8747 | 0.420 | 0.907 |
| multispecies mix.—binary mix. | 1.4718 | 1.8068 | 0.815 | 0.694 |
| multispecies mix.—tert. mix. | 0.6848 | 1.8199 | 0.376 | 0.925 |
| Estimate | SE | z-Value | p-Value | |
|---|---|---|---|---|
| binary mix. ungr.—binary mix. grazed | 0.143 | 2.021 | 0.071 | 1.000 |
| multispecies mix ungr.—multispecies mix. grazed | 2.324 | 2.032 | 1.144 | 0.523 |
| multispecies mix. ungr.—binary mix. ungr. | 3.652 | 2.290 | 1.595 | 0.262 |
| Binary Mixture, Grazed | Tertiary Mixture, Grazed | Multispecies Mixture, Grazed | Binary Mixture, Ungrazed | Multispecies Mixture, Ungrazed | Conve-ntional | |
|---|---|---|---|---|---|---|
| Perennial ryegrass | 45.85 | 41.85 | 35,05 | 59.7 | 33.3 | 100 |
| White clover | 53.55 | 27.85 | 27.65 | 39.1 | 13.8 | |
| Red clover | 29.75 | 19.2 | 37.8 | |||
| Birdsfoot trefoil | 2.95 | 11.1 | ||||
| Other sown species | 14,95 | 2.9 | ||||
| Unsown species | 0.6 | 0.55 | 0.35 | 1.2 | 1.1 | |
| Total per year | 11.2 | 13.1 | 12.1 | 7.3 | 10.7 | 12.5 |
References
- Van Vliet, J.; de Groot, H.L.; Rietveld, P.; Verburg, P.H. Manifestations and Underlying Drivers of Agricultural Land Use Change in Europe. Landsc. Urban Plan. 2015, 133, 24–36. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renard, D.; Tilman, D. National Food Production Stabilized by Crop Diversity. Nature 2019, 571, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Meehan, T.D.; Werling, B.P.; Landis, D.A.; Gratton, C. Agricultural Landscape Simplification and Insecticide Use in the Midwestern United States. Proc. Natl. Acad. Sci. USA 2011, 108, 11500–11505. [Google Scholar] [CrossRef] [Green Version]
- Sims, B.; Kienzle, J. Making Mechanization Accessible to Smallholder Farmers in Sub-Saharan Africa. Environments 2016, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Jaleta, M.; Baudron, F.; Krivokapic-Skoko, B.; Erenstein, O. Agricultural Mechanization and Reduced Tillage. Int. J. Agric. Sustain. 2019, 17, 219–230. [Google Scholar] [CrossRef]
- Meeus, J.H.A. The Transformation of Agricultural Landscapes in Western Europe. Sci. Total Environ. 1993, 129, 171–190. [Google Scholar] [CrossRef]
- Hooftman, D.A.P.; Bullock, J.M. Mapping to Inform Conservation. Biol. Conserv. 2012, 145, 30–38. [Google Scholar] [CrossRef]
- White, E.V.; Roy, D.P. A Contemporary Decennial Examination of Changing Agricultural Field Sizes Using Landsat Time Series Data. Geogr. Environ. 2015, 2, 33–54. [Google Scholar] [CrossRef]
- Aguilar, J.; Gramig, G.G.; Hendrickson, J.R.; Archer, D.W.; Forcella, F.; Liebig, M.A.; Hart, J.P. Crop Species Diversity Changes in the United States. PLoS ONE 2015, 10, e0136580. [Google Scholar] [CrossRef]
- Feranec, J.; Jaffrain, G.; Soukup, T.; Hazeu, G. Determining Changes and Flows in European Landscapes 1990–2000 Using CORINE Land Cover Data. Appl. Geogr. 2010, 30, 19–35. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland Biodiversity. Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Allan, E.; Bossdorf, O.; Dormann, C.F.; Prati, D.; Gossner, M.M.; Tscharntke, T.; Blüthgen, N.; Bellach, M.; Birkhofer, K.; Boch, S.; et al. Interannual Variation in Land-Use Intensity Enhances Grassland Multidiversity. Proc. Natl. Acad. Sci. USA 2014, 111, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Kremen, C.; Merenlender, A.M. Landscapes That Work for Biodiversity and People. Science 2018, 362, eaau6020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruijven, J.; et al. High Plant Diversity Is Needed to Maintain Ecosystem Services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Tittonell, P. Ecological Intensification of Agriculture—Sustainable by Nature. Curr. Opin. Environ. Sustain. 2014, 8, 53–61. [Google Scholar] [CrossRef]
- Taube, F.; Gierus, M.; Hermann, A.; Loges, R.; Schönbach, P. Grassland and Globalization—Challenges for North-West European Grass and Forage Research. Grass Forage Sci. 2014, 69, 2–16. [Google Scholar] [CrossRef]
- Reinsch, T.; Loza, C.; Malisch, C.S.; Vogeler, I.; Kluß, C.; Loges, R.; Taube, F. Toward Specialized or Integrated Systems in Northwest Europe. Front. Sustain. Food Syst. 2021, 5, 383. [Google Scholar] [CrossRef]
- Ashman, T.-L.; Knight, T.M.; Steets, J.A.; Amarasekare, P.; Burd, M.; Campbell, D.R.; Dudash, M.R.; Johnston, M.O.; Mazer, S.J.; Mitchell, R.J.; et al. Pollen Limitation of Plant Reproduction. Ecology 2004, 85, 2408–2421. [Google Scholar] [CrossRef] [Green Version]
- Herbertsson, L.; Ekroos, J.; Albrecht, M.; Bartomeus, I.; Batáry, P.; Bommarco, R.; Caplat, P.; Diekötter, T.; Eikestam, J.M.; Entling, M.H.; et al. Bees Increase Seed Set of Wild Plants While the Proportion of Arable Land Has a Variable Effect on Pollination in European Agricultural Landscapes. Plant Ecol. Evol. 2021, 154, 341–350. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. Lond. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.D.; Howlett, B.G.; Winfree, R.; Cunningham, S.A.; Mayfield, M.M.; Arthur, A.D.; Andersson, G.K.S.; et al. Non-Bee Insects Are Important Contributors to Global Crop Pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleijn, D.; Winfree, R.; Bartomeus, I.; Carvalheiro, L.G.; Henry, M.; Isaacs, R.; Klein, A.-M.; Kremen, C.; M’Gonigle, L.K.; Rader, R.; et al. Delivery of Crop Pollination Services Is an Insufficient Argument for Wild Pollinator Conservation. Nat. Commun. 2015, 6, 550. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.; Steffan–Dewenter, I.; Tscharntke, T. Fruit Set of Highland Coffee Increases with the Diversity of Pollinating Bees. Proc. R. Soc. Lond. B 2003, 270, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Fijen, T.P.M.; Scheper, J.A.; Boom, T.M.; Janssen, N.; Raemakers, I.; Kleijn, D.; Irwin, R. Insect Pollination Is at Least as Important for Marketable Crop Yield as Plant Quality in a Seed Crop. Ecol. Lett. 2018, 21, 1704–1713. [Google Scholar] [CrossRef]
- Brittain, C.; Kremen, C.; Garber, A.; Klein, A.-M. Pollination and Plant Resources Change the Nutritional Quality of Almonds for Human Health. PLoS ONE 2014, 9, e90082. [Google Scholar] [CrossRef]
- Klatt, B.K.; Holzschuh, A.; Westphal, C.; Clough, Y.; Smit, I.; Pawelzik, E.; Tscharntke, T. Bee Pollination Improves Crop Quality, Shelf Life and Commercial Value. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132440. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The Global Stock of Domesticated Honey Bees Is Growing Slower than Agricultural Demand for Pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Diekötter, T.; Kadoya, T.; Peter, F.; Wolters, V.; Jauker, F. Oilseed Rape Crops Distort Plant-Pollinator Interactions. J. Appl. Ecol. 2010, 47, 209–214. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Alomar, D.; González-Estévez, M.A.; Traveset, A.; Lázaro, A. The Intertwined Effects of Natural Vegetation, Local Flower Community, and Pollinator Diversity on the Production of Almond Trees. Agric. Ecosyst. Environ. 2018, 264, 34–43. [Google Scholar] [CrossRef]
- Hegland, S.J.; Nielsen, A.; Lázaro, A.; Bjerknes, A.-L.; Totland, Ø. How Does Climate Warming Affect Plant-Pollinator Interactions? Ecol. Lett. 2009, 12, 184–195. [Google Scholar] [CrossRef]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed Coating with a Neonicotinoid Insecticide Negatively Affects Wild Bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWRs) Threatened and Endemic to Italy: Urgent Actions for Protection and Use. Biology 2022, 11, 193. [Google Scholar] [CrossRef]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; vanEngelsdorp, D.; Pettis, J.S.; Marion-Poll, F. High Levels of Miticides and Agrochemicals in North American Apiaries. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [Green Version]
- Pettis, J.S.; Lichtenberg, E.M.; Andree, M.; Stitzinger, J.; Rose, R.; vanEngelsdorp, D.; Nascimento, F.S. Crop Pollination Exposes Honey Bees to Pesticides Which Alters Their Susceptibility to the Gut Pathogen Nosema Ceranae. PLoS ONE 2013, 8, e70182. [Google Scholar] [CrossRef]
- Warzecha, D.; Diekötter, T.; Wolters, V.; Jauker, F. Intraspecific Body Size Increases with Habitat Fragmentation in Wild Bee Pollinators. Landsc. Ecol. 2016, 31, 1449–1455. [Google Scholar] [CrossRef]
- Grab, H.; Brokaw, J.; Anderson, E.; Gedlinske, L.; Gibbs, J.; Wilson, J.; Loeb, G.; Isaacs, R.; Poveda, K.; Diamond, S. Habitat Enhancements Rescue Bee Body Size from the Negative Effects of Landscape Simplification. J. Appl. Ecol. 2019, 56, 2144–2154. [Google Scholar] [CrossRef]
- Goulson, D.; Hanley, M.E.; Darvill, B.; Ellis, J.S.; Knight, M.E. Causes of Rarity in Bumblebees. Biol. Conserv. 2005, 122, 1–8. [Google Scholar] [CrossRef]
- Grass, I.; Albrecht, J.; Farwig, N.; Jauker, F. Plant Traits and Landscape Simplification Drive Intraspecific Trait Diversity of Bombus Terrestris in Wildflower Plantings. Basic Appl. Ecol. 2021, 57, 91–101. [Google Scholar] [CrossRef]
- Bommarco, R.; Lundin, O.; Smith, H.G.; Rundlöf, M. Drastic Historic Shifts in Bumble-Bee Community Composition in Sweden. Proc. R. Soc. B Biol. Sci. 2012, 279, 309–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleijn, D.; Baquero, R.A.; Clough, Y.; Díaz, M.; Esteban, J.; Fernández, F.; Gabriel, D.; Herzog, F.; Holzschuh, A.; Jöhl, R.; et al. Mixed Biodiversity Benefits of Agri-Environment Schemes in Five European Countries. Ecol. Lett. 2006, 9, 243–254; discussion 254–256. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, D.; Roschewitz, I.; Tscharntke, T.; Thies, C. Beta diversity at different spatial scales. Ecol. Appl. 2006, 16, 2011–2021. [Google Scholar] [CrossRef]
- Gabriel, D.; Tscharntke, T. Insect Pollinated Plants Benefit from Organic Farming. Agric. Ecosyst. Environ. 2007, 118, 43–48. [Google Scholar] [CrossRef]
- Klaus, V.H.; Kleinebecker, T.; Prati, D.; Gossner, M.M.; Alt, F.; Boch, S.; Gockel, S.; Hemp, A.; Lange, M.; Müller, J.; et al. Does Organic Grassland Farming Benefit Plant and Arthropod Diversity at the Expense of Yield and Soil Fertility? Agric. Ecosyst. Environ. 2013, 177, 1–9. [Google Scholar] [CrossRef]
- Power, E.F.; Stout, J.C. Organic Dairy Farming. J. Appl. Ecol. 2011, 48, 561–569. [Google Scholar] [CrossRef]
- Ebeling, A.; Klein, A.-M.; Schumacher, J.; Weisser, W.W.; Tscharntke, T. How Does Plant Richness Affect Pollinator Richness and Temporal Stability of Flower Visits? Oikos 2008, 117, 1808–1815. [Google Scholar] [CrossRef]
- Sutter, L.; Jeanneret, P.; Bartual, A.M.; Bocci, G.; Albrecht, M.; MacIvor, S. Enhancing Plant Diversity in Agricultural Landscapes Promotes Both Rare Bees and Dominant Crop-Pollinating Bees through Complementary Increase in Key Floral Resources. J. Appl. Ecol. 2017, 54, 1856–1864. [Google Scholar] [CrossRef] [Green Version]
- Orford, K.A. Modest Enhancements to Conventional Grassland Diversity Improve the Provision of Pollination Services. J. Appl. Ecol. 2016, 53, 906–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, S.G.; Woodcock, B.A.; Roberts, S.P.M.; Tscheulin, T.; Pilgrim, E.S.; Brown, V.K.; Tallowin, J.R. Enhancing Pollinator Biodiversity in Intensive Grasslands. J. Appl. Ecol. 2009, 46, 369–379. [Google Scholar] [CrossRef]
- Loges, R.; Mues, S.; Kluß, C.; Malisch, C.; Loza, C.; Poyda, A.; Reinsch, T.; Taube, F. Dairy Cows Back to Arable Regions? GRazing Leys for Eco-Efficient Milk Production Systems. Grassl. Sci. Eur. Meet. Future Demands Grassl. Prod. 2020, 25, 400–403. [Google Scholar]
- Loza, C.; Reinsch, T.; Loges, R.; Taube, F.; Gere, J.I.; Kluß, C.; Hasler, M.; Malisch, C.S. Methane Emission and Milk Production from Jersey Cows Grazing Perennial Ryegrass–White Clover and Multispecies Forage Mixtures. Agriculture 2021, 11, 175. [Google Scholar] [CrossRef]
- Smit, H.P.J.; Reinsch, T.; Kluß, C.; Loges, R.; Taube, F. Very Low Nitrogen Leaching in Grazed Ley-Arable-Systems in Northwest Europe. Agronomy 2021, 11, 2155. [Google Scholar] [CrossRef]
- Jauker, F.; Diekoetter, T.; Schwarzbach, F.; Wolters, V. Pollinator Dispersal in an Agricultural Matrix: Opposing Responses of Wild Bees and Hoverflies to Landscape Structure and Distance from Main Habitat. Landsc. Ecol. 2009, 24, 547–555. [Google Scholar] [CrossRef]
- Schall, R. Estimation in Generalized Linear Models with Random Effects. Biometrika 1991, 78, 719–727. [Google Scholar] [CrossRef]
- Laird, N.M.; Ware, J.H. Random-Effects Models for Longitudinal Data. Biometrics 1982, 38, 963–974. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: New York, NY, USA, 2006; ISBN 0-387-22747-4. [Google Scholar]
- Schaarschmidt, F.; Vaas, L. Analysis of Trials with Complex Treatment Structure Using Multiple Contrast Tests. Hortscience 2009, 44, 188–195. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S.; Schielzeth, H.; O’Hara, R.B. A General and Simple Method for Obtaining R2 from Generalized Linear Mixed-Effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Cochran, W.G. Analysis of Covariance. Biometrics 1957, 13, 261–281. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretz, F.; Hothorn, T.; Westfall, P. Multiple Comparisons Using R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2016; ISBN 978-1-4200-1090-9. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, R package version 2.5-7; University of Münster: Münster, Germany, 2013; Volume 2. [Google Scholar]
- Nyameasem, J.K.; Malisch, C.S.; Loges, R.; Taube, F.; Kluß, C.; Vogeler, I.; Reinsch, T. Nitrous Oxide Emission from Grazing Is Low across a Gradient of Plant Functional Diversity and Soil Conditions. Atmosphere 2021, 12, 223. [Google Scholar] [CrossRef]
- Lorenz, H.; Reinsch, T.; Kluß, C.; Taube, F.; Loges, R. Does the Admixture of Forage Herbs Affect the Yield Performance, Yield Stability and Forage Quality of a Grass Clover Ley? Sustainability 2020, 12, 5842. [Google Scholar] [CrossRef]
- Nyameasem, J.K.; Ben Halima, E.; Malisch, C.S.; Razavi, B.S.; Taube, F.; Reinsch, T. Nitrous Oxide Emission from Forage Plantain and Perennial Ryegrass Swards Is Affected by Belowground Resource Allocation Dynamics. Agronomy 2021, 11, 1936. [Google Scholar] [CrossRef]
- Scheper, J.; Holzschuh, A.; Kuussaari, M.; Potts, S.G.; Rundlöf, M.; Smith, H.G.; Kleijn, D. Environmental Factors Driving the Effectiveness of European Agri-Environmental Measures in Mitigating Pollinator Loss—A Meta-Analysis. Ecol. Lett. 2013, 16, 912–920. [Google Scholar] [CrossRef]
- Vickery, J.A.; Tallowin, J.R.; Feber, R.E.; Asteraki, E.J.; Atkinson, P.W.; Fuller, R.J.; Brown, V.K. The Management of Lowland Neutral Grasslands in Britain. J. Appl. Ecol. 2001, 38, 647–664. [Google Scholar] [CrossRef]
- European Commission; Statistical Office of the European Union. LUCAS: The EU’s Land Use and Land Cover Survey: 2021 Edition; Publications Office: Luxembourg, 2021. [Google Scholar]
- Peters, T.; Taube, F.; Kluß, C.; Reinsch, T.; Loges, R.; Fenger, F. How Does Nitrogen Application Rate Affect Plant Functional Traits and Crop Growth Rate of Perennial Ryegrass-Dominated Permanent Pastures? Agronomy 2021, 11, 2499. [Google Scholar] [CrossRef]
- Loges, R.; Loza, C.; Voss, P.; Kluß, C.; Malisch, C.; Taube, F. The Potential of Multispecies Swards for Eco-Efficient Dairy Productin in Northern Germany. In Meeting the Future Demands for Grassland Product, Proceedings of the 28th General Meeting of the European Grassland Federation, Helsinki, Finland, 19–22 October 2020; Natural Resources Institute Finland (Luke): Helsinki, Finland, 2020; p. 312. [Google Scholar]
- Cole, L.J.; Baddeley, J.A.; Robertson, D.; Topp, C.F.E.; Walker, R.L.; Watson, C.A. Supporting Wild Pollinators in Agricultural Landscapes through Targeted Legume Mixtures. Agric. Ecosyst. Environ. 2022, 323, 107648. [Google Scholar] [CrossRef]
- Diekötter, T.; Walther-Hellwig, K.; Frankl, R. Verbreitung, Lokale Häufigkeit Und Gefährdung Der Mooshummel (Bombus muscorum) Und Der Waldhummel (Bombus sylvarum) Im Amöneburger Becken, Jahrb. Nat. Hess. 2001, 6, 92–99. [Google Scholar]
- Hanley, M.E.; Franco, M.; Pichon, S.; Darvill, B.; Goulson, D. Breeding System, Pollinator Choice and Variation in Pollen Quality in British Herbaceous Plants. Funct. Ecol. 2008, 22, 592–598. [Google Scholar] [CrossRef]
- Pywell, R.F.; Warman, E.A.; Carvell, C.; Sparks, T.H.; Dicks, L.V.; Bennett, D.; Wright, A.; Critchley, C.N.R.; Sherwood, A. Providing Foraging Resources for Bumblebees in Intensively Farmed Landscapes. Biol. Conserv. 2005, 121, 479–494. [Google Scholar] [CrossRef]
- Memmott, J.; Waser, N.M.; Price, M.V. Tolerance of Pollination Networks to Species Extinctions. Proc. R. Soc. Lond. B 2004, 271, 2605–2611. [Google Scholar] [CrossRef] [PubMed]
- Sidemo-Holm, W.; Carrié, R.; Ekroos, J.; Lindström, S.A.; Smith, H.G. Reduced Crop Density Increases Floral Resources to Pollinators without Affecting Crop Yield in Organic and Conventional Fields. J. Appl. Ecol. 2021, 58, 1421–1430. [Google Scholar] [CrossRef]
- Happe, A.-K.; Riesch, F.; Rösch, V.; Gallé, R.; Tscharntke, T.; Batáry, P. Small-Scale Agricultural Landscapes and Organic Management Support Wild Bee Communities of Cereal Field Boundaries. Agric. Ecosyst. Environ. 2018, 254, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Holzschuh, A.; Steffan-Dewenter, I.; Kleijn, D.; Tscharntke, T. Diversity of Flower-visiting Bees in Cereal Fields: Effects of Farming System, Landscape Composition and Regional Context. J. Appl. Ecol. 2007, 44, 41–49. [Google Scholar] [CrossRef]
- Kaiser-Bunbury, C.N.; Muff, S.; Memmott, J.; Müller, C.B.; Caflisch, A. The Robustness of Pollination Networks to the Loss of Species and Interactions. Ecol. Lett. 2010, 13, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Dupont, Y.L.; Damgaard, C.; Simonsen, V.; Stout, J.C. Quantitative Historical Change in Bumblebee (Bombus Spp.) Assemblages of Red Clover Fields. PLoS ONE 2011, 6, e25172. [Google Scholar] [CrossRef] [Green Version]
- Diekötter, T. Effects of Landscape Elements on the Distribution of the Rare Bumblebee Species Bombus muscorum in an Agricultural Landscape. Biodivers. Conserv. 2006, 15, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Westphal, C.; Steffan-Dewenter, I.; Tscharntke, T. Mass Flowering Crops Enhance Pollinator Densities at a Landscape Scale. Ecol. Lett. 2003, 6, 961–965. [Google Scholar] [CrossRef]
- Westrich, P. Wildbienen Baden-Württembergs; Ulmer, E., Ed.; Eugen Ulmer: Stuttgart, Germany, 1990; ISBN 3-8001-3307-5. [Google Scholar]
- Wan, H.; Bai, Y.; Hooper, D.U.; Schönbach, P.; Gierus, M.; Schiborra, A.; Taube, F. Selective Grazing and Seasonal Precipitation Play Key Roles in Shaping Plant Community Structure of Semi-Arid Grasslands. Landsc. Ecol. 2015, 30, 1767–1782. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beye, H.; Taube, F.; Lange, K.; Hasler, M.; Kluß, C.; Loges, R.; Diekötter, T. Species-Enriched Grass-Clover Mixtures Can Promote Bumblebee Abundance Compared with Intensively Managed Conventional Pastures. Agronomy 2022, 12, 1080. https://doi.org/10.3390/agronomy12051080
Beye H, Taube F, Lange K, Hasler M, Kluß C, Loges R, Diekötter T. Species-Enriched Grass-Clover Mixtures Can Promote Bumblebee Abundance Compared with Intensively Managed Conventional Pastures. Agronomy. 2022; 12(5):1080. https://doi.org/10.3390/agronomy12051080
Chicago/Turabian StyleBeye, Henriette, Friedhelm Taube, Katharina Lange, Mario Hasler, Christof Kluß, Ralf Loges, and Tim Diekötter. 2022. "Species-Enriched Grass-Clover Mixtures Can Promote Bumblebee Abundance Compared with Intensively Managed Conventional Pastures" Agronomy 12, no. 5: 1080. https://doi.org/10.3390/agronomy12051080