
Yields and Nitrogen Dynamics in Ley-Arable Systems—Comparing Different Approaches in the APSIM Model

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Experimental Sites
2.2. Description of the APSIM Model
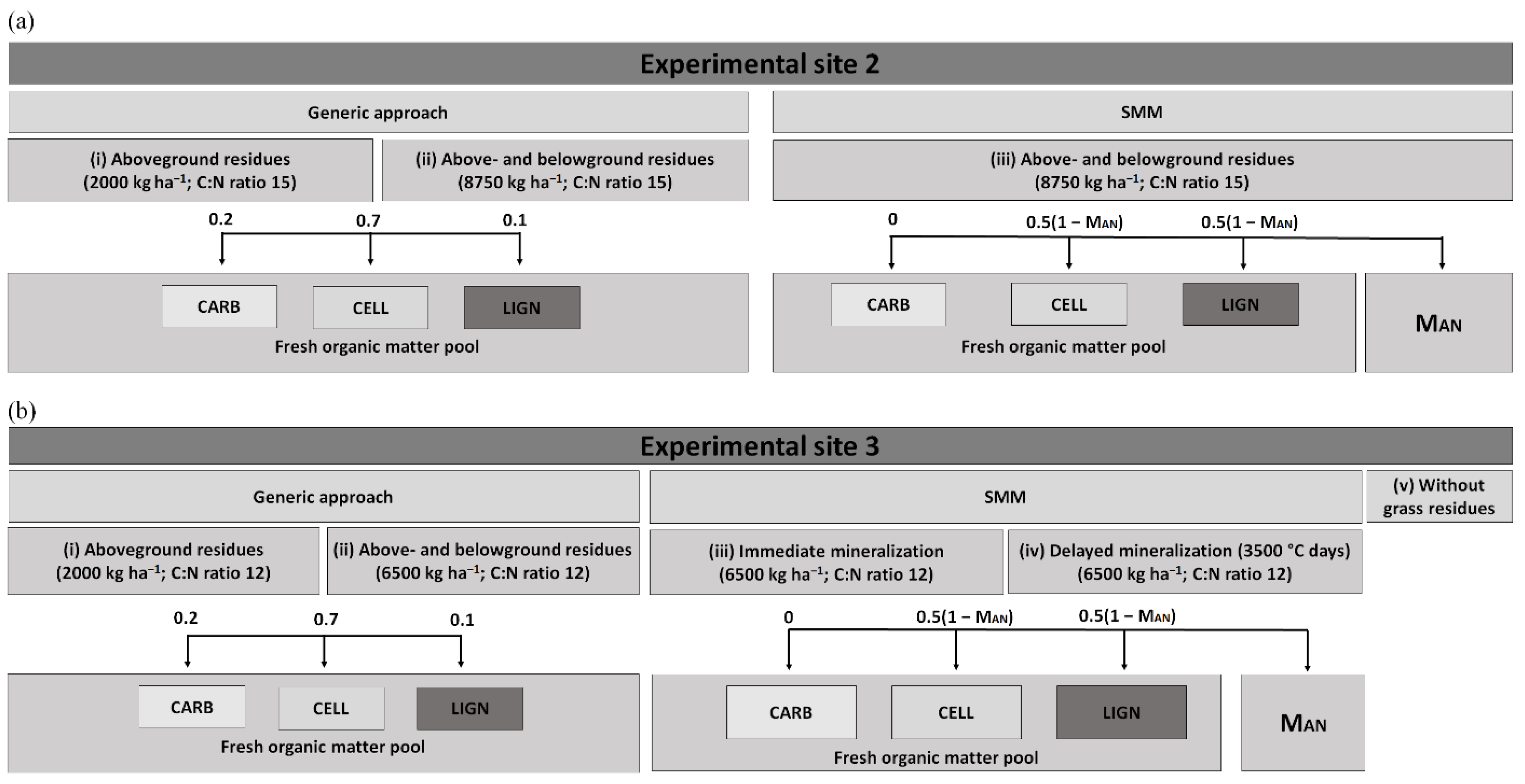
2.3. Description of the Model Set Up
2.4. Model Adaptations to Predict Residual Nitrogen Effect
2.5. Model Evaluation
3. Results
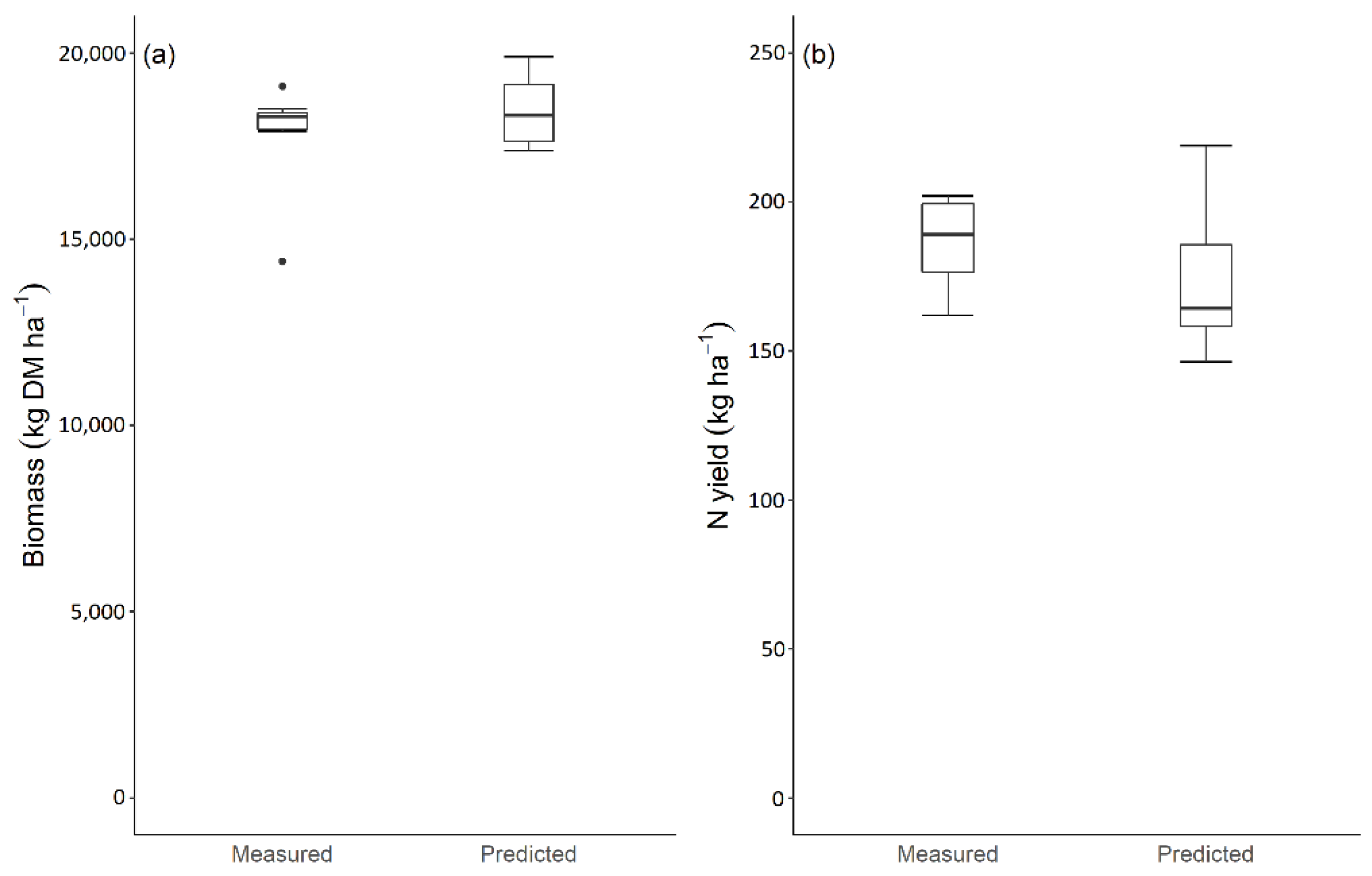
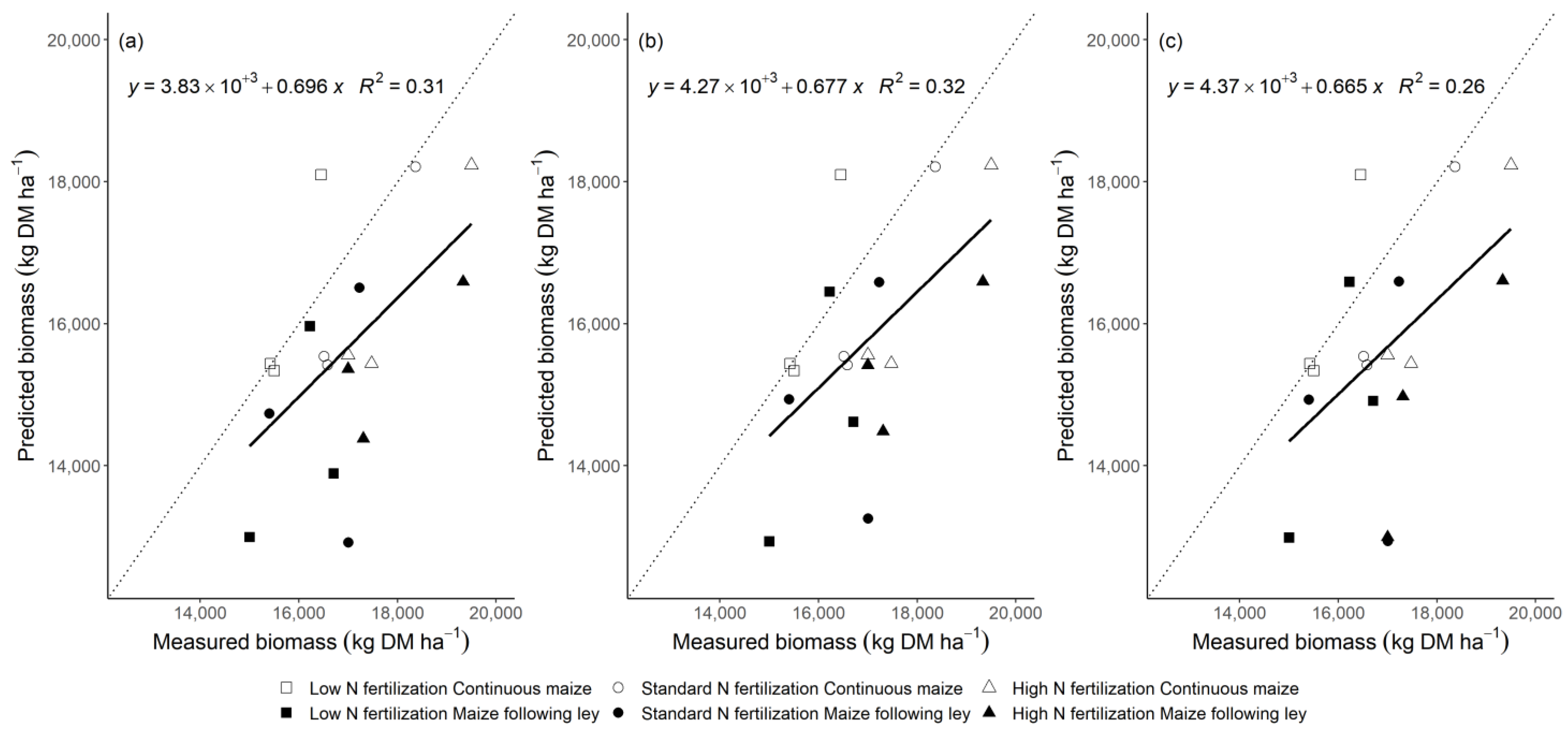
3.1. Model Performance to Predict Maize Biomass Yield
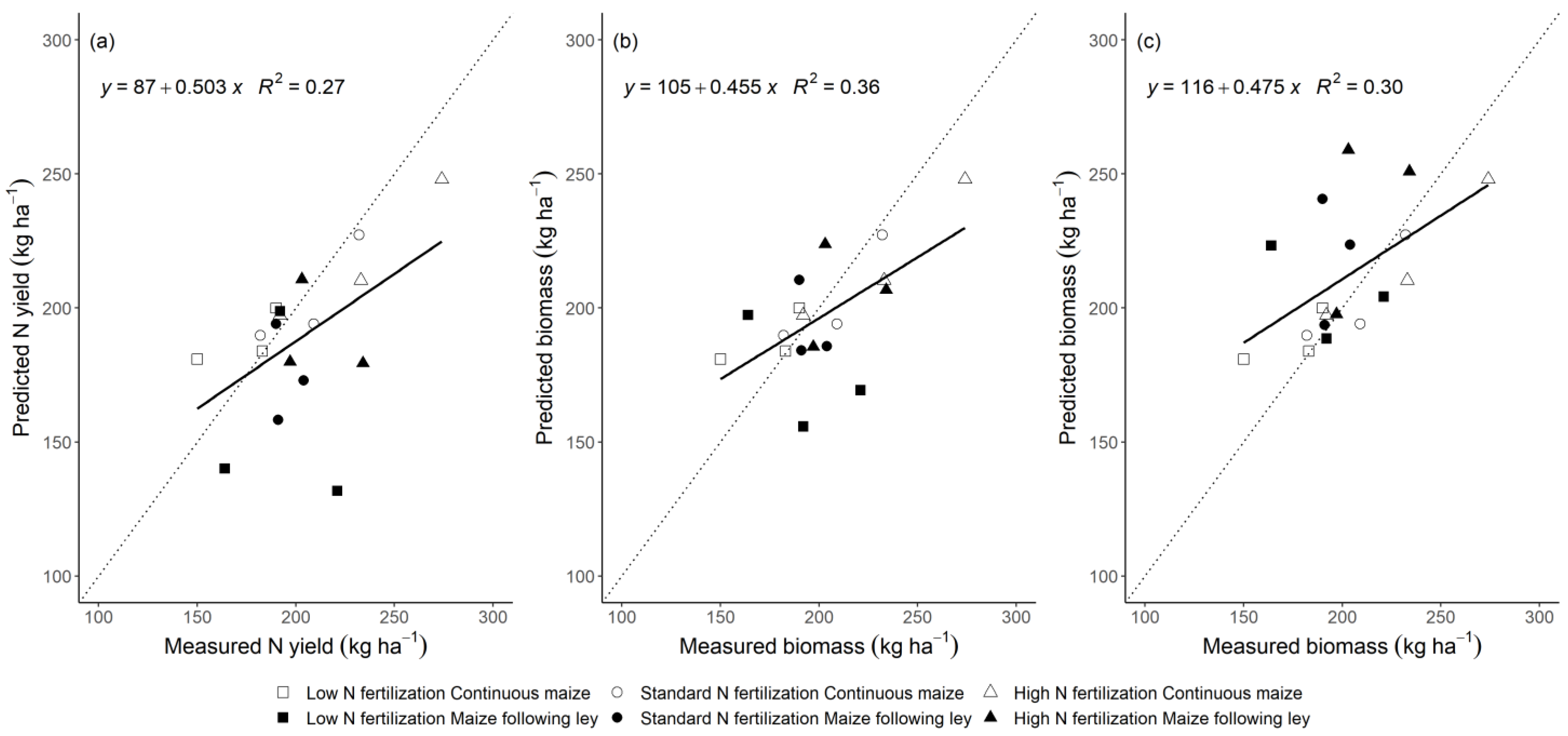
3.2. Model Performance to Predict Maize Nitrogen Yield
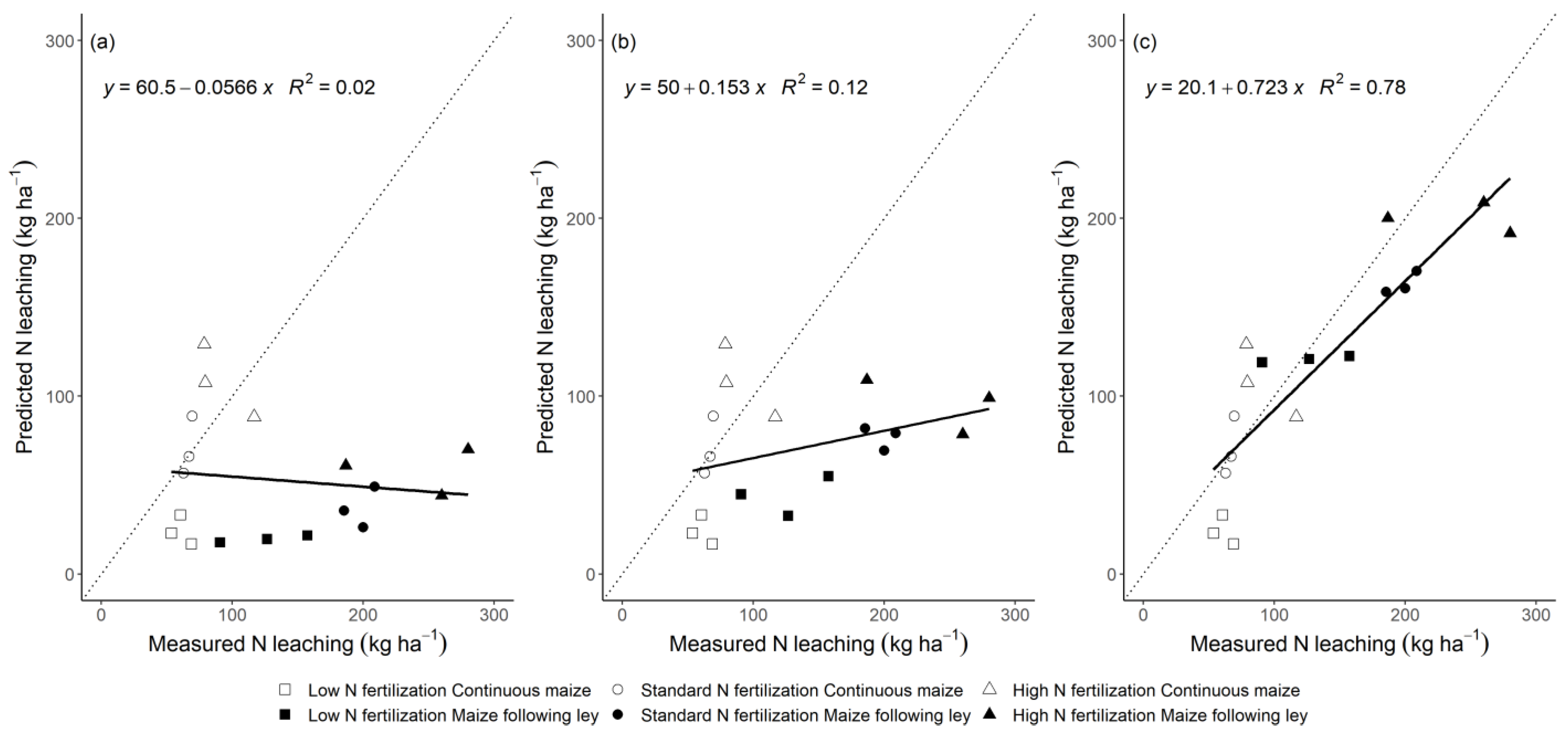
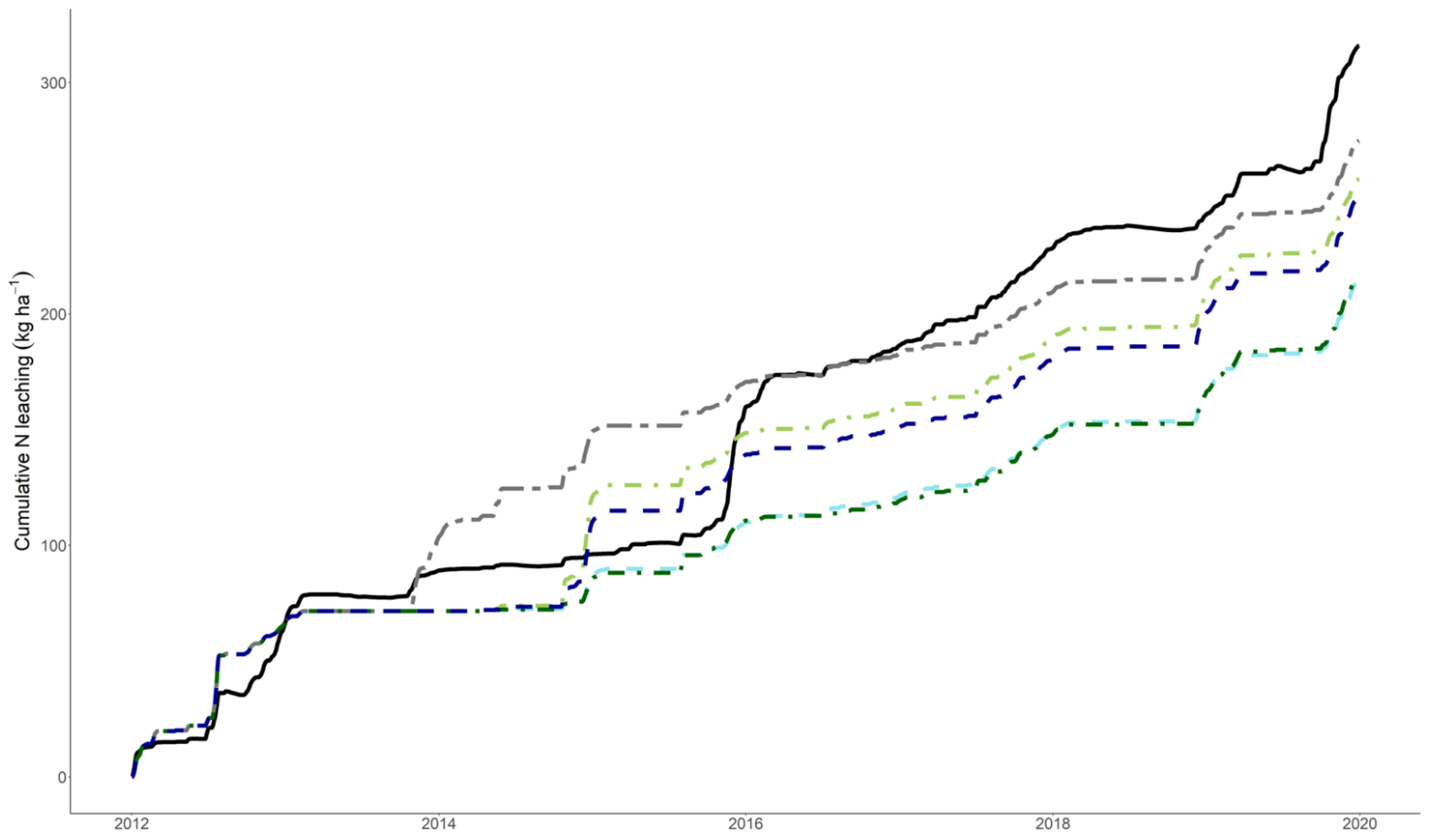
3.3. Model Performance to Predict Nitrogen Leaching
4. Discussion
4.1. Overall Model Performance of APSIM to Simulate Biomass Yield, N Yield, and N Leaching in Maize Systems in Northwest Europe
4.2. Belowground Biomass and Residual Nitrogen Effect of Ley Pastures
4.3. Comparison of the Generic and the Modified Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Boer, I.J.M.; van Ittersum, M.K. Circularity in Agricultural Production; Wageningen University & Research: Wageningen, The Netherlands, 2018; pp. 1–74. [Google Scholar]
- Velthof, G.L.; Lesschen, J.P.; Webb, J.; Pietrzak, S.; Miatkowski, Z.; Pinto, M.; Kros, J.; Oenema, O. Science of the Total Environment The impact of the Nitrates Directive on nitrogen emissions from agriculture in the EU-27 during 2000–2008. Sci. Total Environ. 2014, 468–469, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Wachendorf, M.; Buchter, M.; Trott, H.; Taube, F. Performance and environmental effects of forage production on sandy soils. II. Impact of defoliation system and nitrogen input on nitrate leaching losses. Grass Forage Sci. 2004, 59, 56–68. [Google Scholar] [CrossRef]
- Abalos, D.; van Groenigen, J.W.; Philippot, L.; Lubbers, I.M.; De Deyn, G.B. Plant trait-based approaches to improve nitrogen cycling in agroecosystems. J. Appl. Ecol. 2019, 56, 2454–2466. [Google Scholar] [CrossRef]
- Lemaire, G.; Franzluebbers, A.; de Faccio Carvalho, P.C.; Dedieu, B. Integrated crop-livestock systems: Strategies to achieve synergy between agricultural production and environmental quality. Agric. Ecosyst. Environ. 2014, 190, 4–8. [Google Scholar] [CrossRef]
- Biernat, L.; Taube, F.; Vogeler, I.; Reinsch, T.; Kluß, C.; Loges, R. Is organic agriculture in line with the EU-Nitrate directive? On-farm nitrate leaching from organic and conventional arable crop rotations. Agric. Ecosyst. Environ. 2020, 298, 106964. [Google Scholar] [CrossRef]
- Guillaume, M.; Durand, J.; Duru, M.; Gastal, F.; Julier, B.; Litrico, I.; Louarn, G.; Médiène, S.; Moreau, D.; Valentin-morison, M.; et al. Role of ley pastures in tomorrow’s cropping systems. A review. Agron. Sustain. Dev. 2020, 40, 17. [Google Scholar] [CrossRef]
- Crème, A.; Rumpel, C.; Le Roux, X.; Romian, A.; Lan, T.; Chabbi, A. Ley grassland under temperate climate had a legacy effect on soil organic matter quantity, biogeochemical signature and microbial activities. Soil Biol. Biochem. 2018, 122, 203–210. [Google Scholar] [CrossRef]
- van Eekeren, N.; Bommelé, L.; Bloem, J.; Schouten, T.; Rutgers, M.; de Goede, R.; Reheul, D.; Brussaard, L. Soil biological quality after 36 years of ley-arable cropping, permanent grassland and permanent arable cropping. Appl. Soil Ecol. 2008, 40, 432–446. [Google Scholar] [CrossRef] [Green Version]
- Soussana, J.F.; Lemaire, G. Coupling carbon and nitrogen cycles for environmentally sustainable intensification of grasslands and crop-livestock systems. Agric. Ecosyst. Environ. 2014, 190, 9–17. [Google Scholar] [CrossRef]
- Christensen, B.T.; Rasmussen, J.; Eriksen, J.; Hansen, E.M. Soil carbon storage and yields of spring barley following grass leys of different age. Eur. J. Agron. 2009, 31, 29–35. [Google Scholar] [CrossRef]
- Vertès, F.; Hatch, D.; Velthof, G.; Taube, F.; Laurent, F.; Loiseau, P.; Recous, S. Short-Term and Cumulative Effects of Grassland Cultivation on Nitrogen and Carbon Cycling in Ley-Arable Rotations. Grassl. Sci. Eur. 2007, 12, 227–246. [Google Scholar]
- Nevens, F.; Reheul, D. The nitrogen- and non-nitrogen-contribution effect of ploughed grass leys on the following arable forage crops: Determination and optimum use. Eur. J. Agron. 2002, 16, 57–74. [Google Scholar] [CrossRef]
- Loges, R.; Bunne, I.; Reinsch, T.; Malisch, C.; Kluß, C.; Herrmann, A.; Taube, F. Forage production in rotational systems generates similar yields compared to maize monocultures but improves soil carbon stocks. Eur. J. Agron. 2018, 97, 11–19. [Google Scholar] [CrossRef]
- Deru, J.; Schilder, H.; van der Schoot, J.R.; van Eekeren, N. Genetic differences in root mass of Lolium perenne varieties under field conditions. Euphytica 2014, 199, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Iepema, G.L.; Deru, J.G.C.; Hoekstra, N.J.; Van Eekeren, N.; Höglind, M.; Bakken, A.K.; Hovstad, K.A.; Kallioniemi, E.; Riley, H.; Steinshamn, H.; et al. Rooting of permanent grassland in relation to build-up of soil organic matter for climate mitigation. In The Multiple Roles of Grassland in the European Bioeconomy, Proceedings of the 26th General Meeting of the European Grassland Federation, Trondheim, Norway, 4–8 September 2016; Höglind, M., Bakken, A.K., Hovstad, K.A., Kallioniemi, E., Riley, H., Steinshamn, H., Østrem, L., Eds.; NIBIO, The Norwegian Institute of Bioeconomy Research: Trondheim, Norway, 2016; pp. 777–779. [Google Scholar]
- Hoffmann, M.P.; Isselstein, J.; Rötter, R.P.; Kayser, M. Agriculture, Ecosystems and Environment Nitrogen management in crop rotations after the break-up of grassland: Insights from modelling. Agric. Ecosyst. Environ. 2018, 259, 28–44. [Google Scholar] [CrossRef]
- Watson, C.A.; Baddeley, J.A.; Edwards, A.C.; Rees, R.M.; Walker, R.L.; Topp, C.F.E. Influence of ley duration on the yield and quality of the subsequent cereal crop (spring oats) in an organically managed long-term crop rotation experiment. Org. Agric. 2011, 1, 147–159. [Google Scholar] [CrossRef]
- Holzworth, D.P.; Huth, N.I.; DeVoil, P.G.; Zurcher, E.J.; Herrmann, N.I.; McLean, G.; Chenu, K.; van Oosterom, E.J.; Snow, V.; Murphy, C.; et al. APSIM—Evolution towards a new generation of agricultural systems simulation. Environ. Model. Softw. 2014, 62, 327–350. [Google Scholar] [CrossRef]
- Moeller, C.; Pala, M.; Manschadi, A.M.; Meinke, H.; Sauerborn, J. Assessing the sustainability of wheat-based cropping systems using APSIM: Model parameterisation and evaluation. Aust. J. Agric. Res. 2007, 58, 75–86. [Google Scholar] [CrossRef]
- Böldt, M.; Taube, F.; Vogeler, I.; Reinsch, T.; Kluß, C.; Loges, R. Evaluating Different Catch Crop Strategies for Closing the Nitrogen Cycle in Cropping Systems—Field Experiments and Modelling. Sustainability 2021, 13, 394. [Google Scholar] [CrossRef]
- Vogeler, I.; Kaag, I.; Friedhelm, T.; Poulsen, V.; Loges, R.; Møller, E. Effect of winter cereal sowing time on yield and nitrogen leaching based on experiments and modelling. Soil Use Manag. 2021, 38, 663–675. [Google Scholar] [CrossRef]
- Manevski, K.; Børgesen, C.D.; Andersen, M.N.; Kristensen, I.S. Reduced nitrogen leaching by intercropping maize with red fescue on sandy soils in North Europe: A combined field and modeling study. Plant Soil 2015, 388, 67–85. [Google Scholar] [CrossRef]
- Hammer, G.L.; Kropff, M.J.; Sinclair, T.R.; Porter, J.R. Future contributions of crop modelling from heuristics and supporting decision making to understanding genetic regulation and aiding crop improvement. Eur. J. Agron. 2002, 18, 15–31. [Google Scholar] [CrossRef]
- Soufizadeh, S.; Munaro, E.; Mclean, G.; Massignam, A.; Van Oosterom, E.J.; Chapman, S.C.; Messina, C.; Cooper, M.; Hammer, G.I. Modelling the nitrogen dynamics of maize crops—Enhancing the APSIM maize model. Eur. J. Agron. 2018, 100, 118–131. [Google Scholar] [CrossRef]
- Schrama, M.; De Haan, J.J.; Kroonen, M.; Verstegen, H.; Putten, W.H. Van Der Crop yield gap and stability in organic and conventional farming systems. Agric. Ecosyst. Environ. 2018, 256, 123–130. [Google Scholar] [CrossRef]
- Keating, B.A.; Carberry, P.S.; Hammer, G.L.; Probert, M.E.; Robertson, M.J.; Holzworth, D.; Huth, N.I.; Hargreaves, J.N.G.; Meinke, H.; Hochman, Z.; et al. An overview of APSIM, a model designed for farming systems simulation. Eur. J. Agron. 2003, 18, 267–288. [Google Scholar] [CrossRef] [Green Version]
- Probert, M.E.; Dimes, J.P.; Keating, B.A.; Dalal, R.C.; Strong, W.M. APSIM’s Water and Nitrogen Modules and Simulation of the Dynamics of Water and Nitrogen in Fallow Systems. Agric. Syst. 1998, 56, 1–28. [Google Scholar] [CrossRef]
- Snow, V.; Huth, N. The APSIM-Micromet Module; HortResearch Internal Report No. 2004/12848; HortResearch: Palmerston North, New Zealand, 2004. [Google Scholar]
- Foley, J.; Fainges, J. Soil Evaporation: How Much Water Is Lost from Northern Crop Systems and Do Agronomic Models Accurately Represent This Loss? 2014. Available online: https://grdc.com.au/resources-and-publications/grdc-update-papers (accessed on 16 March 2022).
- Dalgliesh, N.; Hochman, Z.; Huth, N.; Holzworth, D. A Protocol for the Development of APSoil Parameter Values for Use in APSIM; Version 4; CSIRO: Canberra, Australia, 2016. [Google Scholar]
- Ellermann, T.; Nygaard, J.; Christensen, J.H.; Løfstrøm, P.; Geels, C.; Nielsen, I.E.; Poulsen, M.B.; Monies, C.; Gyldenkærne, S.; Brandt, J.; et al. Nitrogen Deposition on Danish Nature. Atmosphere 2018, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Moore, A.D.; Holzworth, D.P.; Herrmann, N.I.; Brown, H.E.; De Voil, P.G.; Snow, V.O.; Zurcher, E.J.; Huth, N.I. Environmental Modelling & Software Modelling the manager: Representing rule-based management in farming systems simulation models. Environ. Model. Softw. 2014, 62, 399–410. [Google Scholar] [CrossRef]
- Acharya, B.S.; Rasmussen, J.; Eriksen, J. Grassland carbon sequestration and emissions following cultivation in a mixed crop rotation. Agric. Ecosyst. Environ. 2012, 153, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Vogeler, I.; Cichota, R.; Thomsen, I.K.; Bruun, S.; Jensen, L.S.; Pullens, J.W.M. Estimating nitrogen release from Brassicacatch crop residues—Comparison of different approaches within the APSIM model. Soil Tillage Res. 2019, 195, 104358. [Google Scholar] [CrossRef]
- Hansen, J.P.; Eriksen, J.; Jensen, L.S. Residual nitrogen effect of a dairy crop rotation as influenced by grass-clover ley management, manure type and age. Soil Use Manag. 2005, 21, 278–286. [Google Scholar] [CrossRef]
- Moriasi, D.N.; Arnold, J.G.; Van Liew, M.W.; Bingner, R.L.; Harmel, R.D.; Veith, T.L. Model Evaluation Guidelines for Systematic Quantification of Accuracy in Watershed Simulations. Trans. ASABE 2007, 50, 885–900. [Google Scholar] [CrossRef]
- Kollas, C.; Kersebaum, K.C.; Nendel, C.; Manevski, K.; Müller, C.; Palosuo, T.; Armas-Herrera, C.M.; Beaudoin, N.; Bindi, M.; Charfeddine, M.; et al. Crop rotation modelling-A European model intercomparison. Eur. J. Agron. 2015, 70, 98–111. [Google Scholar] [CrossRef]
- Salo, T.J.; Palosuo, T.; Kersebaum, K.C.; Nendel, C.; Angulo, C.; Ewert, F.; Bindi, M.; Calanca, P.; Klein, T.; Moriondo, M.; et al. Comparing the performance of 11 crop simulation models in predicting yield response to nitrogen fertilization. J. Agric. Sci. 2016, 154, 1218–1240. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Kersebaum, K.C.; Kollas, C.; Baby, S.; Beaudoin, N.; Manevski, K.; Palosuo, T.; Nendel, C.; Wu, L.; Hoffmann, M.; et al. Multi-model uncertainty analysis in predicting grain N for crop rotations in Europe. Eur. J. Agron. 2017, 84, 152–165. [Google Scholar] [CrossRef]
- Yin, X.; Kersebaum, K.C.; Kollas, C.; Manevski, K.; Baby, S.; Beaudoin, N.; Öztürk, I.; Gaiser, T.; Wu, L.; Hoffmann, M.; et al. Performance of process-based models for simulation of grain N in crop rotations across Europe. Agric. Syst. 2017, 154, 63–77. [Google Scholar] [CrossRef]
- Wolf, J.; Broeke, M.J.D.H.; Ro, R. Simulation of nitrogen leaching in sandy soils in The Netherlands with the ANIMO model and the integrated modelling system STONE. Agric. Ecosyst. Environ. 2005, 105, 523–540. [Google Scholar] [CrossRef]
- Liu, H.L.; Yang, J.Y.; Drury, C.F.; Reynolds, W.D.; Tan, C.S.; Bai, Y.L.; He, P.; Jin, J.; Hoogenboom, G. Using the DSSAT-CERES-Maize model to simulate crop yield and nitrogen cycling in fields under long-term continuous maize production. Nutr. Cycl. Agroecosystems 2011, 89, 313–328. [Google Scholar] [CrossRef]
- Peake, A.S.; Huth, N.I.; Carberry, P.S.; Raine, S.R.; Smith, R.J. Quantifying potential yield and lodging-related yield gaps for irrigated spring wheat in sub-tropical Australia. Field Crops Res. 2014, 158, 1–14. [Google Scholar] [CrossRef]
- Yang, X.; Zheng, L.; Yang, Q.; Wang, Z.; Cui, S.; Shen, Y. Modelling the effects of conservation tillage on crop water productivity, soil water dynamics and evapotranspiration of a maize-winter wheat-soybean rotation system on the Loess Plateau of China using APSIM. Agric. Syst. 2018, 166, 111–123. [Google Scholar] [CrossRef]
- Wallach, D.; Palosuo, T.; Thorburn, P.; Hochman, Z.; Andrianasolo, F.; Asseng, S.; Basso, B.; Buis, S.; Crout, N.; Dumont, B.; et al. Multi-model evaluation of phenology prediction for wheat in Australia. Agric. For. Meteorol. 2021, 298–299, 108289. [Google Scholar] [CrossRef]
- Archontoulis, S.V.; Castellano, M.J.; Licht, M.A.; Nichols, V.; Baum, M.; Huber, I.; Martinez-Feria, R.; Puntel, L.; Ordóñez, R.A.; Iqbal, J.; et al. Predicting crop yields and soil-plant nitrogen dynamics in the US Corn Belt. Crop Sci. 2020, 60, 721–738. [Google Scholar] [CrossRef] [Green Version]
- Parent, B.; Leclere, M.; Lacube, S.; Semenov, M.A.; Welcker, C.; Martre, P.; Tardieu, F. Maize yields over Europe may increase in spite of climate change, with an appropriate use of the genetic variability of flowering time. Proc. Natl. Acad. Sci. USA 2018, 115, 10642–10647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhavizadegan, F.; Ansarifar, J.; Wang, L.; Huber, I.; Archontoulis, S.V. A time-dependent parameter estimation framework for crop modeling. Sci. Rep. 2021, 11, 11437. [Google Scholar] [CrossRef]
- Wallach, D.; Palosuo, T.; Thorburn, P.; Hochman, Z.; Gourdain, E.; Andrianasolo, F.; Asseng, S.; Basso, B.; Buis, S.; Crout, N.; et al. The chaos in calibrating crop models: Lessons learned from a multi-model calibration exercise. Environ. Model. Softw. 2021, 145, 105206. [Google Scholar] [CrossRef]
- Wang, E.; Martre, P.; Zhao, Z.; Ewert, F.; Maiorano, A.; Rötter, R.P.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. The uncertainty of crop yield projections is reduced by improved temperature response functions. Nat. Plants 2017, 3, 17102. [Google Scholar] [CrossRef] [Green Version]
- Basso, B.; Liu, L.; Ritchie, J.T. A Comprehensive Review of the CERES-Wheat, -Maize and -Rice Models’ Performances. Adv. Agron. 2016, 136, 27–132. [Google Scholar] [CrossRef]
- Weihermüller, L.; Siemens, J.; Deurer, M.; Knoblauch, S.; Rupp, H.; Göttlein, A.; Pütz, T. In Situ Soil Water Extraction: A Review. J. Environ. Qual. 2007, 36, 1735–1748. [Google Scholar] [CrossRef]
- Dietzel, R.; Liebman, M.; Ewing, R.; Helmers, M.; Horton, R.; Jarchow, M.; Archontoulis, S. How efficiently do corn- and soybean-based cropping systems use water? A systems modeling analysis. Glob. Chang. Biol. 2016, 22, 666–681. [Google Scholar] [CrossRef]
- Li, F.Y.; Newton, P.C.D.; Lieffering, M. Testing simulations of intra- and inter-annual variation in the plant production response to elevated CO2 against measurements from an 11-year FACE experiment on grazed pasture. Glob. Chang. Biol. 2014, 20, 228–239. [Google Scholar] [CrossRef]
- Vogeler, I.; Lucci, G.; Shephard, M. Effects of fertiliser nitrogen management on nitrate leaching risk from grazed dairy pasture An assessment of the effects of fertilizer nitrogen management on nitrate leaching risk from grazed dairy pasture. J. Agric. Sci. 2016, 154, 407–424. [Google Scholar] [CrossRef]
- Li, F.Y.; Snow, V.O.; Holzworth, D.P. Modelling the seasonal and geographical pattern of pasture production in New Zealand. New Zeal. J. Agric. Res. 2011, 54, 331–352. [Google Scholar] [CrossRef] [Green Version]
- Attard, E.; Le Roux, X.; Charrier, X.; Delfosse, O.; Guillaumaud, N.; Lemaire, G.; Recous, S. Delayed and asymmetric responses of soil C pools and N fluxes to grassland/cropland conversions. Soil Biol. Biochem. 2016, 97, 31–39. [Google Scholar] [CrossRef]
- Eriksen, J. Nitrate leaching and growth of cereal crops following cultivation of contrasting temporary grasslands. J. Agric. Sci. 2001, 136, 271–281. [Google Scholar] [CrossRef]
- Garwood, E.A.; Clement, C.R.; Williams, T.E. Leys and soil organic matter. J. Agric. Sci. 1972, 78, 333–341. [Google Scholar] [CrossRef]
- Chen, S.M.; Lin, S.; Loges, R.; Reinsch, T.; Hasler, M.; Taube, F. Independence of seasonal patterns of root functional traits and rooting strategy of a grass-clover sward from sward age and slurry application. Grass Forage Sci. 2016, 71, 607–621. [Google Scholar] [CrossRef]
- Kayser, M.; Seidel, K.; Müller, J.; Isselstein, J. The effect of succeeding crop and level of N fertilization on N leaching after break-up of grassland. Eur. J. Agron. 2008, 29, 200–207. [Google Scholar] [CrossRef]
- Schils, R.L.M.; Aarts, H.F.M.; Bussink, D.W.; Conijn, J.G.; Corré, W.J.; Van Dam, A.M.; Hoving, I.E.; Van Der Meer, H.G.; Velthof, G.L. Grassland renovation in the Netherlands; agronomic, environmental and economic issues. Grassl. Resowing Grass-Arable Crop Rotat. 2002, 47, 9–24. [Google Scholar]
- Eriksen, J.; Askegaard, M.; Søegaard, K. Residual effect and nitrate leaching in grass-arable rotations: Effect of grassland proportion, sward type and fertilizer history. Soil Use Manag. 2008, 24, 373–382. [Google Scholar] [CrossRef]
- Kunrath, T.R.; de Berranger, C.; Charrier, X.; Gastal, F.; de Faccio Carvalho, P.C.; Lemaire, G.; Emile, J.C.; Durand, J.L. How much do sod-based rotations reduce nitrate leaching in a cereal cropping system? Agric. Water Manag. 2015, 150, 46–56. [Google Scholar] [CrossRef]
- Probert, M.E.; Delve, R.J.; Kimani, S.K.; Dimes, J.P. Modelling nitrogen mineralization from manures: Representing quality aspects by varying C:N ratio of sub-pools. Soil Biol. Biochem. 2005, 37, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, M.; Reddy, K.S.; Probert, M.E.; Dalal, R.C.; Rao, A.S.; Menzies, N.W. Modelling N mineralization from green manure and farmyard manure from a laboratory incubation study. Ecol. Model. 2011, 222, 719–726. [Google Scholar] [CrossRef]
- Vogeler, I.; Böldt, M.; Taube, F. Mineralisation of catch crop residues and N transfer to the subsequent crop. Sci. Total Environ. 2022, 810, 152142. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Site 1 (E1) | Experimental Site 2 (E2) | Experimental Site 3 (E3) | |
|---|---|---|---|
| Location | Vredepeel, Netherlands (51.32° N, 5.32° E) | Jyndevad, Denmark (54.54° N, 9.46° E) | Schuby, Germany (54.31° N, 9.26° E). |
| Modeled cropping system | Single forage maize years (7 year) in a crop rotation | Continous forage maize and a ley-forage maize system | Continous cropping system (6 year forage maize, 2 year winter cereals, and once a ley period) |
| Soil texture | 92% sand, 7% silt, and 1% clay | 89% sand, 7% silt, and 4% clay | 84% sand, 11% silt, and 5% clay |
| Organic carbon content 1 | 2.3% | 3.0% | 3.0% |
| Soil pH 1 | 5.6 | 7.0 | 6.0 |
| Mean annual precipitation 2 | 661 mm | 973 mm | 895 mm |
| Mean annual temperature 2 | 10.6 °C | 7.9 °C | 8.6 °C |
| Category | Coefficient/Variable | Default | Fitted |
|---|---|---|---|
| Thermal time units | Time lag before linear coleoptile growth starts | 15 | 50 |
| tt_emerg_to_endjuv | 135 | 200 | |
| tt_endjuv_to_init | 180 | 135 | |
| tt_flower_to_maturity | 990 | 700 | |
| Biomass | Radiation use efficiency | 1.6 | 1.7 |
| Phyllochron interval | Leaf_app_rate1 | 65 | 60 |
| Grain yield | GNmaxCoef | 170 | 220 |
| DM | N Yield | |
|---|---|---|
| R2 | 0.16 | 0.04 |
| RMSE | 1506 | 38 |
| NSE | −0.11 | −1.95 |
| Pbias | 3.80 | −7.50 |
| Generic Approach | Generic Approach | SMM | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Aboveground | Above- and Belowground | ||||||||
| DM | N Yield | N Leaching | DM | N Yield | N Leaching | DM | N Yield | N Leaching | |
| RMSE | 1846 | 30 | 113 | 1735 | 23 | 90 | 1920 | 27 | 37 |
| NSE | −1.32 | −0.18 | −1.51 | −1.05 | 0.30 | −0.61 | −1.51 | 0.09 | 0.72 |
| Pbias | −7.70 | −6.70 | −59.40 | −7.10 | −2.50 | −46.40 | −7.60 | 4.70 | −12.30 |
| Generic Approach | Generic Approach | SMM | SMM | Without a Ryegrass Period | |
|---|---|---|---|---|---|
| Aboveground | Above- and Belowground | Immediate Mineralization | Delayed Mineralization | ||
| R2 | 0.82 | 0.79 | 0.75 | 0.74 | 0.86 |
| RMSE | 1487 | 1666 | 1664 | 1680 | 1299 |
| NSE | 0.48 | 0.34 | 0.35 | 0.33 | 0.60 |
| Pbias | 7.90 | 9.50 | 8.90 | 8.90 | 6.60 |
| Generic Approach | Generic Approach | SMM | SMM | Without a Ryegrass Period | |
|---|---|---|---|---|---|
| Aboveground | Above- and Belowground | Immediate Mineralization | Delayed Mineralization | ||
| R2 | 0.25 | 0.49 | 0.56 | 0.06 | 0.44 |
| RMSE | 37 | 29 | 27 | 43 | 35 |
| NSE | −0.17 | 0.31 | 0.38 | −0.53 | −0.02 |
| Pbias | −12.80 | −5.00 | −4.40 | −11.30 | −13.00 |
| Generic Approach | Generic Approach | SMM | SMM | Without a Ryegrass Period | |
|---|---|---|---|---|---|
| Aboveground | Above- and Belowground | Immediate Mineralization | Delayed Mineralization | ||
| R2 | 0.34 | 0.09 | 0.03 | 0.37 | 0.00 |
| RMSE | 21 | 24 | 26 | 21 | 25 |
| NSE | 0.08 | −0.14 | −0.38 | 0.13 | −0.27 |
| Pbias | −35.60 | −21.80 | −17.20 | −33.80 | −14.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alderkamp, L.M.; Vogeler, I.; Poyda, A.; Manevski, K.; van Middelaar, C.E.; Taube, F. Yields and Nitrogen Dynamics in Ley-Arable Systems—Comparing Different Approaches in the APSIM Model. Agronomy 2022, 12, 738. https://doi.org/10.3390/agronomy12030738
Alderkamp LM, Vogeler I, Poyda A, Manevski K, van Middelaar CE, Taube F. Yields and Nitrogen Dynamics in Ley-Arable Systems—Comparing Different Approaches in the APSIM Model. Agronomy. 2022; 12(3):738. https://doi.org/10.3390/agronomy12030738
Chicago/Turabian StyleAlderkamp, Lianne M., Iris Vogeler, Arne Poyda, Kiril Manevski, Corina E. van Middelaar, and Friedhelm Taube. 2022. "Yields and Nitrogen Dynamics in Ley-Arable Systems—Comparing Different Approaches in the APSIM Model" Agronomy 12, no. 3: 738. https://doi.org/10.3390/agronomy12030738