
Phenology and Population Differentiation in Reproductive Plasticity in Feathertop Rhodes Grass (Chloris virgata Sw.)



Abstract
:1. Introduction
- Is there any impact of different emergence dates on the growth and development timing of phenological, morphological and reproductive traits of feathertop Rhodes grass?
- Does timing and duration of each phenological events vary among populations of feathertop Rhodes grass?
2. Materials and Methods
2.1. Environmental (Emergence Timing) and Populations Treatment
2.2. Experiment Set Up and Measurements
2.3. Design and Data Analysis
3. Results
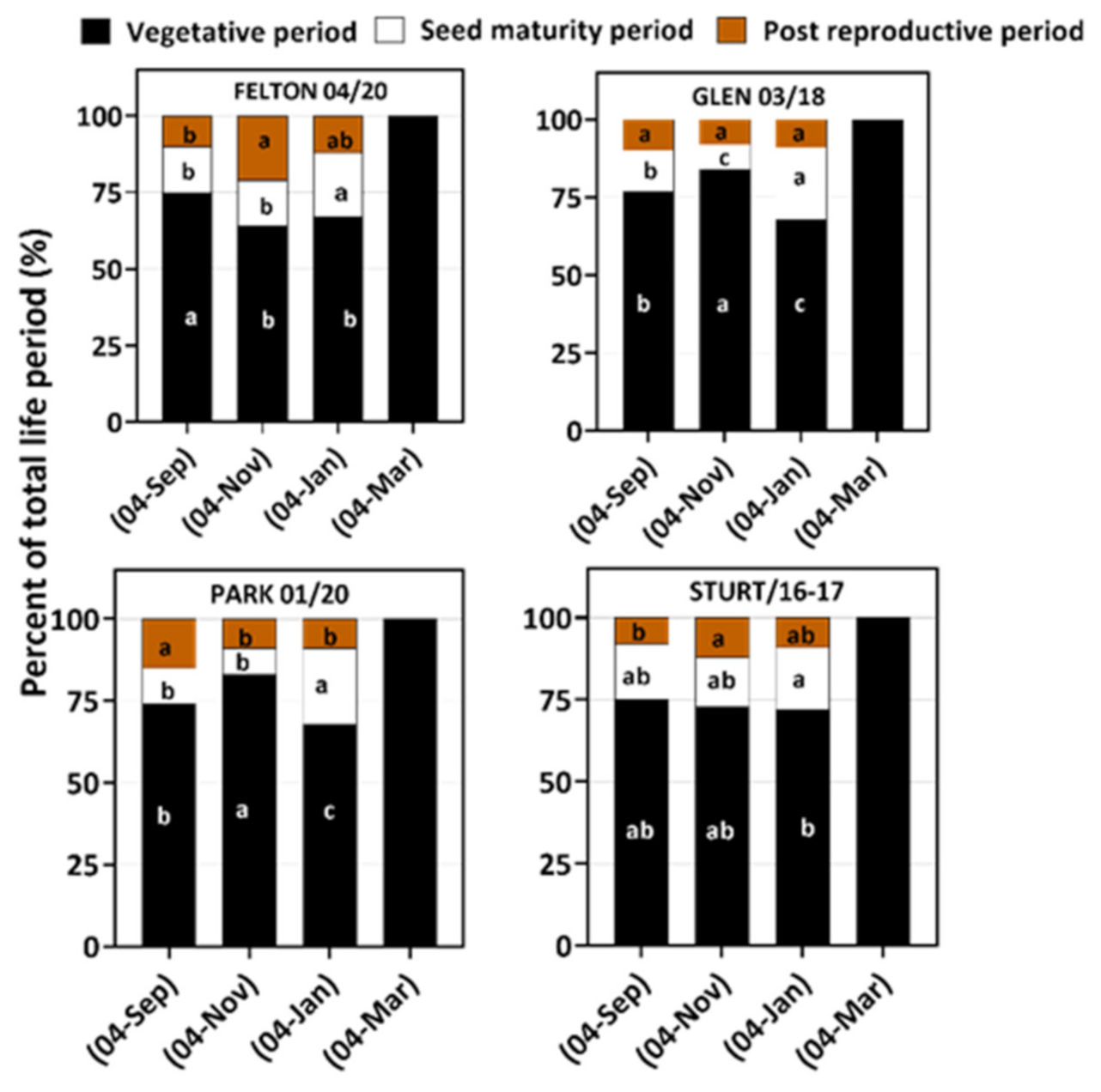
3.1. Phenophase of Four Populations
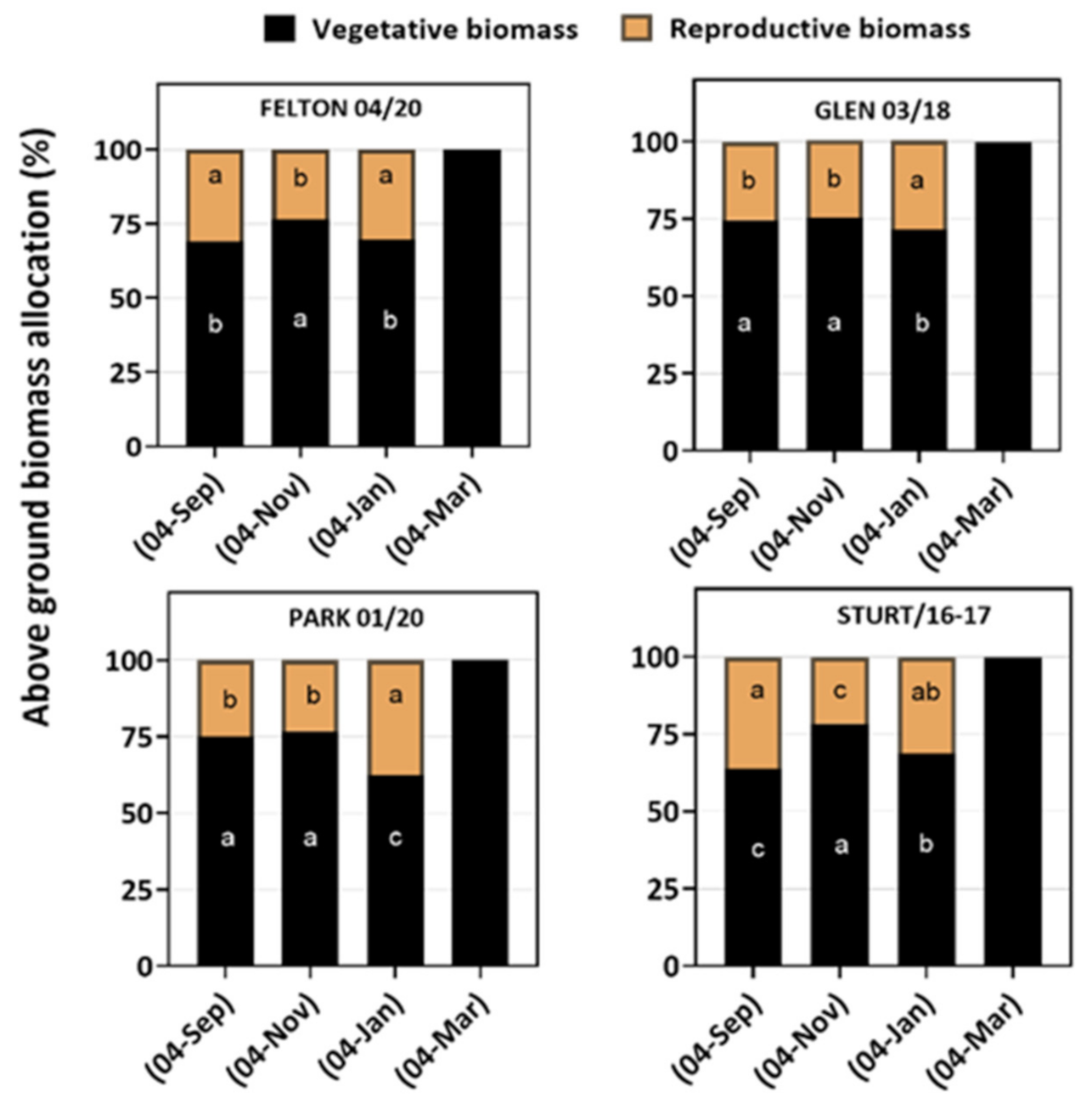
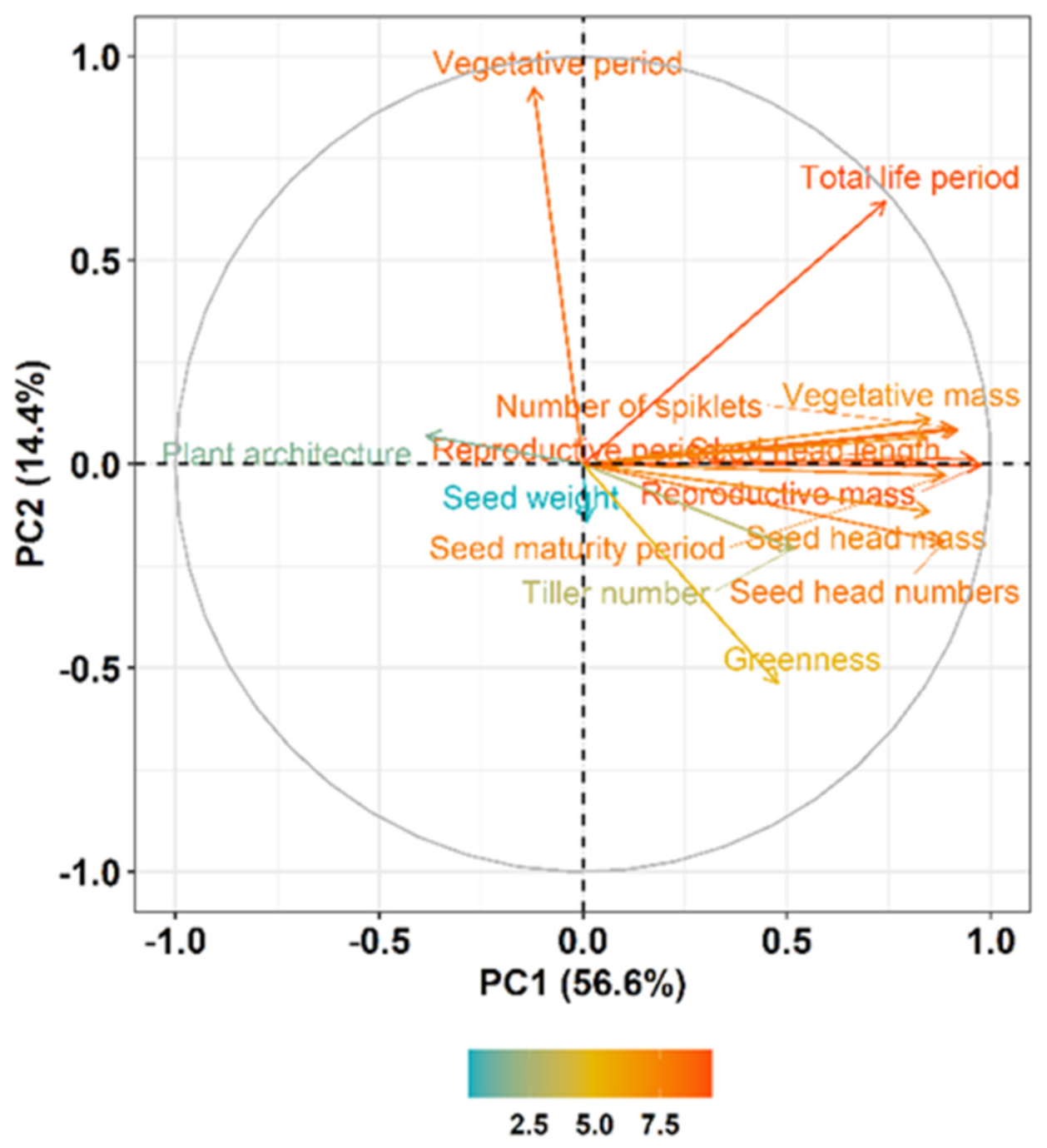
3.2. Plasticity in Plant Growth and Biomass Allocation
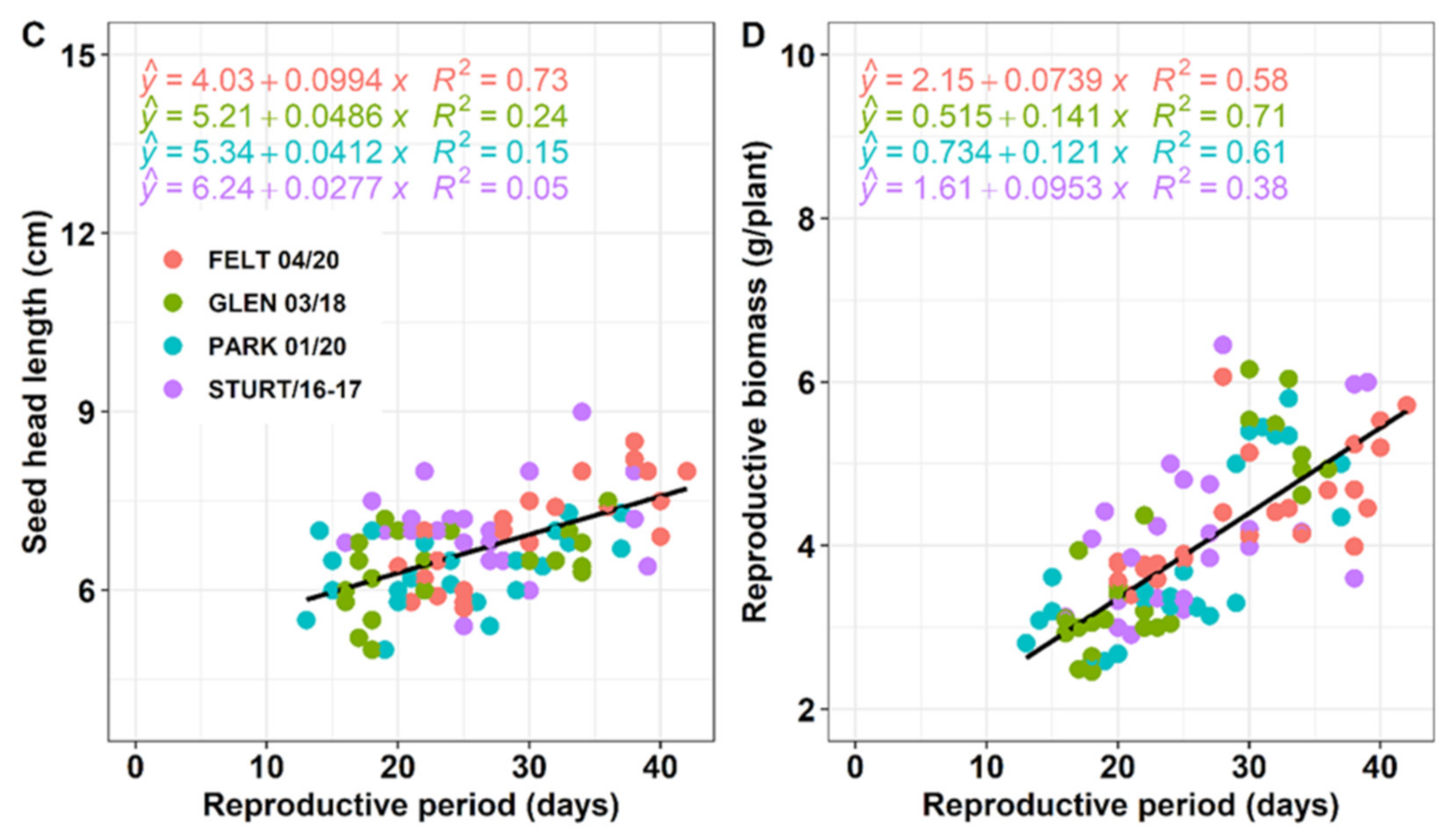
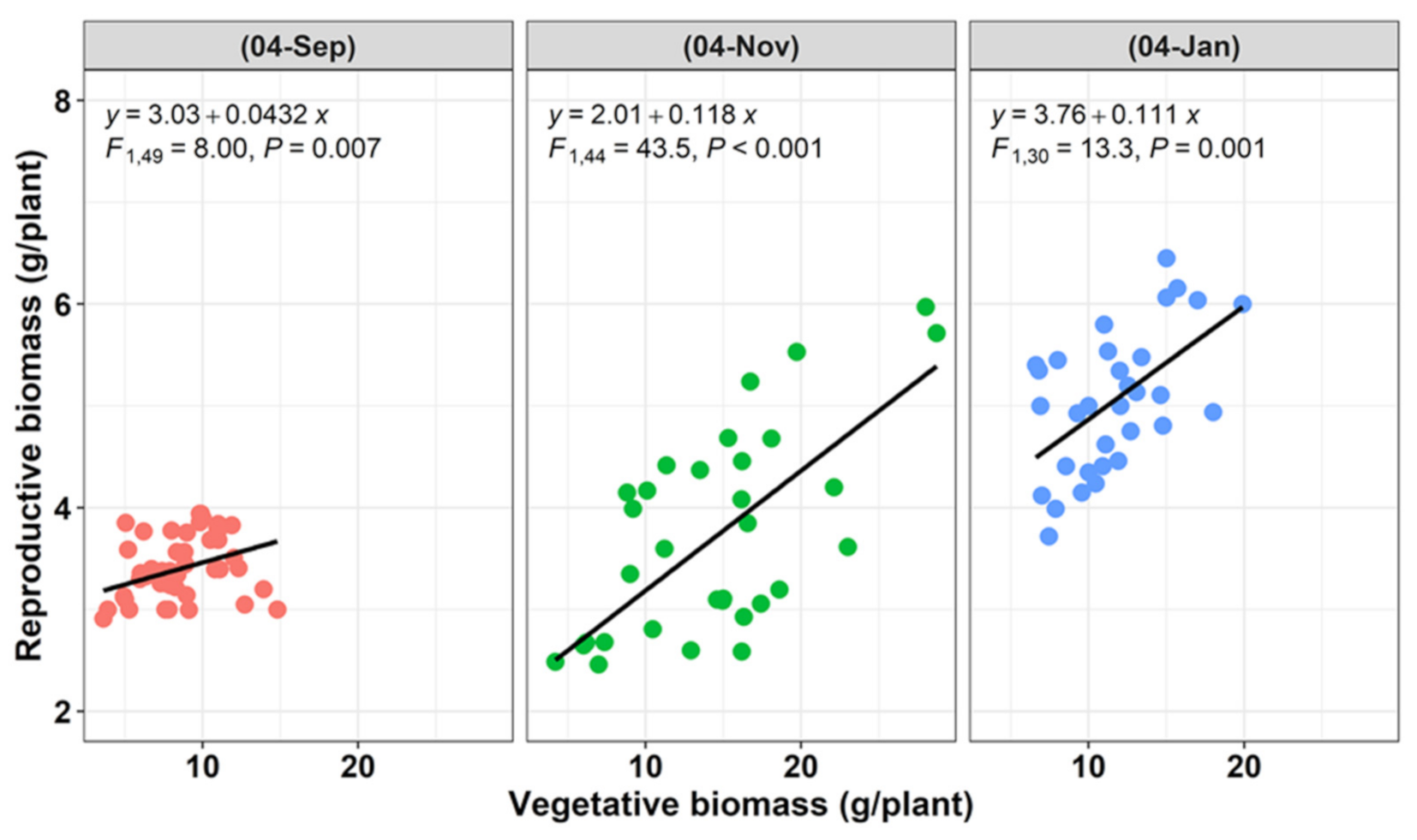
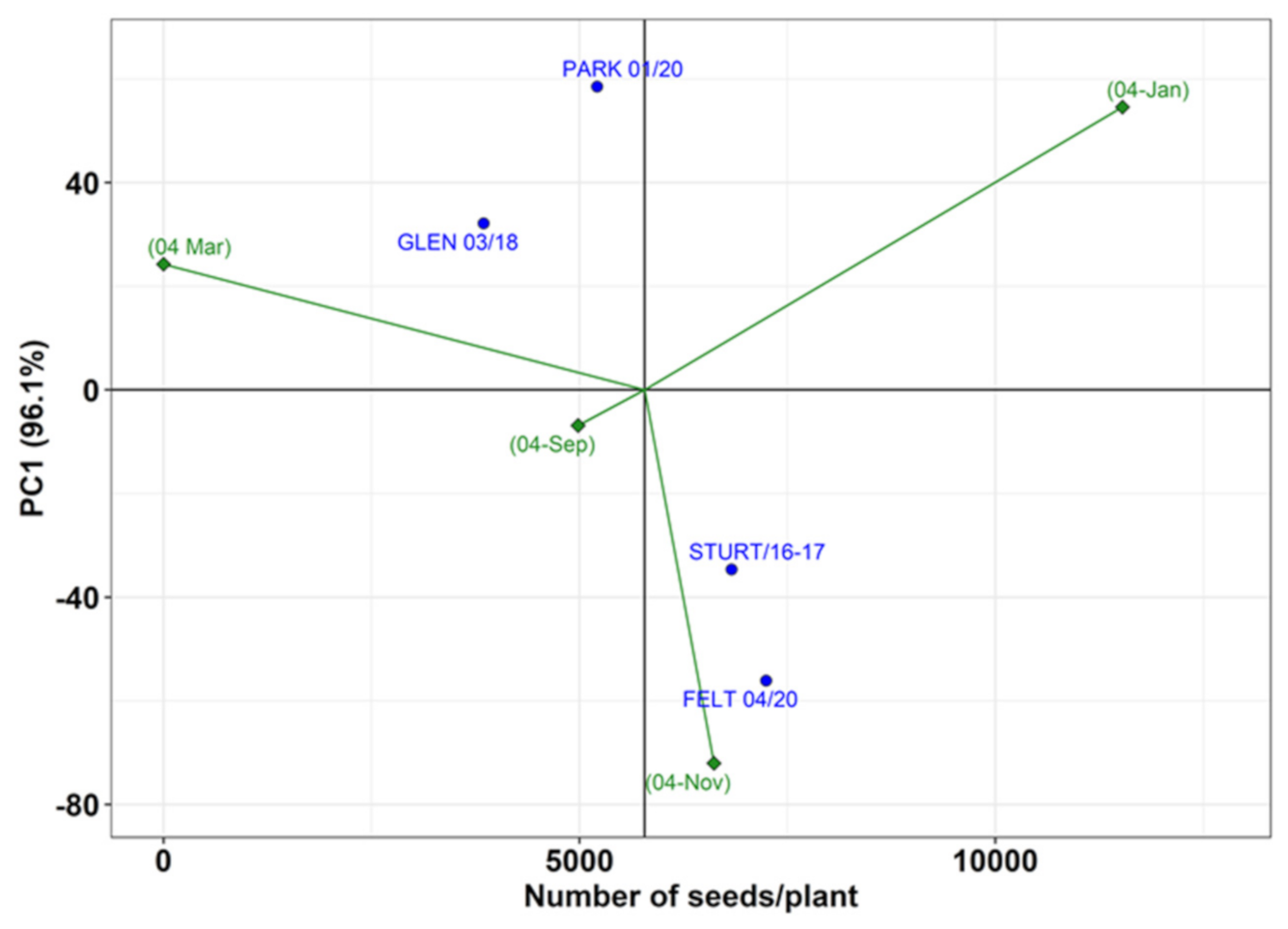
3.3. Plasticity in Reproductive Effort
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Schwartz, M.D. Phenology: An Integrated Environmental Science, 3rd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 275–293. [Google Scholar]
- Godoy, O.; Richardson, D.; Valladares, F.; Castro-Diez, P. Flowering phenology of invasive alien plant species compared with native species in three Mediterranean-type ecosystems. Ann. Bot. 2008, 103, 485–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.A.; Evans, R.A.; Kay, B. Phenology of Reproduction of Medusa Head. Weed Sci. 1970, 18, 451–454. [Google Scholar] [CrossRef]
- Ghersa, C.M.; Holt, J.S. Using phenology prediction in weed management: A review. Weed Res. 1995, 35, 461–470. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Bahadur, H.; Chand, B.; Kumar, B. Phenological documentation of Lantana camara L. using modified BBCH scale in relation to climatic variables. Plant Sci. Today 2022, 1481, 1–10. [Google Scholar] [CrossRef]
- Bryson, C.T.; Carter, R. Biology of Pathways for Invasive Weeds1. Weed Technol. 2004, 18, 1216–1220. [Google Scholar] [CrossRef]
- MacLaren, C.; Storkey, J.; Menegat, A.; Metcalfe, H.; Dehnen-Schmutz, K. An ecological future for weed science to sustain crop production and the environment. A review. Agron. Sustain. Dev. 2020, 40, 1–29. [Google Scholar] [CrossRef]
- Dinçer, D.; Var, M.; Baykal, H.; Atamov, V. Phenological features of some geophytes from the Anzer plateau in Rize and utilization possibilities for landscape architecture. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC2014), Brisbane, Australia, 17–22 August 2014; Volume 1108, pp. 187–194. [Google Scholar]
- Rafferty, N.E.; Ives, A.R. Effects of experimental shifts in flowering phenology on plant-pollinator interactions. Ecol. Lett. 2010, 14, 69–74. [Google Scholar] [CrossRef]
- Sultan, E.E. Evolutionary implications of phenotypic plasticity in plants. In Evolutionary Biology; Hecht, M., Wallace, B., Prance, G., Eds.; Plenum Press: New York, NY, USA, 1987; pp. 127–178. [Google Scholar]
- Alpert, P.; Simms, E.L. The relative advantages of plasticity and fixity in different environments: When is it good for a plant to adjust? Evol. Ecol. 2002, 16, 285–297. [Google Scholar] [CrossRef]
- Bradshaw, W.E.; Holzapfel, C.M. Evolutionary Response to Rapid Climate Change. Science 2006, 312, 1477–1478. [Google Scholar] [CrossRef] [Green Version]
- Clements, D.R.; Jones, V.L. Rapid Evolution of Invasive Weeds Under Climate Change: Present Evidence and Future Research Needs. Front. Agron. 2021, 3, 10. [Google Scholar] [CrossRef]
- Schlichting, C.D. The evolution of phenotypic plasticity in plants. Ann. Rev. Ecol. Syst. 1986, 17, 667–693. [Google Scholar] [CrossRef]
- Sans, F.X.; Masalles, R.M. Demography of the arable weed Diplotaxis erucoides in central Catalonia, Spain. Can. J. Bot. 1997, 75, 86–95. [Google Scholar] [CrossRef]
- Donohue, K.; De Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, Postgermination Adaptation, and Species Ecological Ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Carter, S.K.; Saenz, D.; Rudolf, V.H.W. Shifts in phenological distributions reshape interaction potential in natural communities. Ecol. Lett. 2018, 21, 1143–1151. [Google Scholar] [CrossRef]
- Anderson, D.E. Taxonomy of the Genus Chloris (Gramineae). Brigh. Young Univ. Sci. Bull. 1974, 2, 133. [Google Scholar]
- Werth, J.; Boucher, L.; Thornby, D.; Walker, S.; Charles, G. Changes in weed species since the introduction of glyphosate-resistant cotton. Crop Pasture Sci. 2013, 64, 791–798. [Google Scholar] [CrossRef]
- Llewellyn, R.; Ronning, D.; Ouzman, J.; Walker, S.; Mayfield, A.; Clarke, M. The cost of weeds to Australian grain growers and the adoption of weed management and tillage practices. In Impact of Weeds on Australian Grain Production; GRDC: Kingston, Australia, 2016; Volume 112. [Google Scholar]
- Atlas of Living Australia. Available online: https://ala.org.au/ (accessed on 10 March 2022).
- Weiner, J.; Campbell, L.G.; Pino, J.; Echarte, L. The allometry of reproduction within plant populations. J. Ecol. 2009, 97, 1220–1233. [Google Scholar] [CrossRef]
- Manuel, C.; Molles, J. Ecology: Concepts and Applicants, 2nd ed.; Higher Education Press: Beijing, China, 2002; pp. 279–299. [Google Scholar]
- Forcella, F.; Arnold, R.L.B.; Sanchez, R.; Ghersa, C.M. Modeling seedling emergence. Field Crop. Res. 2000, 67, 123–139. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, T.; Valentine, I. Phenotypic plasticity of life-history characters in response to different germination timing in two annual weeds. Can. J. Bot. 2005, 83, 28–36. [Google Scholar] [CrossRef]
- Li, H.; Lindquist, J.L.; Yang, Y. Effects of Sowing Date on Phenotypic Plasticity of Fitness-Related Traits in Two Annual Weeds on the Songnen Plain of China. PLoS ONE 2015, 10, e0127795. [Google Scholar] [CrossRef] [Green Version]
- Yamasue, Y.; Murayama, H.; Inoue, H.; Matsui, T.; Kusanagi, T. Productive structures of rice and Echinochloa phyllopogon Vasing. in mixed stands. J. Weed Sci. Technol. 1997, 42, 57–365. [Google Scholar]
- Clements, D.R.; DiTommaso, A.; Jordan, N.; Booth, B.D.; Cardina, J.; Doohan, D.; Mohler, C.L.; Murphy, S.D.; Swanton, C.J. Adaptability of plants invading North American cropland. Agric. Ecosyst. Environ. 2004, 104, 379–398. [Google Scholar] [CrossRef]
- Moran, E.V.; Alexander, J. Evolutionary responses to global change: Lessons from invasive species. Ecol. Lett. 2014, 17, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Andersson, S.; Shaw, R.G. Phenotypic plasticity in Crepis tectorum: Genetic correlations across light regimes. Heredity 1994, 72, 113–125. [Google Scholar] [CrossRef]
- Kaweck, T.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Gould, B.; Moeller, D.A.; Eckhart, V.M.; Tiffin, P.; Fabio, E.; Geber, M.A. Local adaptation and range boundary formation in response to complex environmental gradients across the geographical range of Clarkia xantiana ssp. xantiana. J. Ecol. 2014, 102, 95–107. [Google Scholar] [CrossRef]
- Schlichting, C.D.; Smith, H. Phenotypic plasticity: Linking molecular mechanisms with evolutionary outcomes. Evol. Ecol. 2002, 16, 189–211. [Google Scholar] [CrossRef]
- Weiner, J.; Rosenmeier, L.; Massoni, E.S.; Vera, J.N.; Hernández Plaza, E.; Sebastia, M.T. Is reproductive allocation in Senecio vulgaris plastic? Botany 2009, 87, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Aarssen, L.W.; Taylor, D.R. Fecundity Allocation in Herbaceous Plants. Oikos 1992, 65, 225. [Google Scholar] [CrossRef]
- Grundy, A.C.; Mead, A.; Burston, S.; Overs, T. Seed production of Chenopodium album in competition with field vegetables. Weed Res. 2004, 44, 271–281. [Google Scholar] [CrossRef]
- Neytcheva, M.S.; Aarssen, L.W. More plant biomass results in more offspring production in annuals, or does it? Oikos 2008, 117, 1298–1307. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Morse, S.R. Annual plants potential responses to multiple stresses. In Response of Plants to Multiple Stresses; Mooney, H.A., Winner, W.E., Pell, E.J., Chu, E., Eds.; Academic Press: San Diego, CA, USA, 1991; Volume 1, pp. 283–305. [Google Scholar]
- Knezevic, S.; Evans, S.; Blankenship, E.; Van Acker, R.; Lindquist, J. Critical period for weed control: The concept and data analysis. Weed Sci. 2017, 50, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Knezevic, S.Z.; Weise, S.F.; Swanton, C.J. Interference of redroot pigweed (Amaranthus retroflexus L.) in corn (Zea mays L.). Weed Sci. 1994, 42, 568–573. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Geographic Location | Habitat | Ecotype |
|---|---|---|---|
| FELT 04/20 | Felton, QLD | Roadside | Northern |
| GLEN 03/18 | Toowoomba, QLD | Cropping | Northern |
| STURT 16–17 | Wagga Wagga, NSW | Roadside | Southern |
| PARK 01/20 | Parkes, NSW | Cropping | Southern |
| Month | Temperature (°C) | Rainfall (mm) | Solar Exposure (MJ m−2) |
|---|---|---|---|
| January | 24.15 | 43.4 | 27.3 |
| February | 23.65 | 53.8 | 24.0 |
| March | 20.6 | 51.2 | 19.9 |
| April | 15.9 | 29.4 | 14.4 |
| May | 11.65 | 36.6 | 10.3 |
| June | 8.8 | 52.1 | 7.7 |
| July | 7.8 | 46.0 | 8.4 |
| August | 9.0 | 42.6 | 11.6 |
| September | 11.40 | 42.0 | 16.1 |
| October | 14.75 | 40.7 | 21.4 |
| November | 18.45 | 62.3 | 26.6 |
| December | 21.75 | 51.5 | 27.3 |
| Month | Min. Temperature (°C) | Max. Temperature (°C) | Rainfall (mm) | Solar Exposure (MJ m−2) |
|---|---|---|---|---|
| September/2020 | 5.6 | 23.4 | 57.4 | 16.0 |
| October/2020 | 8.8 | 28.2 | 85.0 | 18.4 |
| November/2020 | 11.6 | 36.3 | 60.0 | 25.7 |
| December/2020 | 12.1 | 34.9 | 78.2 | 26.6 |
| January/2021 | 14.9 | 40.1 | 77.4 | 25.0 |
| February/2021 | 14.1 | 40.03 | 78.2 | 23.8 |
| March/2021 | 12.2 | 33.4 | 112.2 | 16.6 |
| April/2021 | 6.5 | 31.7 | 2.4 | 14.7 |
| May/2021 | 5.0 | 25.5 | 32.2 | 10.4 |
| June/2021 | 3.8 | 19.1 | 74.2 | 7.7 |
| July/2021 | 3.9 | 13.8 | 59.2 | 8.5 |
| Sowing Time | Population | Emergence Date | Emergence to First Seed Head Emerging (Days) | Vegetative Period | Seed Maturity Period | Post-Reproductive Period | Reproductive Period | Life Period |
|---|---|---|---|---|---|---|---|---|
| 4 September | FELT 04/20 | 7 September | 69–71 | 64. 00 (±0.28) | 13.07 (±0.65) | 9.38 (±0.89) | 22.45 (±0.70) | 86.45 (±0.65) |
| STURT16–17 | 8 September | 63–73 | 63.87 (±0.85) | 14.50 (±1.11) | 6.60 (±0.50) | 21.10 (±0.93) | 84.97 (±1.34) | |
| PARK 01/20 | 8 September | 72–80 | 69.00 (±1.17) | 10.00 (±1.30) | 14.71 (±1.11) | 24.28 (±0.92) | 93.75 (±1.5) | |
| GLEN 03/18 | 9 September | 85–95 | 65.27 (±4.90) | 11.37 (±0.77) | 9.00 (±0.54) | 20.37 (±0.92) | 85.64 (±5.4) | |
| 4 November | FELT 04/20 | 7 November | 70–72 | 68.10 (±2.00) | 15.28 (±0.60) | 23.27 (±1.78) | 39.07 (±0.85) | 107.17 (±2.34) |
| STURT16–17 | 8 November | 85–95 | 77.07 (±4.00) | 16.00 (±2.15) | 12.72 (±1.05) | 28.72 (±2.76) | 105.79 (±5.00) | |
| PARK 01/20 | 8 November | 95–100 | 69.00 (±0.75) | 8.25 (±0.80) | 8.54 (±0.70) | 16.79 (±0.99) | 85.79 (±3.86) | |
| GLEN 03/18 | 10 November | 100–110 | 90.00 (±2.26) | 8.78 (±0.65) | 8.89 (±0.84) | 17.67 (±0.71) | 107.67 (±3.28) | |
| 4 January | FELT 04/20 | 6 January | 65–70 | 61.50 (±1.13) | 19.75 (±1.30) | 10.63 (±0.74) | 30.38 (±1.81) | 91.80 (±1.61) |
| STURT16–17 | 6 January | 63–75 | 70.75 (±4.20) | 18.75 (±1.03) | 9.12 (±0.93) | 27.87 (±1.77) | 98.62 (±0.73) | |
| PARK 01/20 | 7 January | 72–78 | 83.18 (±3.03) | 23.75 (±1.04) | 9.00 (±0.46) | 32.75 (±1.04) | 115.93 (±1.07) | |
| GLEN 03/18 | 7 January | 78–85 | 70.37 (±1.50) | 23.00 (±0.92) | 9.25 (±0.59) | 32.25 (±1.04) | 102.62 (±1.64) | |
| 4 March | FELT 04/20 | 7 March | 67–80 | 69.57 (±1.34) | - | - | - | - |
| STURT16–17 | 7 March | 67–81 | 64.15 (±0.48) | - | - | - | - | |
| PARK 01/20 | 8 March | 80–90 | 71.25 (±1.11) | - | - | - | - | |
| GLEN 03/18 | 8 March | 80–90 | 79.26 (±1.63) | - | - | - | - | |
| - | p-value | - | p = 0.003 | p = 0.004 | p = 0.003 | p = 0.002 | p = 0.002 | p = 0.002 |
| Sowing Time | Population | Tillers/Plant | Plant Architecture | Number of Seed Heads/Plant | Seed Head Length (cm) | Seed Head Biomass (g) | Number of Spikelets/Head | Seeds/Plant | Slope (R-V) |
|---|---|---|---|---|---|---|---|---|---|
| 4 September | FELT 04/20 | 13 (±1.0) | Semi-erect | 9.60 (±0.90) | 6.09 (±0.77) | 1.22 (±0.03) | 10.53 (±0.21) | 6521 (±770) | 0.045c |
| STURT16-17 | 14 (±1.6) | Semi-erect | 9.46 (±0.22) | 6.72 (±0.25) | 0.74 (±0.08) | 9.46 (±0.25) | 5951 (±390) | 0.063 b | |
| PARK 01/20 | 15 (±1.3) | Semi-erect | 6.20 (±0.41) | 6.15 (±0.17) | 0.98 (±0.05) | 10.14 (±0.18) | 4390 (±415) | 0.061 b | |
| GLEN 03/18 | 17 (±1.0) | Semi-prostrate | 4.45 (±1.40) | 6.75 (±0.11) | 0.82 (±0.14) | 8.80 (±0.61) | 1969 (±600) | 0.008 c | |
| 4 November | FELT 04/20 | 13 (±1.3) | Erect | 17.90 (±1.88) | 8.00 (±0.28) | 2.42 (±0.20) | 10.63 (±0.33) | 11112 (±1575) | 0.085 b |
| STURT16-17 | 22 (±2.8) | Erect | 18.18 (±2.28) | 7.20 (±0.28) | 2.03 (±0.26) | 9.36 (±0.21) | 9875 (±1362) | 0.094 b | |
| PARK 01/20 | 24 (±1.8) | Erect | 3.50 (±0.55) | 6.05 (±0.11) | 1.10 (±0.45) | 10.36 (±0.20) | 8071 (±446) | 0.042 c | |
| GLEN 03/18 | 12 (±1.0) | Semi-prostrate | 4.50 (±1.01) | 7.10 (±0.08) | 1.14 (±0.53) | 8.92 (±0.14) | 2937 (±1400) | 0.059 c | |
| 4 January | FELT 04/20 | 13 (±1.3) | Erect | 12.12 (±1.50) | 6.90 (±0.18) | 2.19 (±0.14) | 13.00 (±0.50) | 9930 (±1232) | 0.233 a |
| STURT16-17 | 13 (±1.7) | Erect | 14.37 (±1.80) | 7.00 (±0.18) | 2.64 (±0.32) | 12.00 (±0.52) | 10867 (±1362) | 0.194 a | |
| PARK 01/20 | 13 (±1.4) | Semi-prostrate | 15.00 (±1.01) | 6.68 (±0.18) | 3.56 (±0.35) | 13.00 (±0.35) | 16687 (±670) | 0.002 c | |
| GLEN 03/18 | 12 (±1.3) | Erect | 12.00 (±2.0) | 6.30 (±0.16) | 2.98 (±0.18) | 13.00 (±0.908) | 12285 (±579) | 0.078 b | |
| 4 March | FELT 04/20 | 09 (±1.0) | Semi-prostrate | - | - | - | - | - | - |
| STURT16-17 | 13 (±1.3) | Semi-prostrate | - | - | - | - | - | - | |
| PARK 01/20 | 14 (±1.2) | Semi-prostrate | - | - | - | - | - | - | |
| GLEN 03/18 | 11 (±11) | Prostrate | - | - | - | - | - | - | |
| p-value | p = 0.004 | p = 0.0001 | p = 0.0002 | p = 0.0002 | p = 0.0001 | p = 0.0002 | p = 0.0003 | p = 0.0005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asaduzzaman, M.; Wu, H.; Koetz, E.; Hopwood, M.; Shepherd, A. Phenology and Population Differentiation in Reproductive Plasticity in Feathertop Rhodes Grass (Chloris virgata Sw.). Agronomy 2022, 12, 736. https://doi.org/10.3390/agronomy12030736
Asaduzzaman M, Wu H, Koetz E, Hopwood M, Shepherd A. Phenology and Population Differentiation in Reproductive Plasticity in Feathertop Rhodes Grass (Chloris virgata Sw.). Agronomy. 2022; 12(3):736. https://doi.org/10.3390/agronomy12030736
Chicago/Turabian StyleAsaduzzaman, Md, Hanwen Wu, Eric Koetz, Michael Hopwood, and Adam Shepherd. 2022. "Phenology and Population Differentiation in Reproductive Plasticity in Feathertop Rhodes Grass (Chloris virgata Sw.)" Agronomy 12, no. 3: 736. https://doi.org/10.3390/agronomy12030736