
Meloidogyne graminicola’s Effect on Growth Performance of Rice under Low Population Density



Abstract
:1. Introduction
2. Materials and Methods
2.1. Rice Seedling Nursery
2.2. Nematode Inoculum
2.3. Experimental Design
2.4. Statistical Analyses
3. Results
3.1. Effect of Inoculum Density on Rice Plant Growth
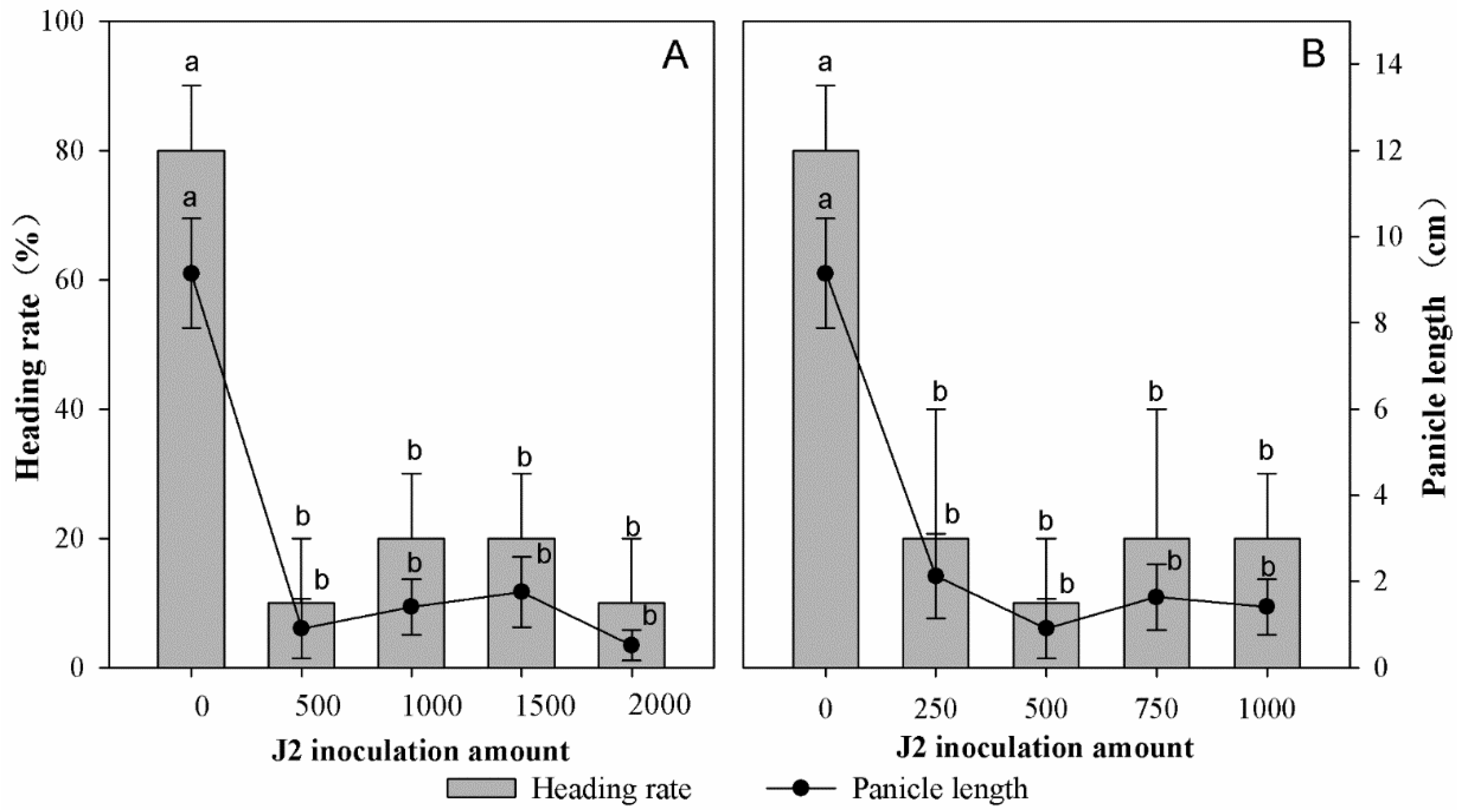
3.2. Effect of Inoculum Density on Rice Panicle Development
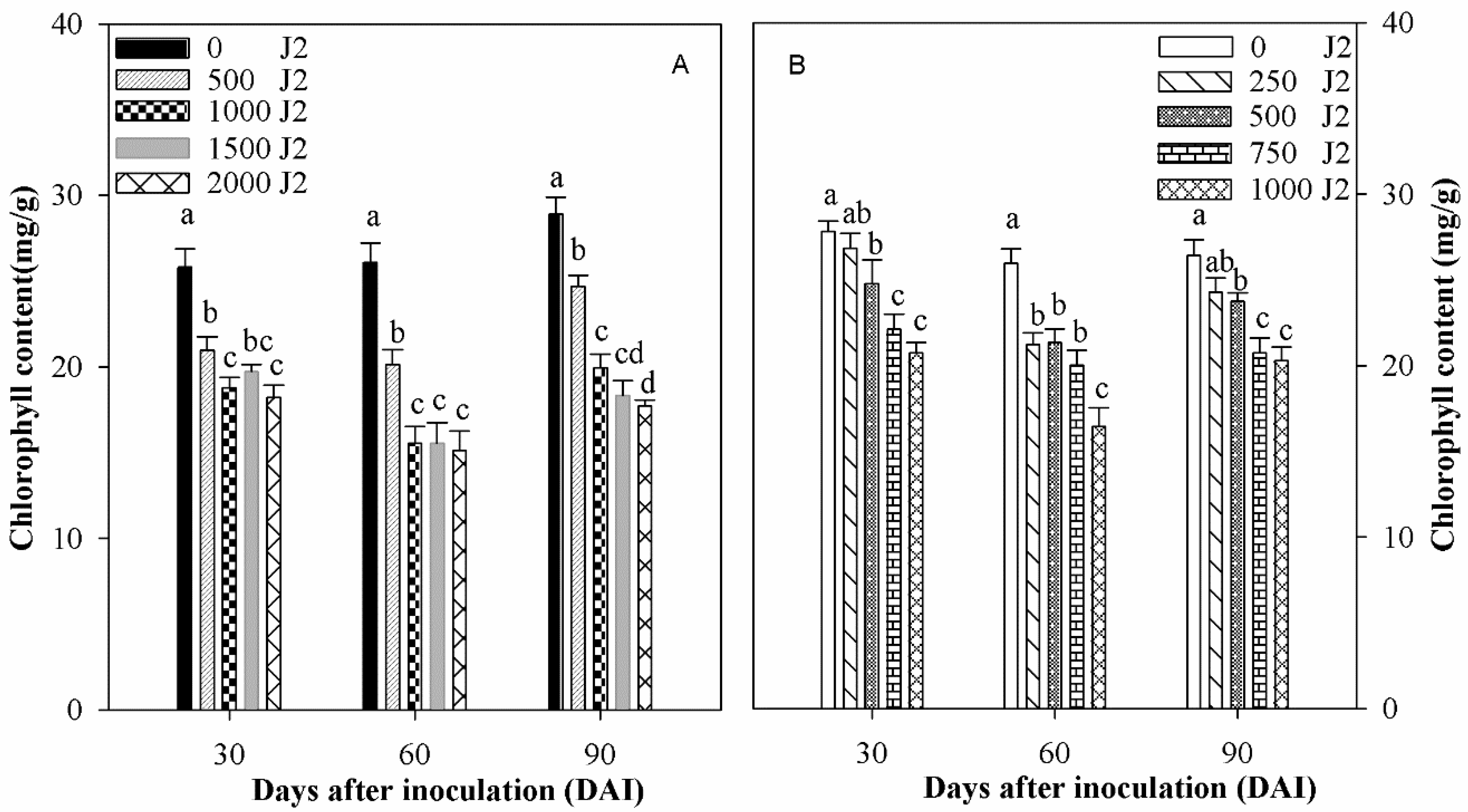
3.3. Effect of Inoculum Density on Chlorophyll Content in Rice
3.4. Effect of Inoculum Density on Photosynthetic Parameters of Rice
3.4.1. Effects of Inoculum Density on Transpiration Rate and Net Photosynthetic Rate of Rice
3.4.2. Effects of Inoculum Density on Total Conductance of CO2 and Water of Rice
3.4.3. Effects of Inoculum Density on Partial Pressure of Intercellular CO2 and Intracellular CO2 Concentration of Rice
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. FAO Statistics. Available online: http://www.fao.org/faostat/zh/#data/QCL/visualize (accessed on 18 March 2021).
- Jain, R.K.; Khan, M.R.; Kumar, V. Rice root-knot nematode (Meloidogyne graminicola) infestation in rice. Arch. Phytopathol. Plant Prot. 2012, 45, 635–645. [Google Scholar] [CrossRef]
- Besnard, G.; Thi-Phan, N.; Ho-Bich, H.; Dereeper, A.; Nguyen, H.T.; Queneherve, P.; Aribi, J.; Bellafiore, S. On the close relatedness of two rice-parasitic root-knot nematode species and the recent expansion of Meloidogyne graminicola in Southeast Asia. Genes 2019, 10, 175. [Google Scholar] [CrossRef] [Green Version]
- Mantelin, S.; Bellafiore, S.; Kyndt, T. Meloidogyne graminicola: A major threat to rice agriculture. Mol. Plant Pathol. 2017, 18, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Ruanpanun, P.; Khun-In, A. First report of Meloidogyne incognita caused root knot disease of upland rice in Thailand. J. Int. Soc. Southeast Asian Agric. Sci. 2015, 21, 68–77. [Google Scholar]
- Negretti, R.R.R.D.; Gomes, C.B.; Mattos, V.S.; Somavilla, L.; Manica-Berto, R.; Agostinetto, D.; Castagnone-Sereno, P.; Carneiro, R.M.D.G. Characterisation of a Meloidogyne species complex parasitising rice in southern Brazil. Nematology 2017, 19, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.R.; Haque, Z.; Kausar, N. Management of the root-knot nematode Meloidogyne graminicola infesting rice in the nursery and crop field by integrating seed priming and soil application treatments of pesticides. Crops Prot. 2014, 63, 15–25. [Google Scholar] [CrossRef]
- Xie, J.; Fang, Y.; Huang, W.; Peng, D.; Peng, Y.; Ji, H. Advances in major rice parasitic nematodes in recent years. J. Plant Prot. 2017, 44, 940–949. [Google Scholar]
- Baojun, Y. A new species of root—knot nematode—Meloidogyne lini n. sp. parasitizing rice. J. Yunnan Agric. Univ. 1988, 1, 11–17. [Google Scholar]
- Liao, J.; Feng, Z. A new species of the genus Meloidogyne—Meloidogyne hainanensis. J. South China Agric. Univ. 1995, 3, 34–39. [Google Scholar]
- Zhao, H.; Liu, W.; Chen, L.; Yu, D. Meloidogyne graminicola, a new record species from China. Acta Phytopathol. Sin. 2001, 31, 184–188. [Google Scholar]
- Liu, G.; Xiao, S.; Zhang, S.; Zhang, D.; Wang, Y. Infection characteristic and life cycle of rice root-knot nematode, Meloidogyne graminicola in rice root. Chin. J. Trop. Crops 2011, 32, 743–748. [Google Scholar]
- Tang, B.; Wang, D.; Wang, J.; He, Q.; Liu, Y.; Zhang, D.; Cheng, F. Effects of different planting modes on occurrence of rice root-knot nematode disease. Plant Prot. 2021, 47, 188–191. [Google Scholar]
- Tian, Z.L.; Barsalote, E.M.; Li, X.L.; Cai, R.H.; Zheng, J.W. First report of root-knot nematode, Meloidogyne graminicola, on rice in Zhejiang, Eastern China. Plant Dis. 2017, 101, 2152–2153. [Google Scholar]
- Wang, G.F.; Xiao, L.Y.; Luo, H.G.; Peng, D.L.; Xiao, Y.N. First report of Meloidogyne graminicola on rice in Hubei Province of China. Plant Dis. 2017, 101, 1056–1057. [Google Scholar] [CrossRef]
- Song, Z.Q.; Zhang, D.Y.; Liu, Y.; Cheng, F.X. First report of Meloidogyne graminicola on rice (Oryza sativa) in Hunan Province, China. Plant Dis. 2017, 101, 2153. [Google Scholar] [CrossRef]
- Xie, J.L.; Xu, X.; Yang, F.; Xue, Q.; Peng, Y.L.; Ji, H.L. First report of root-knot nematode, Meloidogyne graminicola, on rice in Sichuan Province, Southwest China. Plant Dis. 2019, 103, 2142. [Google Scholar]
- Ju, Y.L.; Wu, X.; Tan, G.J.; Peng, D.L.; Xu, J.F.; Qiu, K.; Wu, H.P. First report of Meloidogyne graminicola on rice in Anhui Province, China. Plant Dis. 2021, 105, 512. [Google Scholar] [CrossRef]
- Liu, M.Y.; Liu, J.; Huang, W.; Peng, D. First report of Meloidogyne graminicola on rice in Henan Province, China. Plant Dis. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Luo, M.; Li, B.X.; Wu, H.Y. Incidence of the rice root-knot nematode, Meloidogyne graminicola, in Guangxi, China. Plant Pathol. J. 2020, 36, 384. [Google Scholar] [CrossRef]
- Huang, W.; Xiang, C.; Liu, Y.; Ding, Z.; Peng, D. Rearch progress on the occurrence and controlling of root-knot nematode Meloidogyne graminicola in rice. Acta Phytopathol. Sin. 2018, 48, 289–296. [Google Scholar]
- Eickhorst, T.; Tippkotter, R. Management-induced structural dynamics in paddy soils of south east China simulated in microcosms. Soil Tillage Res. 2009, 102, 168–178. [Google Scholar] [CrossRef]
- Abad, P.; Favery, B.; Rosso, M.N.; Castagnone-Sereno, P. Root-knot nematode parasitism and host response: Molecular basis of a sophisticated interaction. Mol. Plant Pathol. 2003, 4, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Strajnar, P.; Sirca, S.; Urek, G.; Sircelj, H.; Zeleznik, P.; Vodnik, D. Effect of Meloidogyne ethiopica parasitism on water management and physiological stress in tomato. Eur. J. Plant Pathol. 2012, 132, 49–57. [Google Scholar] [CrossRef]
- Kyndt, T.; Fernandez, D.; Gheysen, G. Plant-parasitic nematode infections in rice: Molecular and cellular insights. Annu. Rev. Phytopathol. 2014, 52, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Soriano, I.R.; Reversat, G. Management of Meloidogyne graminicola and yield of upland rice in South-Luzon, Philippines. Nematology 2003, 5, 879–884. [Google Scholar] [CrossRef]
- Kumari, C.; Dutta, T.K.; Banakar, P.; Rao, U. Comparing the defence-related gene expression changes upon root-knot nematode attack in susceptible versus resistant cultivars of rice. Sci. Rep. 2016, 6, 22846. [Google Scholar] [CrossRef] [Green Version]
- Barna, B.; Dám, A.L.; Király, Z. Juvenility and resistance of a superoxide tolerant plant to diseases and other stresses. Naturwissenschaften 1993, 80, 420–422. [Google Scholar] [CrossRef]
- Ort, D.R.; Merchant, S.S.; Alric, J.; Barkan, A.; Blankenship, R.E.; Bock, R.; Croce, R.; Hanson, M.R.; Hibberd, J.M.; Long, S.P.; et al. Redesigning photosynthesis to sustainably meet global food and bioenergy demand. Proc. Natl. Acad. Sci. USA 2015, 112, 8529–8536. [Google Scholar] [CrossRef] [Green Version]
- Roca, L.F.; Romero, J.; Bohorquez, J.M.; Alcantara, E.; Fernandez-Escobar, R.; Trapero, A. Nitrogen status affects growth, chlorophyll content and infection by Fusicladium oleagineum in olive. Crop Prot. 2018, 109, 80–85. [Google Scholar] [CrossRef]
- Alizade, S.; Keshtkar, E.; Mokhtasi-Bidgoli, A.; Sasanfar, H.; Streibig, J.C. Effect of water deficit stress on benzoylprop-ethyl performance and physiological traits of winter wild oat (Avena sterilis subsp. ludoviciana). Crop Prot. 2020, 137, 105292. [Google Scholar] [CrossRef]
- Maqsood, A.; Wu, H.; Kamran, M.; Altaf, H.; Mustafa, A.; Ahmar, S.; Nguyen Thi Thang, H.; Tariq, K.; He, Q.; Chen, J.T. Variations in growth, physiology, and antioxidative defense responses of two tomato (Solanum lycopersicum L.) cultivars after co-infection of Fusarium oxysporum and Meloidogyne incognita. Agronomy 2020, 10, 159. [Google Scholar]
- Gine, A.; Lopez-Gomez, M.; Vela, M.D.; Ornat, C.; Talavera, M.; Verdejo-Lucas, S.; Sorribas, F.J. Thermal requirements and population dynamics of root-knot nematodes on cucumber and yield losses under protected cultivation. Plant Pathol. 2014, 63, 1446–1453. [Google Scholar] [CrossRef]
- Kayani, M.Z.; Mukhtar, T.; Hussain, M.A. Effects of southern root knot nematode population densities and plant age on growth and yield parameters of cucumber. Crop Prot. 2017, 92, 207–212. [Google Scholar] [CrossRef]
- Cabasan, M.T.N.; Kumar, A.; De Waele, D. Effects of initial nematode population density and water regime on resistance and tolerance to the rice root-knot nematode Meloidogyne graminicola in African and Asian rice genotypes. Int. J. Pest Manag. 2018, 64, 252–261. [Google Scholar] [CrossRef]
- Win, P.P.; Kyi, P.P.; De Waele, D. Effect of agro-ecosystem on the occurrence of the rice root-knot nematode Meloidogyne graminicola on rice in Myanmar. Australas. Plant Pathol. 2011, 40, 187–196. [Google Scholar] [CrossRef]
- Ling, F.; Su, Q.; Jiang, H.; Cui, J.; He, X.; Wu, Z.; Zhang, Z.; Liu, J.; Zhao, Y. Effects of strigolactone on photosynthetic and physiological characteristics in salt-stressed rice seedlings. Sci. Rep. 2020, 10, 6183. [Google Scholar]
- Salalia, R.; Walia, R.K.; Somvanshi, V.S.; Kumar, P.; Kumar, A. Morphological, morphometric, and molecular characterization of intraspecific variations within Indian populations of Meloidogyne graminicola. J. Nematol. 2017, 49, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Haseeb, A.; Pandey, R.J.P.P.A.; Summaries, N. Root-knot nematodes—A constraint to cultivation of Davana, Artemisia pallens. Trop. Pest Manag. 1990, 36, 317–319. [Google Scholar] [CrossRef]
- Patil, J.; Gaur, H.S. The effect of root-knot nematode, Meloidogyne graminicola, on the quality and vigour of rice seed. Nematology 2014, 16, 555–564. [Google Scholar] [CrossRef]
- Narayana, R.; Sheela, M.; Lal, D.S. Effect of different inoculum levels of Meloidogyne graminicola golden and birchfield on growth and biochemical parameters of rice (Oryza sativa L.). Entomon 2015, 40, 163–168. [Google Scholar]
- Rampazzo, P.E.; Marcos, F.C.C.; Cipriano, M.A.P.; Marchiori, P.E.R.; Freitas, S.S.; Machado, E.C.; Nascimento, L.C.; Brocchi, M.; Ribeiro, R.V. Rhizobacteria improve sugarcane growth and photosynthesis under well-watered conditions. Ann. Appl. Biol. 2018, 172, 309–320. [Google Scholar] [CrossRef]
- Ahmad, S.; Kumar, R. Effect of root-knot nematode on chlorophyll content and plant growth of chickpea. Acta Bot. Indica 1990, 18, 314–315. [Google Scholar]
- Hisamuddin, S.S.; Azam, T. Pathogenicity of root-knot nematode, Meloidogyne incognita on Lens culinaris (Medik.). Arch. Phytopathol. Plant Prot. 2010, 43, 1504–1511. [Google Scholar] [CrossRef]
- Bhau, B.S.; Bitupon, B.; Reshma, A.; Phukon, P.; Barbi, G.; Sarmah, D.K.; Lal, M.; Wann, S.B. Influence of root-knot nematode infestation on antioxidant enzymes, chlorophyll content and growth in Pogostemon cablin (Blanco) Benth. Indian J. Exp. Biol. 2016, 54, 254–261. [Google Scholar]
- Liu, Y.; Xiang, Y.; Guo, W.; Gao, X.; Li, X. Photosynthetic physiology responses of Sapium sebiferum to the interactions between Meloidogyne incognita and aboveground herbivores with different diet breadths. Plant Prot. 2019, 45, 222–228. [Google Scholar]
- Wu, L.; Sun, L.; Peng, Y.; Zhang, Y.; Ma, X. Study on the effect of photovoltaic modules on greenhouse microenvironment and net photosynthetic rate of crops in greenhouse—a case study of strawberry greenhouse in Kunming. J. Yunnan Norm. Univ. 2021, 41, 10–15. [Google Scholar]
- Li, W. Effects of root-knot nematode on physiological and biochemical characteristics of cucumber in greenhouse. Shandong Agric. Univ. 2005, 7, 1–64. [Google Scholar]
- Huan-Zhong, L.I.; Zhang, J.L. Effect of spray application on Cotinus coggygria atropurpureus leaf changes of transpiration rate and intercellular carbon dioxide concentration. North. Hortic. 2011, 5, 94–97. [Google Scholar]
- Ma, X.; Jia, Z.; Liu, Y. Study on photosynthesis and chlorophyll metabolism of Avena sativa L. under salt stress. Chin. Qinghai J. Anim. Vet. Sci. 2021, 51, 7–14. [Google Scholar]
- Carneiro, R.G.; Mazzafera, P.; Ferraz, L.C.C.B.; Muraoka, T.; Trivelin, P.C.O. Uptake and translocation of nitrogen, phosphorus and calcium in soybean infected with Meloidogyne incognita and M. javanica. Trop. Plant Pathol. 2002, 27, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Wenting, H.; Deliang, P. Physiological and biochemical response of plant to nematode stress. Plant Prot. 2007, 33, 11–15. [Google Scholar]
- Saeed, I.; Macguidwin, A.E.; Rouse, D.I.; Sharkey, T.D. Limitation to photosynthesis in Pratylenchus penetrans- and Verticillium dahliae-infected potato. Crop Sci. 1999, 39, 1340–1346. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| J2 Population | Inoculation Time (Day) | ||
|---|---|---|---|
| 30 | 60 | 90 | |
| 0 | 106.4 ± 4.91 a | 138.0 ± 8.83 a | 184.6 ± 12.89 a |
| 500 | 116.4 ± 5.36 a | 131.8 ± 16.18 a | 149.4 ± 9.95 b |
| 1000 | 80.4 ± 2.13 b | 89.2 ± 5.67 b | 98.6 ± 6.68 c |
| 1500 | 80.2 ± 1.24 b | 86.4 ± 5.30 b | 111.0 ± 6.30 c |
| 2000 | 74.2 ± 3.18 b | 100.8 ± 6.71 b | 108.6± 2.69 c |
| J2 Population | Inoculation Time(Days) | ||
|---|---|---|---|
| 30 | 60 | 90 | |
| 0 | 93.89 ± 6.87 a | 131.00 ± 10.05 a | 169.71 ± 13.09 a |
| 250 | 103.00 ± 11.43 a | 126.67 ± 19.00 ab | 155.71 ± 25.04 a |
| 500 | 79.89 ± 11.26 ab | 115.57 ± 15.31 abc | 130.71 ± 13.99 ab |
| 750 | 62.50 ± 6.95 b | 80.87 ± 8.18 c | 83.14 ± 7.04 c |
| 1000 | 65.50 ± 5.51 b | 93.17 ± 6.10 bc | 93.50 ± 7.46 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Sikandar, A.; Ahmad, S.; Luo, M.; Wu, H. Meloidogyne graminicola’s Effect on Growth Performance of Rice under Low Population Density. Agronomy 2022, 12, 587. https://doi.org/10.3390/agronomy12030587
Chen B, Sikandar A, Ahmad S, Luo M, Wu H. Meloidogyne graminicola’s Effect on Growth Performance of Rice under Low Population Density. Agronomy. 2022; 12(3):587. https://doi.org/10.3390/agronomy12030587
Chicago/Turabian StyleChen, Bochang, Aatika Sikandar, Shakeel Ahmad, Man Luo, and Haiyan Wu. 2022. "Meloidogyne graminicola’s Effect on Growth Performance of Rice under Low Population Density" Agronomy 12, no. 3: 587. https://doi.org/10.3390/agronomy12030587