
Post-Release Evaluation of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and Tamarixia radiata (Hymenoptera: Eulophidae) for Biological Control of Diaphorina citri (Hemiptera: Liviidae) in Urban California, USA

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasitoid Release and Monitoring
2.1.1. Study Sites
2.1.2. Parasitoid Sources and Culture
2.1.3. Field Release and Monitoring
2.2. Parasitoid Establishment
2.3. Data Analysis
3. Results
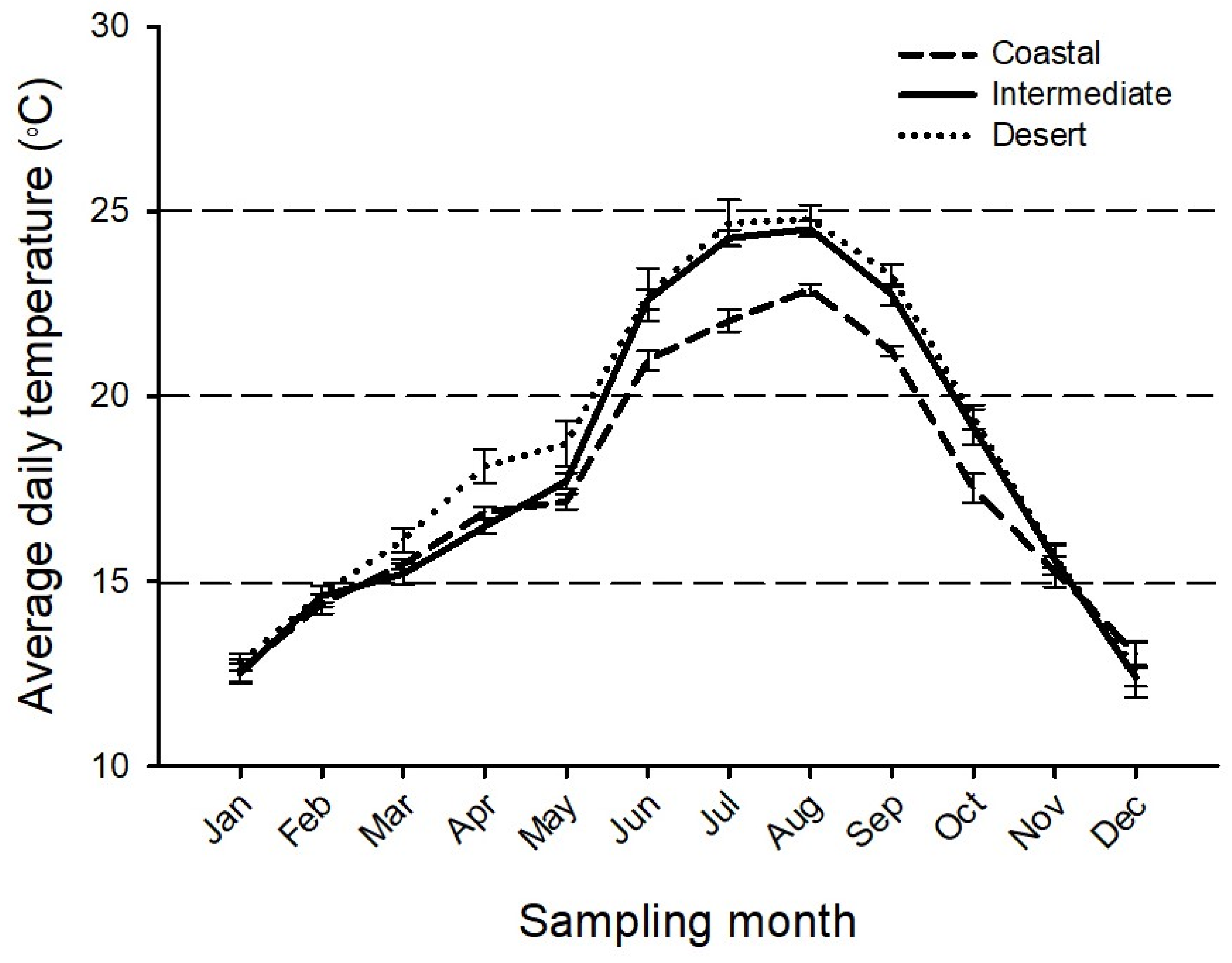
3.1. Effects of Climate Type and Sampling Month on Mean Daily Temperatures
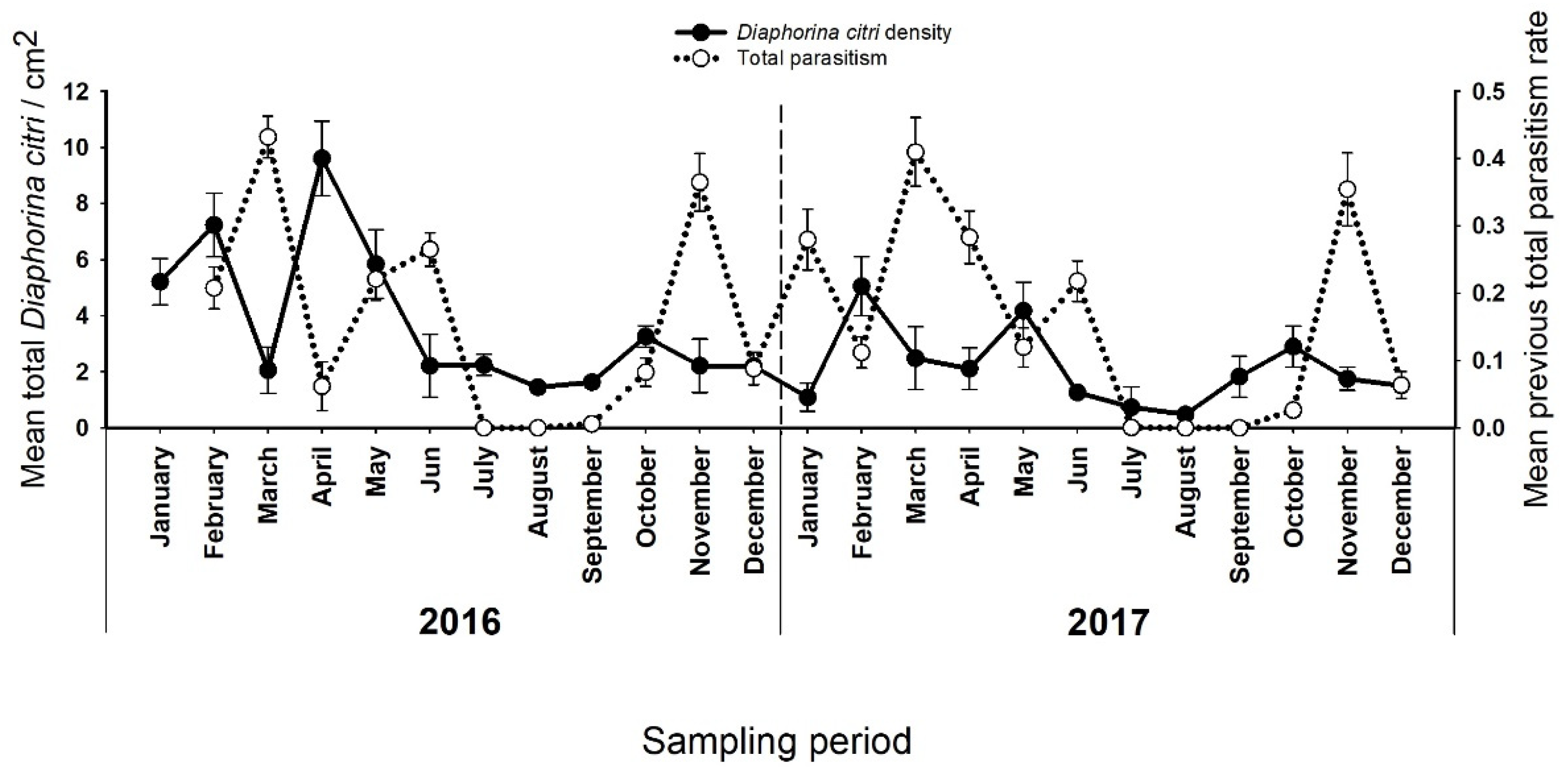
3.2. Effects of Climate Type, Season, and Parasitism on D. citri Densities
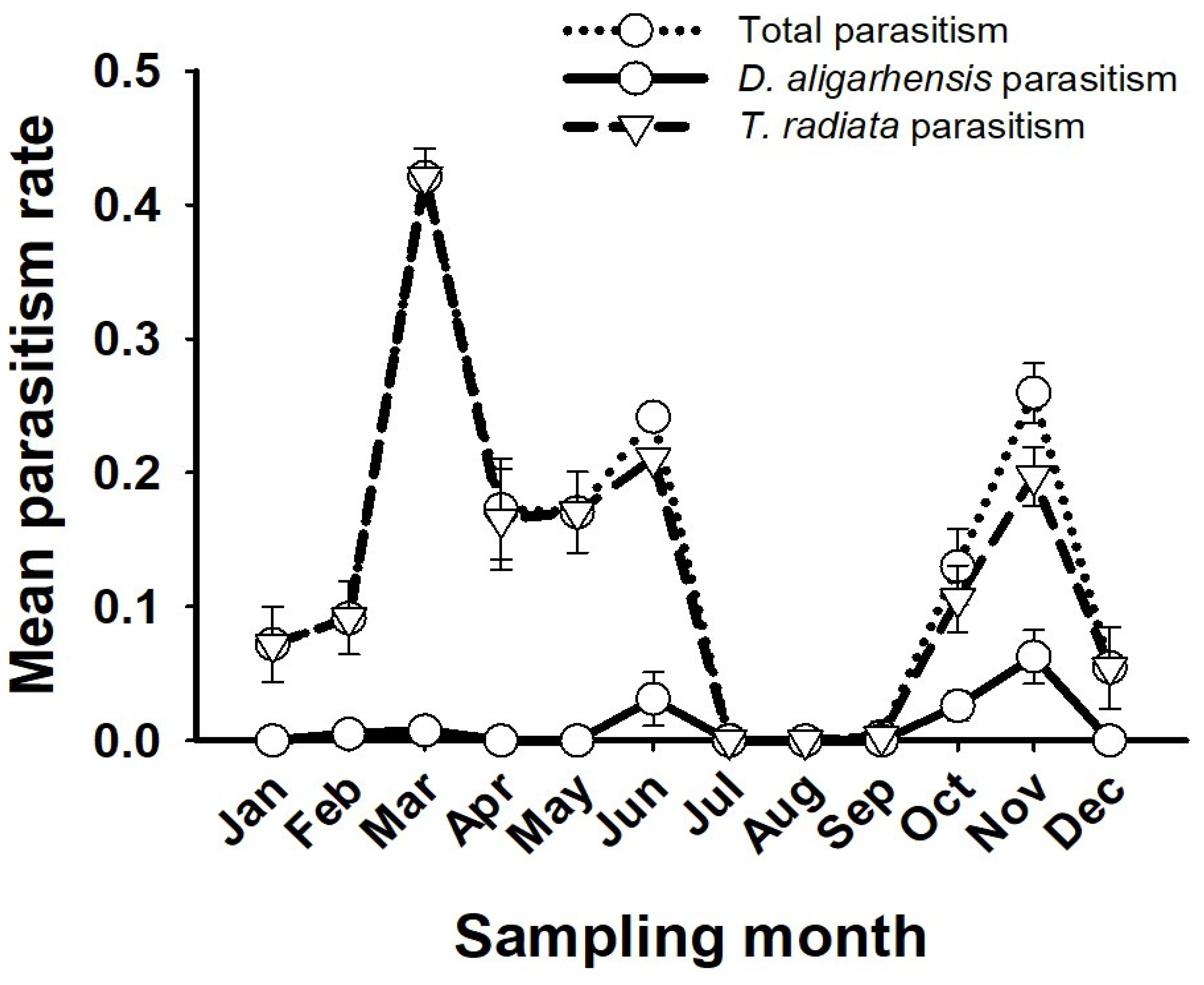
3.3. D. aligarhensis Establishment and Parasitism Rates
3.4. Effects on T. radiata Parasitism Rates and Recovery Success
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grafton-Cardwell, E.E. How serious is the threat of Asian citrus psyllid and Huanglongbing? Citrograph 2010, 1, 8–10. [Google Scholar]
- Bové, J.M. Huanglongbing: A destructive, newly emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar]
- Gottwald, T.R. Current epidemiological understanding of citrus Huanglongbing. Annu. Rev. Phytopathol. 2010, 48, 119–139. [Google Scholar] [CrossRef] [Green Version]
- Milne, A.E.; Gottwald, T.; Parnell, S.R.; Alonso Chavez, V.; Van den Bosch, F. What makes or breaks a campaign to stop an invading plant pathogen? PLoS Comput. Biol. 2020, 16, e1007570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayles, B.R.; Thomas, S.M.; Simmons, G.S.; Grafton-Cardwell, E.E.; Daugherty, M.P. Spatiotemporal dynamics of the Southern California Asian citrus psyllid (Diaphorina citri) invasion. PLoS ONE 2017, 12, e0173226. [Google Scholar]
- United States Department of Agriculture National Agricultural Statistics Services (USDA NASS). Citrus Production Forecast: 2019. Available online: www.nass.usda.gov/Statistics_by_State/Florida/Publications/Citrus/Citrus_Forecast/index.php (accessed on 10 January 2022).
- Babcock, B. Economic impact of California’s citrus industry. Citrograph 2018, 9, 36–39. [Google Scholar]
- Hoddle, M.S. Foreign exploration for natural enemies of Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae), in the Punjab of Pakistan for use in a classical biological control program in California USA. Pak. Entomol. 2012, 34, 1–5. [Google Scholar]
- Milosavljević, I.; Hoddle, M.S. Advances in classical biological control to support IPM of perennial agricultural crops. In Integrated Management of Insect Pests: Current and Future Developments in IPM; Kogan, M., Heinrichs, E.A., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 415–450. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Pandey, R. Host range testing of Tamarixia radiata (Hymenoptera: Eulophidae) sourced from the Punjab of Pakistan for classical biological control of Diaphorina citri (Hemiptera: Liviidae: Euphyllurinae: Diaphorinini) in California. J. Econ. Entomol. 2014, 107, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bistline-East, A.; Pandey, R.; Keçeci, M.; Hoddle, M.S. Host range testing of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) for use in classical biological control of Diaphorina citri (Hemiptera: Liviidae) in California. J. Econ. Entomol. 2015, 108, 940–950. [Google Scholar] [CrossRef]
- Milosavljević, I.; Schall, K.A.; Hoddle, C.D.; Morgan, D.J.W.; Hoddle, M.S. Biocontrol program targets Asian citrus psyllid in California’s urban areas. Calif. Agric. 2017, 71, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Bassanezi, R.B.; Montesino, L.H.; Gimenes-Fernandes, N.; Yamamoto, P.T.; Gottwald, T.R.; Amorim, L.; Filho, A.B. Efficacy of area-wide inoculum reduction and vector control on temporal progress of huanglongbing in young sweet orange plantings. Plant Dis. 2013, 97, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrig, E.; Shirk, P.D.; Hall, D.G.; Stansly, P.A. Larval development of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae), an endoparasitoid of Diaphorina citri (Hemiptera: Psyllidae). Ann. Entomol. Soc. Am. 2011, 104, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Sule, H.; Muhamad, R.; Omar, D.; Hee, A.K.W. Parasitism rate, host stage preference and functional response of Tamarixia radiata on Diaphorina citri. Int. J. Agric. Biol. 2014, 16, 783–788. [Google Scholar]
- Vankosky, M.A.; Hoddle, M.S. Two parasitoids of Diaphorina citri (Hemiptera: Liviidae) have shared, stage-specific preference for host nymphs that does not impact pest mortality rates. Fla. Entomol. 2019, 102, 49–58. [Google Scholar]
- Skelley, L.H.; Hoy, M.A. A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biol. Control 2004, 29, 14–23. [Google Scholar] [CrossRef]
- Rohrig, E. Biology and Behavior of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) an Endoparasitoid of Diaphorina citri (Hemiptera: Psyllidae). Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2010. [Google Scholar]
- Chen, X.; Stansly, P.A. Biology of Tamarixia radiata (Hymenoptera: Eulophidae), parasitoid of the citrus greening disease vector Diaphorina citri (Hemiptera: Psylloidea): A mini review. Fla. Entomol. 2014, 97, 1404–1413. [Google Scholar] [CrossRef]
- California Department of Food and Agriculture (CDFA). CDFA ACP/HLB Regulation and Quarantine Boundaries: 2021. Available online: https://www.cdfa.ca.gov/plant/acp/regulation.html (accessed on 10 January 2022).
- Hoddle, M.S.; Amrich, R.; Hoddle, C.D.; Kistner, E.J. Where’s Tamarixia? Citrograph 2016, 7, 64–66. [Google Scholar]
- Milosavljević, I.; Morgan, D.J.; Massie, R.E.; Hoddle, M.S. Density dependent mortality, climate, and Argentine ants affect population dynamics of an invasive citrus pest, Diaphorina citri, and its specialist parasitoid, Tamarixia radiata, in Southern California, USA. Biol. Control 2021, 159, 104627. [Google Scholar] [CrossRef]
- DeBach, P.; Sundby, R. Competitive displacement between ecological homologues. Hilgardia 1963, 34, 105–166. [Google Scholar] [CrossRef] [Green Version]
- Rohrig, E.A.; Hall, D.G.; Qureshi, J.A.; Stansly, P.A. Field release in Florida of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae), an endoparasitoid of Diaphorina citri (Homoptera: Psyllidae), from mainland China. Fla. Entomol. 2012, 95, 479–481. [Google Scholar] [CrossRef]
- Hoy, M.A.; Nguyen, R.; Jeyaprakash, A. Classical biological control of the Asian citrus psylla-release of Tamarixia radiata. Citrus Ind. 1999, 80, 20–22. [Google Scholar]
- Hoy, M.A.; Nguyen, R. Classical biological control of Asian citrus psylla. Citrus Ind. 2001, 81, 48–50. [Google Scholar]
- Qureshi, J.A.; Rogers, M.E.; Hall, D.G.; Stansly, P.A. Incidence of invasive Diaphorina citri (Hemiptera: Psyllidae) and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrus. J. Econ. Entomol. 2009, 102, 247–256. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). J. Econ. Entomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef]
- Quezada, J.; DeBach, P. Bioecological and population studies of the cottony-cushion scale, Icerya purchasi Mask., and its natural enemies, Rodolia cardinalis Mul. and Cryptochaetum iceryae Will., in southern California. Hilgardia 1973, 41, 631–688. [Google Scholar] [CrossRef] [Green Version]
- Daly, C.; Widrlechner, M.P.; Halbleib, M.D.; Smith, J.I.; Gibson, W.P. Development of a new USDA plant hardiness zone map for the United States. J. Appl. Meteorol. Climatol. 2012, 51, 242–264. [Google Scholar] [CrossRef] [Green Version]
- California Irrigation Management Information System (CIMIS). California Department of Water Resources’ California Irrigation Management Information System: State of California, Sacramento, CA: 2021. Available online: http://www.cimis.water.ca.gov (accessed on 10 January 2022).
- Hopper, K.R.; Roush, R.T.; Powell, W. Management of genetics of biological-control introductions. Ann. Rev. Entomol. 1993, 38, 27–51. [Google Scholar] [CrossRef]
- Roush, R.T.; Hopper, K.R. Use of single family lines to preserve genetic variation in laboratory colonies. Ann. Entomol. Soc. Am. 1995, 88, 713–717. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Hoddle, C.D. Classical biological control of Asian citrus psyllid with Tamarixia radiata in urban Southern California. Citrograph 2013, 4, 52–58. [Google Scholar]
- Ruais, D. The Effects of an Artificial Holding Diet on the Fecundity of the Ectoparasitic Wasp Tamarixia radiata (Waterston) (Hymenoptera: Eulophidae). Ph.D. Thesis, California State Polytechnic University, Pomona, CA, USA, 2018. [Google Scholar]
- Kistner, E.J.; Amrich, R.; Castillo, M.; Strode, V.; Hoddle, M.S. Phenology of Asian citrus psyllid (Hemiptera: Liviidae), with special reference to biological control by Tamarixia radiata, in the residential landscape of Southern California. J. Econ. Entomol. 2016, 109, 1047–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milosavljević, I.; Amrich, R.; Strode, V.; Hoddle, M.S. Modeling the phenology of Asian citrus psyllid (Hemiptera: Liviidae) in urban Southern California: Effects of environment, habitat, and natural enemies. Environ. Entomol. 2018, 47, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.Z.; Arif, M.J.; Hoddle, C.D.; Hoddle, M.S. Phenology of Asian citrus psyllid (Hemiptera: Liviidae) and associated parasitoids on two species of citrus, kinnow mandarin and sweet orange, in Punjab Pakistan. Environ. Entomol. 2014, 43, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Vankosky, M.A.; Hoddle, M.S. The effects of conspecific and heterospecific interactions on foraging and oviposition behaviours of two parasitoids of Diaphorina citri. Biocontrol Sci. Technol. 2017, 27, 739–754. [Google Scholar] [CrossRef]
- Vankosky, M.A.; Hoddle, M.S. An assessment of interspecific competition between two introduced parasitoids of Diaphorina citri (Hemiptera: Liviidae) on caged citrus plants. Insect Sci. 2019, 26, 119–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Institute SAS. SAS/STAT, Version 9.4 User’s Guide, 2nd ed.; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Warton, D.I.; Hui, F.K. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milosavljević, I.; Esser, A.D.; Crowder, D.W. Effects of environmental and agronomic factors on soil-dwelling pest communities in cereal crops. Agric. Ecosyst. Environ. 2016, 225, 192–198. [Google Scholar] [CrossRef]
- Whittingham, M.J.; Stephens, P.A.; Bradbury, R.B.; Freckleton, R.P. Why do we still use stepwise modelling in ecology and behaviour? J. Anim. Ecol. 2006, 75, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Hayat, M. Taxonomic notes on Indian Encyrtidae (Hym.: Chalcidoidea): II. J. Nat. Hist. 1981, 15, 17–29. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Hoddle, C.D.; Triapitsyn, S.V.; Khan, S.Z.; Arif, M.J. How many primary parasitoid species attack nymphs of Diaphorina citri (Hemiptera: Liviidae) in Punjab, Pakistan? Fla. Entomol. 2014, 97, 1825–1828. [Google Scholar] [CrossRef]
- Tang, Y.Q. On the parasite complex of Diaphorina citri Kuwayama (Homoptera: Psyllidae) in Asian-Pacific and other areas. In Proceedings of the 4th International Conference on Citrus Rehabilitation, Chiang Mai, Thailand, 4–10 February 1990; Volume 4, pp. 240–245. [Google Scholar]
- Bistline-East, A.; Hoddle, M.S. Chartocerus sp. (Hymenoptera: Signiphoridae) and Pachyneuron crassiculme (Hymenoptera: Pteromalidae) are obligate hyperparasitoids of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and possibly Tamarixia radiata (Hymenoptera: EulopHidae). Fla. Entomol. 2014, 97, 562–566. [Google Scholar] [CrossRef]
- Bistline-East, A.; Hoddle, M.S. Biology of Psyllaphycus diaphorinae (Hymenoptera: Encyrtidae), a hyperparasitoid of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and Tamarixia radiata (Hymenoptera: Eulophidae). Ann. Entomol. Soc. Am. 2016, 109, 22–28. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Bistline-East, A.; Hoddle, M.S. Threshold temperatures and thermal requirements of Psyllaphycus diaphorinae (Hymenoptera: Encyrtidae), a hyperparasitoid of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and Tamarixia radiata (Hymenoptera: Eulophidae). Fla. Entomol. 2020, 103, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Tena, A.; Hoddle, C.D.; Hoddle, M.S. Competition between honeydew producers in an ant-hemipteran interaction may enhance biological control of an invasive pest. Bull. Entomol. Res. 2013, 103, 714–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasio, O.E.; Mathis, K.A.; Rivera, M.J. Impacts of invasive ant-hemipteran interaction, edge effects and habitat complexities on the spatial distribution of ants in citrus orchards. Agric. Ecosyst. Environ. 2021, 310, 107299. [Google Scholar] [CrossRef]
- McCalla, K.A. Implications of Argentine Ant Management for Biological Control of the Asian Citrus Psyllid and Other Ant-Tended Pests in Citrus. Ph.D. Thesis, University of California, Riverside, CA, USA, 2019. [Google Scholar]
- Aubert, B.; Quilici, S. Biological control of the African and Asian citrus psyllids (Homoptera: Psylloidea), through eulophid and encyrtid parasites (Hymenoptera: Chalcidoidea) in Reunion Island. In Proceedings of the Ninth Conference of the International Organization of Citrus Virologist, Puerto Iguazu, Argentina, 9–13 May 1983; International Organization of Citrus Virologists: Riverside, CA, USA, 1984; Volume 9, pp. 100–108. [Google Scholar]
- Bové, J.M. Greening in the Arabian Peninsula: Toward new techniques for its detection and control. FAO Plant Protect. Bull. 1986, 34, 7–14. [Google Scholar]
- Al-Ghamdi, K.M.S.; Faragalla, A. Parasitism of Diaphoria [sic] citri (Kuwayama) (Homoptera: Psyllidae) by Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) on Mexican lime trees, western Saudi Arabia. Bio Sci. Res. Bull. 2000, 16, 99–105. [Google Scholar]
- Chiu, S.C.; Aubert, B.; Chien, C.C. Attempts to establish Tetrastichus radiatus Waterson (Hymenoptera, Chalcidoidea), a primary parasite of Diaphorina citri Kuwayama in Taiwan. In Proceedings of the Tenth Conference of the International Organization of Citrus Virologists, Valencia, Spain, 17–21 November 1986; International Organization of Citrus Virologists: Riverside, CA, USA, 1988; Volume 10, pp. 265–268. [Google Scholar]
- Kondo, T.; Wooley, J.B.; Arciniegas, K.T.; Campos-Patiño, Y. First report in Colombia and diagnosis of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae), a parasitoid wasp of Diaphorina citri (Hemiptera: Liviidae). Caldasia 2022, 44. [Google Scholar] [CrossRef]
- Gavarra, M.R.; Mercado, B.G.; Gonzales, C.I.; Aubert, B.; Tontyaporn, S.; Buangsuwon, D. Progress report: D. citri trapping, identification of parasite and possible field establishment of the imported parasite, Tamarixia radiata in the Philippines. In Proceedings of the Fourth International Asia Pacific Conference on Citrus Rehabilitation, Chiang Mai, Thailand, 12 September 1990; pp. 246–250. [Google Scholar]
- de Jesus, L.A.; Macasaet, A.A. Insect vector occurrence and greening disease spread in citrus orchard. Philipp. J. Crop Sci. 1991, 16, 23. [Google Scholar]
- Chow, A.; Sétamou, M. Parasitism of Diaphorina citri (Hemiptera: Liviidae) by Tamarixia radiata (Hymenoptera: Eulophidae) on residential citrus in Texas: Importance of colony size and instar composition. Biol. Control 2022, 165, 104796. [Google Scholar] [CrossRef]
- Hall, D.G.; Rohrig, E. Bionomics of Asian citrus psyllid (Hemiptera: Liviidae) associated with orange jasmine hedges in southeast central Florida, with special reference to biological control by Tamarixia radiata. J. Econ. Entomol. 2015, 108, 1198–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluke, R.W.; Qureshi, J.A.; Stansly, P.A. Citrus flushing patterns, Diaphorina citri (Hemiptera: Psyllidae) populations and parasitism by Tamarixia radiata (Hymenoptera: Eulophidae) in Puerto Rico. Fla. Entomol. 2008, 91, 36–42. [Google Scholar] [CrossRef]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The influence of temperature variation on life history parameters and thermal performance curves of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of the Asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef] [PubMed]
- Beirne, B.P. Avoidable obstacles to colonization in classical biological control of insects. Can. J. Zool. 1985, 63, 743–747. [Google Scholar] [CrossRef]
- Stiling, P. Why do natural enemies fail in classical biological control programs? Am. Entomol. 1993, 39, 31–37. [Google Scholar] [CrossRef]
- Van Driesche, R.G.; Hoddle, M.S. Classical arthropod biological control: Measuring success, step by step. In Biological Control: Measures of Success; Gurr, G.M., Wratten, S.D., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 39–75. [Google Scholar]
- Myers, J.H.; Higgins, C.; Kovacs, E. How many insect species are necessary for the biological control of insects? Environ. Entomol. 1989, 18, 541–547. [Google Scholar] [CrossRef]
- Denoth, M.; Frid, L.; Myers, J.H. Multiple agents in biological control: Improving the odds? Biol. Control 2002, 24, 20–30. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Climate Type | County | Site Name | Latitude (deg.) | Longitude (deg.) | Elev. (m) | Surveyed Tree(s) | CIMIS Station | Date of First Parasitoid Release | Most Recent Parasitoid Release | No. D. aligarhensis Release Events in 2015–2017 (2016–2017 Releases) | No. D. aligarhensis Released in 2015–2017 (2016–2017 Releases) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Coastal | Los Angeles | Alameda | N33°59′ | W118°14′ | 51 | Lemon, Orange | 174 | 31 August 15 | 7 November 16 | 39 (31) | 6388 (4711) |
| Orange | Anaheim | N33°51′ | W117°44′ | 316 | Lemon, Orange | 75 | 17 August 15 | 1 March 17 | 49 (32) | 7006 (5602) | |
| Fullerton | N33°53′ | W117°53′ | 76 | Lemon, Orange | 75 | 17 August 15 | 15 March 17 | 37 (31) | 6419 (4072) | ||
| Irvine | N33°43′ | W117°47′ | 33 | Lemon, Orange | 75 | 8 September 15 | 15 March 17 | 46 (31) | 5615 (4904) | ||
| San Diego | Fallbrook | N33°23′ | W117°10′ | 197 | Lemon, Orange | 62 | 21 September 15 | 8 February 17 | 42 (35) | 4389 (3406) | |
| Intermediate | Los Angeles | Claremont | N34°05′ | W117°42′ | 341 | Lemon, Orange | 78 | 31 August 15 | 27 March 17 | 26 (24) | 2616 (2256) |
| Pomona | N34°03′ | W117°44′ | 255 | Lemon, Orange | 78 | 31 August 15 | 27 March 17 | 39 (31) | 5595 (3512) | ||
| Riverside | Citrus Hill | N33°51′ | W117°19′ | 542 | Lemon, Orange | 44 | 21 September 15 | 8 February 17 | 30 (28) | 3102 (2858) | |
| Woodcrest | N33°54′ | W117°20′ | 479 | Lemon, Orange | 44 | 10 August 15 | 8 February 17 | 30 (23) | 4449 (2929) | ||
| San Diego | Ramona | N33°01′ | W116°49′ | 443 | Lemon, Orange | 153 | 24 August 15 | 22 March 17 | 49 (28) | 6524 (4961) | |
| Desert | Riverside | Hemet | N33°44′ | W116°49′ | 645 | Lemon, Orange | 239 | 8 September 15 | 20 March 17 | 20 (18) | 2603 (1952) |
| San Bernardino | Redlands | N34°02′ | W117°13′ | 403 | Lemon, Orange | 251 | 5 October 15 | 20 March 17 | 29 (21) | 3058 (2029) | |
| Warner * | N34°04′ | W117°10′ | 440 | Lemon, Orange | 251 | 5 October 15 | 29 March 17 | 17 (15) | 1981 (1151) | ||
| Crafton | N34°03′ | W117°07′ | 600 | Lemon, Orange | 251 | 5 October 15 | 29 March 17 | 34 (27) | 3684 (2671) | ||
| San Diego | Valley Ctr. * | N33°15′ | W117°03′ | 348 | Lemon, Orange | 207 | 20 July 15 | 8 February 17 | 36 (29) | 5542 (3191) |
| Parameter | Estimate | SE | df | t | p | 95% Conf. Interval | ||
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| (A) Psyllid densities | ||||||||
| Climate type | Coastal_Intermediate | 0.3094 | 0.5231 | 54.33 | 0.59 | 0.82 | −0.7393 | 1.3581 |
| Coastal_Desert | 0.3662 | 0.5796 | 92.48 | 0.63 | 0.81 | −0.7848 | 1.5171 | |
| Desert_Intermediate | −0.0568 | 0.5742 | 70.74 | -0.11 | 0.99 | −1.2018 | 1.0882 | |
| Year | 2016_2017 | 0.3126 | 0.1389 | 634.9 | 2.25 | 0.048 * | 0.0126 | 0.6126 |
| Season | Cool_Moderate | −0.2295 | 0.5874 | 74.82 | -0.39 | 0.92 | −1.3998 | 0.9407 |
| Cool_Warm | 1.5459 | 0.6485 | 105.7 | 2.38 | 0.02 * | 0.6201 | 2.3817 | |
| Moderate_Warm | 1.7755 | 0.5616 | 136.3 | 3.16 | 0.006 * | 0.6648 | 2.8861 | |
| Crop | Lemon_Orange | 0.5701 | 0.4282 | 673.9 | 1.33 | 0.19 | −0.2708 | 1.4109 |
| Flush abundance | 6.6711 | 0.5043 | 689 | 13.23 | <0.001 * | 5.6808 | 7.6611 | |
| Total parasitism | −0.0326 | 0.0119 | 677.8 | −2.74 | 0.006 * | −0.0549 | −0.0092 | |
| (B) Parasitism of T. radiata | ||||||||
| Climate type | Coastal_Intermediate | 0.0964 | 0.4858 | 35.08 | 0.19 | 0.97 | −0.9728 | 1.1655 |
| Coastal_Desert | 0.5147 | 0.5604 | 18.97 | 0.92 | 0.65 | −0.7187 | 1.7481 | |
| Desert_Intermediate | −0.4183 | 0.5509 | 26.24 | −0.76 | 0.73 | −1.6308 | 0.7941 | |
| Year | 2016_2017 | 0.6787 | 0.4112 | 465 | 1.65 | 0.13 | −1.5669 | 0.2098 |
| Season | Cool_Moderate | 0.7115 | 0.4696 | 55.84 | 1.52 | 0.13 | −1.6344 | 0.2113 |
| Cool_Warm | 4.0058 | 0.6947 | 74.18 | 5.77 | <0.001 * | 2.5750 | 5.4365 | |
| Moderate_Warm | 2.3531 | 0.5845 | 76.5 | 4.03 | <0.001 * | 1.1492 | 3.5568 | |
| Crop | Lemon_Orange | 0.2844 | 0.3456 | 413.5 | 0.79 | 0.44 | −0.4816 | 1.0505 |
| Flush abundance | 1.6528 | 0.5425 | 463.4 | 3.05 | 0.009 * | 0.5355 | 2.7700 | |
| Large nymphs | −0.7012 | 0.1989 | 382.6 | 3.52 | <0.001 * | 0.3102 | 1.0922 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milosavljević, I.; Vankosky, M.A.; Morgan, D.J.W.; Hoddle, C.D.; Massie, R.E.; Hoddle, M.S. Post-Release Evaluation of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and Tamarixia radiata (Hymenoptera: Eulophidae) for Biological Control of Diaphorina citri (Hemiptera: Liviidae) in Urban California, USA. Agronomy 2022, 12, 583. https://doi.org/10.3390/agronomy12030583
Milosavljević I, Vankosky MA, Morgan DJW, Hoddle CD, Massie RE, Hoddle MS. Post-Release Evaluation of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and Tamarixia radiata (Hymenoptera: Eulophidae) for Biological Control of Diaphorina citri (Hemiptera: Liviidae) in Urban California, USA. Agronomy. 2022; 12(3):583. https://doi.org/10.3390/agronomy12030583
Chicago/Turabian StyleMilosavljević, Ivan, Meghan A. Vankosky, David J. W. Morgan, Christina D. Hoddle, Rachael E. Massie, and Mark S. Hoddle. 2022. "Post-Release Evaluation of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae) and Tamarixia radiata (Hymenoptera: Eulophidae) for Biological Control of Diaphorina citri (Hemiptera: Liviidae) in Urban California, USA" Agronomy 12, no. 3: 583. https://doi.org/10.3390/agronomy12030583