
De Novo Domestication Concept for Potato Germplasm Enhancement

 , ,
, ,
Abstract
:1. Introduction
2. Pros and Cons of Wild Species for Potato Breeding
3. Agronomically Important Genes of Wild Potato
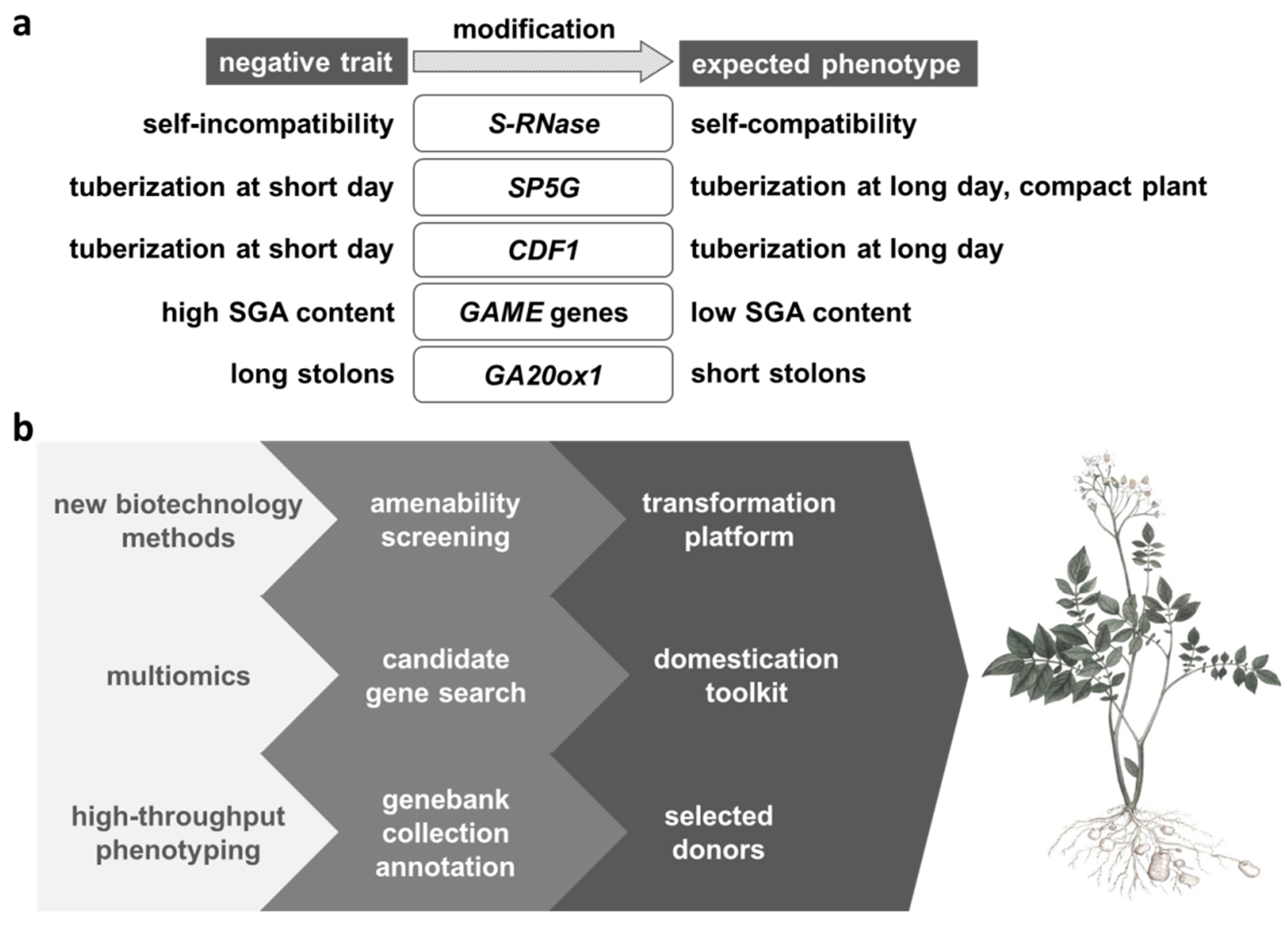
4. Candidate Genes for Correction of Undesirable Traits in Wild Potato
5. The Strategy for Trait Donor Improvement
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Li, Y.; Colleoni, C.; Zhang, J.; Liang, Q.; Hu, Y.; Ruess, H.; Simon, R.; Liu, Y.; Liu, H.; Yu, G.; et al. Genomic Analyses Yield Markers for Identifying Agronomically Important Genes in Potato. Mol. Plant 2018, 11, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bethke, P.C.; Halterman, D.A.; Jansky, S.H. Potato Germplasm Enhancement Enters the Genomics Era. Agronomy 2019, 9, 575. [Google Scholar] [CrossRef] [Green Version]
- Bashir, I.; Nicolao, R.; Heiden, G. Wild Potatoes: A Genetic Reservoir for Potato Breeding. In Wild Germplasm Genet. Improv. Crop Plants; Academic Press, Elsevier: Amsterdam, The Netherlands, 2021; pp. 215–240. [Google Scholar]
- Maune, J.F.; Camadro, E.L.; Erazzú, L.E. Cross-incompatibility and self-incompatibility: Unrelated phenomena in wild and cultivated potatoes? Botany 2018, 96, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Fernie, A.R.; Alseekh, S.; Liu, J.; Yan, J. Using precision phenotyping to inform de novo domestication. Plant Physiol. 2021, 186, 1397–1411. [Google Scholar] [CrossRef]
- Zsögön, A.; Cermak, T.; Voytas, D.; Peres, L.E.P. Genome editing as a tool to achieve the crop ideotype and de novo domestication of wild relatives: Case study in tomato. Plant Sci. 2017, 256, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018, 36, 1211–1216. [Google Scholar] [CrossRef] [Green Version]
- Eck, J. Van Genome editing and plant transformation of solanaceous food crops. Curr. Opin. Biotechnol. 2018, 49, 35–41. [Google Scholar] [CrossRef]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef]
- Hardigan, M.A.; Laimbeer, F.P.E.; Newton, L.; Crisovan, E.; Hamilton, J.P.; Vaillancourt, B.; Wiegert-Rininger, K.; Wood, J.C.; Douches, D.S.; Farré, E.M.; et al. Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proc. Natl. Acad. Sci. USA 2017, 114, E9999–E10008. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, J.G. The Potato: Evolution, Biodiversity and Genetic Resources; Belhaven Press: London, UK, 1990; pp. 1–259. [Google Scholar]
- Ross, H. Potato Breeding. Problems and Perspectives; V.P. Parey: Berlin, Germany, 1986; pp. 1–132. [Google Scholar]
- Bukasov, S.M.; Kameraz, A.Y. Basics of Potato Breeding; Selhozgis: Moscow, Russia, 1959; pp. 1–525. (In Russian) [Google Scholar]
- Jansky, S.H.; De Jong, W.S.; Douches, D.S.; Haynes, K.G.; Holm, D.G. Cultivar Improvement with Exotic Germplasm: An Example from Potato. In The Wild Solanums Genomes. Compendium of Plant Genomes; Carputo, D., Aversano, R., Ercolano, M.R., Eds.; Springer: Cham, Switzerland, 2021; pp. 215–230. [Google Scholar] [CrossRef]
- Gavrilenko, T.A.; Yermishin, A.P. Interspecific hybridization of potato: Theoretical and applied aspects. Vavilov J. Genet. Breed. 2017, 21, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Kostina, L.I.; Kosareva, O.S. Genealogy of Domestic Potato Varieties; VIR: Saint Petersburg, Russia, 2017; pp. 1–72. (In Russian) [Google Scholar]
- Pavek, J.J.; Corsini, D.L. Utilization of potato genetic resources in variety development. Am. J. Potato Res. 2001, 78, 433–441. [Google Scholar] [CrossRef]
- Bradshaw, J.E. Potato Breeding at the Scottish Plant Breeding Station and the Scottish Crop Research Institute: 1920–2008. Potato Res. 2009, 52, 141–172. [Google Scholar] [CrossRef]
- Yermishin, A.P.; Voronkova, E.V.; Kozlov, V.A.; Polyuhovich, Y.V.; Luksha, V.I.; Levy, A.V.; Yakovleva, G.A.; Semanyuk, T.V.; Dubinich, V.L.; Rodkina, I.A. Interspecific Hybridization in Potato Breeding; Yermishin, A.P., Ed.; Institute of Genetics and Cytology NAS of Belarus: Minsk, Belarus, 2021; pp. 1–396. (In Belarus) [Google Scholar]
- Budin, K.Z. Genetic Basis for the Creation of Potato Donors; VIR: Saint Petersburg, Russia, 1997; pp. 1–40. (In Russian) [Google Scholar]
- Budin, K.Z.; Gorbatenco, L.E.; Turuleva, L.M. Mexican Potato Species and Their Importance for Breeding; VIR: Saint Petersburg, Russia, 1989; pp. 1–89. (In Russian) [Google Scholar]
- Zoteeva, N.M.; Khzhanovska, M.; Evstratova, L.P.; Fasulati, S.R.; Yusupov, T.M. Resistance of Wild Potato Species to Diseases and Pests; VIR: Saint Petersburg, Russia, 2004; pp. 1–88. (In Russian) [Google Scholar]
- Gorbatenco, L.E. Potato Species of South America (Ecology, Geography, Introduction, Taxonomy and Breeding Value); VIR: Saint Petersburg, Russia, 2006; pp. 1–456. [Google Scholar]
- Rogozina, E.V.; Khavkin, E.E.; Kuznetsova, M.A.; Gavrilenko, T.A.; Chalaya, N.A.; Beketova, M.P.; Sokolova, E.A.; Antonova, O.Y.; Fadina, O.A.; Smetanina, T.I. Clonal Collection of Wild Potato Species; VIR: Saint Petersburg, Russia, 2015; pp. 1–45. (In Russian) [Google Scholar]
- Tingey, W.M.; Mackenzie, J.D.; Gregory, P. Total foliar glycoalkaloids and resistance of wild potato species toEmpoasca fabae (Harris). Am. Potato J. 1978, 55, 577–585. [Google Scholar] [CrossRef]
- Deahl, K.L.; Sinden, S.L.; Young, R.J. Evaluation of wild tuber-bearing Solanum accessions for foliar glycoalkaloid level and composition. Am. Potato J. 1993, 70, 61–69. [Google Scholar] [CrossRef]
- Leesutthiphonchai, W.; Vu, A.L.; Ah-Fong, A.M.V.; Judelson, H.S. How Does Phytophthora infestans Evade Control Efforts? Modern Insight Into the Late Blight Disease. Phytopathology 2018, 108, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.A.; Ukladov, E.O.; Golubeva, T.S. Phytophthora infestans: An Overview of Methods and Attempts to Combat Late Blight. J. Fungi 2021, 7, 1071. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Thieme, R.; König, J.; Nachtigall, M.; Hammann, T.; Denes, T.E.; Kruppa, K.; Molnár-Láng, M. Introgression of Two Broad-Spectrum Late Blight Resistance Genes, Rpi-Blb1 and Rpi-Blb3, From Solanum bulbocastanum Dun Plus Race-Specific R Genes Into Potato Pre-breeding Lines. Front. Plant Sci. 2020, 11, 699. [Google Scholar] [CrossRef] [PubMed]
- Sanetomo, R.; Habe, I.; Hosaka, K. Sexual introgression of the late blight resistance gene Rpi-blb3 from a Mexican wild diploid species Solanum pinnatisectum Dunal into potato varieties. Mol. Breed. 2019, 39, 13. [Google Scholar] [CrossRef]
- Ghislain, M.; Byarugaba, A.A.; Magembe, E.; Njoroge, A.; Rivera, C.; Román, M.L.; Tovar, J.C.; Gamboa, S.; Forbes, G.A.; Kreuze, J.F.; et al. Stacking three late blight resistance genes from wild species directly into African highland potato varieties confers complete field resistance to local blight races. Plant Biotechnol. J. 2019, 17, 1119–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elison, G.L.; Novy, R.G.; Whitworth, J.L. Russet Potato Breeding Clones with Extreme Resistance to Potato Virus Y Conferred by Rychc as well as Resistance to Late Blight and Cold-Induced Sweetening. Am. J. Potato Res. 2021, 98, 411–419. [Google Scholar] [CrossRef]
- Bradeen, J.M. On the Value of Wild Solanum Species for Improved Crop Disease Resistance: Resistances to Nematodes and Viruses. In the Wild Solanums Genomes; Springer: Berlin/Heidelberg, Germany, 2021; pp. 95–118. [Google Scholar] [CrossRef]
- Naess, S.K.; Bradeen, J.M.; Wielgus, S.M.; Haberlach, G.T.; McGrath, J.M.; Helgeson, J.P. Resistance to late blight in Solanum bulbocastanum is mapped to chromosome 8. Theor. Appl. Genet. 2000, 101, 697–704. [Google Scholar] [CrossRef]
- Van Der Vossen, E.A.G.; Gros, J.; Sikkema, A.; Muskens, M.; Wouters, D.; Wolters, P.; Pereira, A.; Allefs, S. The Rpi-blb2 gene from Solanum bulbocastanum is an Mi-1 gene homolog conferring broad-spectrum late blight resistance in potato. Plant J. 2005, 44, 208–222. [Google Scholar] [CrossRef]
- Park, T.H.; Gros, J.; Sikkema, A.; Vleeshouwers, V.G.A.A.; Muskens, M.; Allefs, S.; Jacobsen, E.; Visser, R.G.F.; Van Der Vossen, E.A.G. The late blight resistance locus Rpi-bib3 from Solanum bulbocastanum belongs to a major late blight R gene cluster on chromosome 4 of potato. Mol. Plant Microbe Interact. 2005, 18, 722–729. [Google Scholar] [CrossRef] [Green Version]
- Oosumi, T.; Rockhold, D.R.; Maccree, M.M.; Deahl, K.L.; McCue, K.F.; Belknap, W.R. Gene Rpi-bt1 from Solanum bulbocastanum confers resistance to late blight in transgenic potatoes. Am. J. Potato Res. 2009, 86, 456–465. [Google Scholar] [CrossRef]
- Brown, C.R.; Yang, C.P.; Mojtahedi, H.; Santo, G.S.; Masuelli, R. RFLP analysis of resistance to Columbia root-knot nematode derived from Solanum bulbocastanum in a BC2 population. Theor. Appl. Genet. 1996, 92, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Hein, I.; Birch, P.R.J.; Danan, S.; Lefebvre, V.; Odeny, D.A.; Gebhardt, C.; Trognitz, F.; Bryan, G.J. Progress in Mapping and Cloning Qualitative and Quantitative Resistance Against Phytophthora infestans in Potato and Its Wild Relatives. Potato Res. 2009, 52, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Śliwka, J.; Jakuczun, H.; Chmielarz, M.; Hara-Skrzypiec, A.; Tomczyńska, I.; Kilian, A.; Zimnoch-Guzowska, E. A resistance gene against potato late blight originating from Solanum × michoacanum maps to potato chromosome VII. Theor. Appl. Genet. 2012, 124, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Kuhl, J.; Hanneman, R.; Havey, M. Characterization and mapping of Rpi1, a late-blight resistance locus from diploid (1EBN) Mexican Solanum pinnatisectum. Mol. Genet. Genom. 2001, 265, 977–985. [Google Scholar] [CrossRef]
- Jacobs, M.M.J.; Vosman, B.; Vleeshouwers, V.G.A.A.; Visser, R.G.F.; Henken, B.; van den Berg, R.G. A novel approach to locate Phytophthora infestans resistance genes on the potato genetic map. Theor. Appl. Genet. 2010, 120, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Rietman, H. Putting the Phytophthora Infestans Genome Sequence at Work: Multiple Novel Avirulence and Potato Resistance Gene Candidates Revealed. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2011. [Google Scholar]
- Hosaka, K.; Hosaka, Y.; Mori, M.; Maida, T.; Matsunaga, H. Detection of a simplex RAPD marker linked to resistance to potato virus Y in a tetraploid potato. Am. J. Potato Res. 2001, 78, 191–196. [Google Scholar] [CrossRef]
- Cockerham, G. Genetical studies on resistance to potato viruses X and Y. Heredity 1970, 25, 309–348. [Google Scholar] [CrossRef]
- Marczewski, W.; Strzelczyk-Żyta, D.; Hennig, J.; Witek, K.; Gebhardt, C. Potato chromosomes IX and XI carry genes for resistance to potato virus M. Theor. Appl. Genet. 2006, 112, 1232–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villamon, F.G.; Spooner, D.M.; Orrillo, M.; Mihovilovich, E.; Pérez, W.; Bonierbale, M. Late blight resistance linkages in a novel cross of the wild potato species Solanum paucissectum (series Piurana). Theor. Appl. Genet. 2005, 111, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Smilde, W.D.; Brigneti, G.; Jagger, L.; Perkins, S.; Jones, J.D.G. Solanum mochiquense chromosome IX carries a novel late blight resistance gene Rpi-moc1. Theor. Appl. Genet. 2005, 110, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Danan, S.; Chauvin, J.-E.; Caromel, B.; Moal, J.-D.; Pellé, R.; Lefebvre, V. Major-effect QTLs for stem and foliage resistance to late blight in the wild potato relatives Solanum sparsipilum and S. spegazzinii are mapped to chromosome X. Theor. Appl. Genet. 2009, 119, 705–719. [Google Scholar] [CrossRef]
- Moury, B.; Caromel, B.; Johansen, E.; Simon, V.; Chauvin, L.; Jacquot, E.; Kerlan, C.; Lefebvre, V. The Helper Component Proteinase Cistron of Potato virus Y Induces Hypersensitivity and Resistance in Potato Genotypes Carrying Dominant Resistance Genes on Chromosome IV. Mol. Plant-Microbe Interact. 2011, 24, 787–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caromel, B.; Mugniéry, D.; Kerlan, M.-C.; Andrzejewski, S.; Palloix, A.; Ellissèche, D.; Rousselle-Bourgeois, F.; Lefebvre, V. Resistance Quantitative Trait Loci Originating from Solanum sparsipilum Act Independently on the Sex Ratio of Globodera pallida and Together for Developing a Necrotic Reaction. Mol. Plant-Microbe Interact. 2005, 18, 1186–1194. [Google Scholar] [CrossRef] [Green Version]
- Park, T.-H.; Foster, S.; Brigneti, G.; Jones, J.D.G. Two distinct potato late blight resistance genes from Solanum berthaultii are located on chromosome 10. Euphytica 2009, 165, 269–278. [Google Scholar] [CrossRef]
- Tan, M.Y.A.; Hutten, R.C.B.; Celis, C.; Park, T.-H.; Niks, R.E.; Visser, R.G.F.; van Eck, H.J. The R Pi-mcd1 Locus from Solanum microdontum Involved in Resistance to Phytophthora infestans, Causing a Delay in Infection, Maps on Potato Chromosome 4 in a Cluster of NBS-LRR Genes. Mol. Plant-Microbe Interact. 2008, 21, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Barone, A.; Ritter, E.; Schachtschabel, U.; Debener, T.; Salamini, F.; Gebhardt, C. Localization by restriction fragment length polymorphism mapping in potato of a major dominant gene conferring resistance to the potato cyst nematode Globodera rostocbiensis. Mol. Genet. Genom. MGG 1990, 224, 177–182. [Google Scholar] [CrossRef]
- Paal, J.; Henselewski, H.; Muth, J.; Meksem, K.; Menéndez, C.M.; Salamini, F.; Ballvora, A.; Gebhardt, C. Molecular cloning of the potato Gro1-4 gene conferring resistance to pathotype Ro1 of the root cyst nematode Globodera rostochiensis, based on a candidate gene approach. Plant J. 2004, 38, 285–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreike, C.M.; de Koning, J.R.A.; Vinke, J.H.; van Ooijen, J.W.; Stiekema, W.J. Quantitatively-inherited resistance to Globodera pallida is dominated by one major locus in Solanum spegazzinii. Theor. Appl. Genet. 1994, 88, 764–769. [Google Scholar] [CrossRef]
- Caromel, B.; Mugniéry, D.; Lefebvre, V.; Andrzejewski, S.; Ellissèche, D.; Kerlan, M.C.; Rousselle, P.; Rousselle-Bourgeois, F. Mapping QTLs for resistance against Globodera pallida (Stone) Pa2/3 in a diploid potato progeny originating from Solanum spegazzinii. Theor. Appl. Genet. 2003, 106, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.J.; Park, T.-H.; Pel, M.; Brigneti, G.; Śliwka, J.; Jagger, L.; van der Vossen, E.; Jones, J.D.G. Rpi-vnt1.1, a Tm-2 2 Homolog from Solanum venturii, Confers Resistance to Potato Late Blight. Mol. Plant-Microbe Interact. 2009, 22, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querci, M. Analysis of the Resistance-Breaking Determinants of Potato Virus X (PVX) Strain HB on Different Potato Genotypes Expressing Extreme Resistance to PVX. Phytopathology 1995, 85, 1003. [Google Scholar] [CrossRef]
- Jacobs, J.M.E.; van Eck, H.J.; Horsman, K.; Arens, P.F.P.; Verkerk-Bakker, B.; Jacobsen, E.; Pereira, A.; Stiekema, W.J. Mapping of resistance to the potato cyst nematode Globodera rostochiensis from the wild potato species Solanum vernei. Mol. Breed. 1996, 2, 51–60. [Google Scholar] [CrossRef]
- Aversano, R.; Contaldi, F.; Ercolano, M.R.; Grosso, V.; Iorizzo, M.; Tatino, F.; Xumerle, L.; Molin, A.D.; Avanzato, C.; Ferrarini, A.; et al. The Solanum commersonii genome sequence provides insights into adaptation to stress conditions and genome evolution of wild potato relatives. Plant Cell 2015, 27, 954–968. [Google Scholar] [CrossRef] [Green Version]
- Blennow, A.; Skryhan, K.; Tanackovic, V.; Krunic, S.L.; Shaik, S.S.; Andersen, M.S.; Kirk, H.G.; Nielsen, K.L. Non-GMO potato lines, synthesizing increased amylose and resistant starch, are mainly deficient in isoamylase debranching enzyme. Plant Biotechnol. J. 2020, 18, 2096. [Google Scholar] [CrossRef]
- Braun, S.R.; Endelman, J.B.; Haynes, K.G.; Jansky, S.H. Quantitative Trait Loci for Resistance to Common Scab and Cold-Induced Sweetening in Diploid Potato. Plant Genome 2017, 10, plantgenome2016.10.0110. [Google Scholar] [CrossRef] [Green Version]
- Bali, S.; Robinson, B.R.; Sathuvalli, V.; Bamberg, J.; Goyer, A. Single Nucleotide Polymorphism (SNP) markers associated with high folate content in wild potato species. PLoS ONE 2018, 13, e0193415. [Google Scholar] [CrossRef] [Green Version]
- Love, S.L.; Pavek, J.J.; Thompson-Johns, A.; Bohl, W. Breeding progress for potato chip quality in North American cultivars. Am. J. Potato Res. 1998, 75, 27–36. [Google Scholar] [CrossRef]
- Ivanova, K.A.; Gerasimova, S.V.; Khlestkina, E.K. The Biosynthesis Regulation of Potato Steroidal Glycoalkaloids. Vavilov J. Genet. Breed. 2018, 22, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.K.; Zhao, Y.; Chen, S.Y.; Kennelly, E.J. Solanum steroidal glycoalkaloids: Structural diversity, biological activities, and biosynthesis. Nat. Prod. Rep. 2021, 38, 1423–1444. [Google Scholar] [CrossRef]
- Nakayasu, M.; Akiyama, R.; Lee, H.J.; Osakabe, K.; Osakabe, Y.; Watanabe, B.; Sugimoto, Y.; Umemoto, N.; Saito, K.; Muranaka, T.; et al. Generation of α-solanine-free hairy roots of potato by CRISPR/Cas9 mediated genome editing of the St16DOX gene. Plant Physiol. Biochem. 2018, 131, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Sawai, S.; Ohyama, K.; Yasumoto, S.; Seki, H.; Sakuma, T.; Yamamoto, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H.; Aoki, T.; et al. Sterol Side Chain Reductase 2 Is a Key Enzyme in the Biosynthesis of Cholesterol, the Common Precursor of Toxic Steroidal Glycoalkaloids in Potato. Plant Cell 2014, 26, 3763–3774. [Google Scholar] [CrossRef] [Green Version]
- Somalraju, A.; Ghose, K.; Main, D.; Mccallum, J. Altered Steroidal Glycoalkaloid (SGA) Biosynthesis in Diploid Potatoes as Induced by EMS-Mutagenesis. Crop Breed. Genet. Genom. 2020, 4, 2. [Google Scholar] [CrossRef]
- McCue, K.F.; Breksa, A.; Vilches, A.; Belknap, W.R. Modification of Potato Steroidal Glycoalkaloids with Silencing RNA Constructs. Am. J. Potato Res. 2018, 95, 9–14. [Google Scholar] [CrossRef]
- Paudel, J.R.; Davidson, C.; Song, J.; Maxim, I.; Aharoni, A.; Tai, H.H. Pathogen and pest responses are altered due to RNAi-mediated knockdown of GLYCOALKALOID METABOLISM 4 in Solanum tuberosum. Mol. Plant-Microbe Interact. 2017, 30, 876–885. [Google Scholar] [CrossRef]
- Akiyama, R.; Watanabe, B.; Nakayasu, M.; Lee, H.J.; Kato, J.; Umemoto, N.; Muranaka, T.; Saito, K.; Sugimoto, Y.; Mizutani, M. The biosynthetic pathway of potato solanidanes diverged from that of spirosolanes due to evolution of a dioxygenase. Nat. Commun. 2021, 12, 1300. [Google Scholar] [CrossRef]
- Cárdenas, P.D.; Sonawane, P.D.; Pollier, J.; Vanden Bossche, R.; Dewangan, V.; Weithorn, E.; Tal, L.; Meir, S.; Rogachev, I.; Malitsky, S.; et al. GAME9 regulates the biosynthesis of steroidal alkaloids and upstream isoprenoids in the plant mevalonate pathway. Nat. Commun. 2016, 7, 10654. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Wang, P.; Tang, D.; Shang, Y.; Li, C.; Huang, S.; Zhang, C. Inheritance of steroidal glycoalkaloids in potato tuber flesh. J. Integr. Agric. 2019, 18, 2255–2263. [Google Scholar] [CrossRef]
- Abelenda, J.A.; Navarro, C.; Prat, S. From the model to the crop: Genes controlling tuber formation in potato. Curr. Opin. Biotechnol. 2011, 22, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Kloosterman, B.; Abelenda, J.A.; Gomez, M.D.M.C.; Oortwijn, M.; de Boer, J.M.; Kowitwanich, K.; Horvath, B.M.; van Eck, H.J.; Smaczniak, C.; Prat, S.; et al. Naturally occurring allele diversity allows potato cultivation in northern latitudes. Nature 2013, 495, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Abelenda, J.A.; Cruz-Oró, E.; Franco-Zorrilla, J.M.; Prat, S. Potato StCONSTANS-like1 Suppresses Storage Organ Formation by Directly Activating the FT-like StSP5G Repressor. Curr. Biol. 2016, 26, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Soyk, S.; Müller, N.A.; Park, S.J.; Schmalenbach, I.; Jiang, K.; Hayama, R.; Zhang, L.; Van Eck, J.; Jiménez-Gómez, J.M.; Lippman, Z.B. Variation in the flowering gene SELF PRUNING 5G promotes day-neutrality and early yield in tomato. Nat. Genet. 2017, 49, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.A.; Hanneman, R.E. Crossability between cultivated and wild tuber-and non-tuber-bearing Solanums. Euphytica 1999, 109, 51–67. [Google Scholar] [CrossRef]
- Ye, M.; Peng, Z.; Tang, D.; Yang, Z.; Li, D.; Xu, Y.; Zhang, C.; Huang, S. Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 2018, 4, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Enciso-Rodriguez, F.; Manrique-Carpintero, N.C.; Nadakuduti, S.S.; Buell, C.R.; Zarka, D.; Douches, D. Overcoming Self-Incompatibility in Diploid Potato Using CRISPR-Cas9. Front. Plant Sci. 2019, 10, 376. [Google Scholar] [CrossRef]
- Hartmann, A.; Senning, M.; Hedden, P.; Sonnewald, U.; Sonnewald, S. Reactivation of meristem activity and sprout growth in potato tubers require both cytokinin and gibberellin. Plant Physiol. 2011, 155, 776–796. [Google Scholar] [CrossRef] [Green Version]
- Carrera, E.; Bou, J.; Garcia-Martinez, J.L.; Prat, S. Changes in GA 20-oxidase gene expression strongly affect stem length, tuber induction and tuber yield of potato plants. Plant J. 2000, 22, 247–256. [Google Scholar] [CrossRef]
- Kloosterman, B.; Navarro, C.; Bijsterbosch, G.; Lange, T.; Prat, S.; Visser, R.G.F.; Bachem, C.W.B. StGA2ox1 is induced prior to stolon swelling and controls GA levels during potato tuber development. Plant J. 2007, 52, 362–373. [Google Scholar] [CrossRef]
- Gong, H.-L.; Dusengemungu, L.; Igiraneza, C.; Rukundo, P. Molecular regulation of potato tuber dormancy and sprouting: A mini-review. Plant Biotechnol. Rep. 2021, 15, 417–434. [Google Scholar] [CrossRef]
- Cardi, T.; Iannamico, V.; D’Ambrosio, F.; Filippone, E.; Lurquin, P.F. Agrobacterium-mediated genetic transformation of Solanum commersonii Dun. Plant Sci. 1992, 87, 179–189. [Google Scholar] [CrossRef]
- Kumar, A.; Miller, M.; Whitty, P.; Lyon, J.; Davie, P. Agrobacterium-mediated transformation of five wild Solanum species using in vitro microtubers. Plant Cell Rep. 1995, 14, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Das Dangol, S.; Yel, İ.; Çalişkan, M.E.; Bakhsh, A. Manipulating genome of diploid potato inbred line Solanum chacoense M6 using selectable marker gene. Turkish J. Agric. For. 2020, 44, 399–407. [Google Scholar] [CrossRef]
- Kumar, R.; Sharma, V.; Suresh, S.; Ramrao, D.P.; Veershetty, A.; Kumar, S.; Priscilla, K.; Hangargi, B.S.; Narasanna, R.; Pandey, M.K.; et al. Understanding Omics Driven Plant Improvement and de novo Crop Domestication: Some Examples. Front. Genet. 2021, 12, 415. [Google Scholar] [CrossRef]
- Gasparini, K.; dos Reis Moreira, J.; Peres, L.E.P.; Zsögön, A. De novo domestication of wild species to create crops with increased resilience and nutritional value. Curr. Opin. Plant Biol. 2021, 60, 102006. [Google Scholar] [CrossRef]
- Fernie, A.R.; Yan, J. De Novo Domestication: An Alternative Route toward New Crops for the Future. Mol. Plant 2019, 12, 615–631. [Google Scholar] [CrossRef] [Green Version]
- Kausch, A.P.; Nelson-Vasilchik, K.; Hague, J.; Mookkan, M.; Quemada, H.; Dellaporta, S.; Fragoso, C.; Zhang, Z.J. Edit at will: Genotype independent plant transformation in the era of advanced genomics and genome editing. Plant Sci. 2019, 281, 186–205. [Google Scholar] [CrossRef]


{kind=link}
| Species | Pathogen Resistance | Desirable Characteristics for Breeding | Undesirable Characteristics for Breeding | Reference |
|---|---|---|---|---|
| Series Bulbocastana (Rudb.) Hawkes | ||||
| S. bulbocastanum Dunal | Synchytrium endobioticum (Schilb.) Percival, Phytophthora infestans (Mont.) Pectobacterium carotovorum subsp. atrosepticum (van Hall 1902) Gardan et al., Leptinotarsa decemlineata Say, Epilacha vigintioctomaculata Motschulsky., Globodera rostochiensis Woll., Meloidogyne chitwoodi Golden, O’Bannon, Santo et Finley viruses: PVX, PVY. | High starch content in tubers (up to 37%) | Susceptible to Oospora pustulans M.N.Owen et Wakefield Bad tuberization under long-day conditions | [21,22] |
| Series Commersoniana Bukasov | ||||
| S. commersonii Dunal | S. endobioticum, Streptomyces scabies (R. Thaxter), O. pustulans, P. atrosepticum, G. rostochiensis, G. pallida Stone, L. decemlineata, E. vigintioctomaculata, viruses: PVY, PVA, PVM. | High starch content in tubers (up to 37%). Frost, heat, drought tolerant | Small tubers | [23] |
| Series Yungasensa Correll. | ||||
| S. chacoense Bitter | S. endobioticum, G. rostochiensis, G. pallida, L. decemlineata, E. vigintioctomaculata, viruses: PVY, PVA, PLRV. | No data | Susceptible to P. infestans and frost, long stolons | [23,24] |
| Series Megistacroloba Card.et Hawkes | ||||
| S. raphanifolium Cardenas et Hawkes | Verticillium albo-atrum Reinke et Berthold, V. dahlia Kleb. | Frost, drought tolerant. | Susceptible to P. infestans. Bad tuberization under long-day conditions | [23] |
| S. megistacrolobum sabtax toralapanum Cardenas et Hawkes | Viruses: PVX, PVM, PLRV. | Frost tolerant, | Susceptible to P. infestans. | [23] |
| Series Maglia Bitter | ||||
| S. maglia Schltdl. | S. endobioticum | No data | Susceptible to P. infestans. | [23] |
| Series Tuberosa (Rudb.) Hawkes (wild species) i | ||||
| S. verrucosum Schltdl. | P. infestans, Alteraria solani Ell. et Mart., L. decemlineata, E. vigintioctomaculata | High starch content | Small tubers. | [21] |
| Series Tuberosa (wild species) ii | ||||
| S. multidissectum Hawkes | S. endobioticum, P. atrosepticum, G. rostochiensis. | Frost tolerant | Susceptible to P. infestans. | [23] |
| Series Tuberosa (wild species) iii | ||||
| S. berthaultii Hawkes | P. infestans, S. endobioticumP. atrosepticum, G. rostochiensis, L. decemlineata | No data | No data | [22,23,24] |
| S. kurtzianum Bitt. et Wittm. ex Endl. | G. rostochiensis, L. decemlineata, E. vigintioctomaculata, viruses: PVX, PVY. | High starch content in tubers (up to 28%). | Susceptible to P. infestans. High SGA content | [22,23,25] |
| S. microdontum Bitter | P. infestans, S. endobioticum Meloidogyne hapla Chitwood, G. rostochiensis, G. pallida, L. decemlineata, viruses: PVY, PVA, PVS. | Drought tolerant. | Susceptible to Clavibacter michiganensis subsp. sepedonicus (Spiekermann & Kotthoff) Davis et al., black leg disease. High SGA content | [22,23,24,25] |
| S. vernei Bitter et Wittm. | P. infestans, S. endobioticum, A. solani, Rhizoctonia solani (Kühn), O. pustulans, P. atrosepticum, G. rostochiensis, L. decemlineata; viruses: PVX, PVY, PVA, PVS, PVM. | Frost tolerant (up to −4 °C). | Susceptible to High SGA content. | [22,23,26] |
| Series Acaulia Juz. | ||||
| S. acaule Bitter | S. endobioticum, S. scabies, O. pustulans, P. atrosepticum, virus PVX. | Frost tolerant | Bad tuberization under long-day conditions | [23] |
| Series Longipedicellata Buk. | ||||
| S. fendleri A. Gray ex Torrey syn. S. stoloniferum Schltdl. | P. infestans, A. solani, viruses: PVY, PVX, PLRV. | High starch content in tubers (up to 28%). | No data | [21] |
| S. polytrichon Rydberg syn. S. stoloniferum Schltdl. | P. infestans, L. decemlineata, P. atrosepticum, viruses: PVY, PVA. | Tuberization under long-day conditions | No data | [21] |
| S.× vallis-mexici Juzepczuk et Bukasov | P. infestans, S. endobioticum,P. atrosepticum | No data | Bad tuberization under long-day conditions | [21] |
| Series Demissa Bukasov. | ||||
| S. demissum Lindley | P. infestans, S. scabies | Frost, heat, drought tolerant. High starch content in tubers (up to 33%). | Small tubers | [21] |
| Species | Pathogen Resistance | Mapped R-Loci or Identified Genes | Reference |
|---|---|---|---|
| Series Bulbocastana | |||
| S. bulbocastanum Dunal | Phytophthora infestans (Mont.) | Rpi-blb1 | [34] |
| Rpi-blb2 | [35] | ||
| Rpi-blb3 | [36] | ||
| Rpi-bt1 | [37] | ||
| Meloidogyne chitwoodi Golden, O’Bannon, Santo et Finley | RMc1blb | [38] | |
| Series Pinnatisecta (Rudb.) Hawkes | |||
| S. brachistotrichum (Bitter) Rydb | P. infestans | Rpi-bst1 | [39] |
| S.× michoacanum (Bitter) Rydb. | P. infestans | Rpi-mch1 | [40] |
| S. pinnatisectum Dunal | P. infestans | Rpi-pnt1 | [41] |
| Series Circaeifolia Hawkes | |||
| S. capsicibaccatum Cardenas | P. infestans | Rpi-cap1 | [42] |
| Series Yungasensa | |||
| S. chacoense Bitter | P. infestans | Rpi-chc1 | [43] |
| Virus PVY | Ry chc | [44] | |
| Viruses: PVY, PVX | Ny chc, Nx chc | [45] | |
| Series Megistacroloba | |||
| S. megistacrolobum Bitter | Virus PVM | Rm | [46] |
| Series Piurana Ochoa(Piu) | |||
| S. paucissectum Bitter | P. infestans | QTLpcs10, QTLpcs11 QTLpcs12 | [47] |
| Series Tuberosa (wild species) ii | |||
| S. mochiquense Ochoa | P. infestans | Rpi-moc1 | [48] |
| S. sparsipilum Bitter | P. infestans | Pi_QTLspr-1, Pi_QTLspr-2 | [49] |
| Virus PVYc | Nc spl | [50] | |
| Globoderapallida Stone Pa2, Pa3 | Gpa Vsspl, Gpa XIsspl | [51] | |
| Series Tuberosa (wild species) iii | |||
| S. berthaultii Hawkes | P. infestans | Rpi-ber1 Rpi-ber2 | [52] |
| S. microdontum Bitter | P. infestans | Rpi-mcd1 | [53] |
| S. spegazzinii Bitter | P. infestans | Pi_QTLspg | [49] |
| G. rostochiensis Woll. Ro1-Ro5 | Gro1, | [54] | |
| G. rostochiensis Ro1 | Gro1-4 | [55] | |
| G. rostochiensis Ro1 G. pallida Pa2, Pa3 | Gro1.2, Gro1.3, Gro1.4, Gpa3 | [56] | |
| G. pallida Pa2, Pa3 | GpaM1, GpaM2, GpaM3 | [57] | |
| S. venturii Hawkes et Hjerting | P. infestans | Rpi-vnt1.1 | [58] |
| S. vernei Bitter et Wittm. | Virus PVX | Rx vrn | [59] |
| G. rostochiensis Ro1 | GroV1 | [60] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, A.A.; Chalaya, N.A.; Fomin, I.N.; Barchuk, A.I.; Gerasimova, S.V. De Novo Domestication Concept for Potato Germplasm Enhancement. Agronomy 2022, 12, 462. https://doi.org/10.3390/agronomy12020462
Egorova AA, Chalaya NA, Fomin IN, Barchuk AI, Gerasimova SV. De Novo Domestication Concept for Potato Germplasm Enhancement. Agronomy. 2022; 12(2):462. https://doi.org/10.3390/agronomy12020462
Chicago/Turabian StyleEgorova, Anastasiya A., Nadezhda A. Chalaya, Ivan N. Fomin, Alisa I. Barchuk, and Sophia V. Gerasimova. 2022. "De Novo Domestication Concept for Potato Germplasm Enhancement" Agronomy 12, no. 2: 462. https://doi.org/10.3390/agronomy12020462