
NLR Genes Related Transcript Sets in Potato Cultivars Bearing Genetic Material of Wild Mexican Solanum Species

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Resistance Assessment
2.3. RNA-seq
2.4. RT-qPCR
2.5. Bioinformatic Analysis
2.5.1. Read Quality Assessment, Filtering and Mapping
2.5.2. Transcriptome Assembly De Novo
2.5.3. Transcriptome Analysis
2.5.4. Compilation of the Marker Genes Associated with Wide-Range Resistance to P. infestans
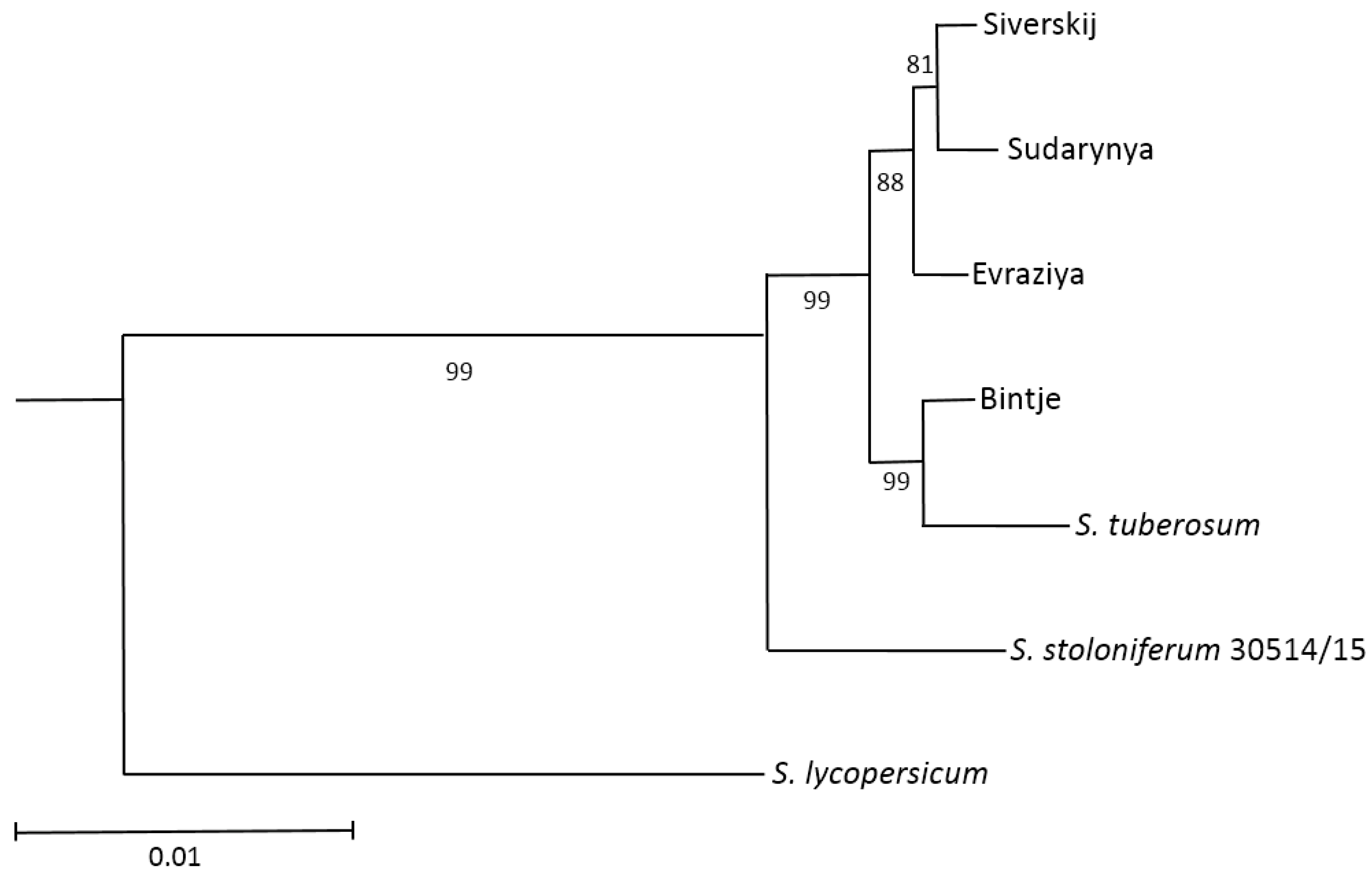
2.5.5. Phylogenetic Analysis of the Novel NBS-LRR Sequences
3. Results
3.1. Assessment of Resistance to P. infestans
3.2. Libraries Comparison and Quality Evaluation
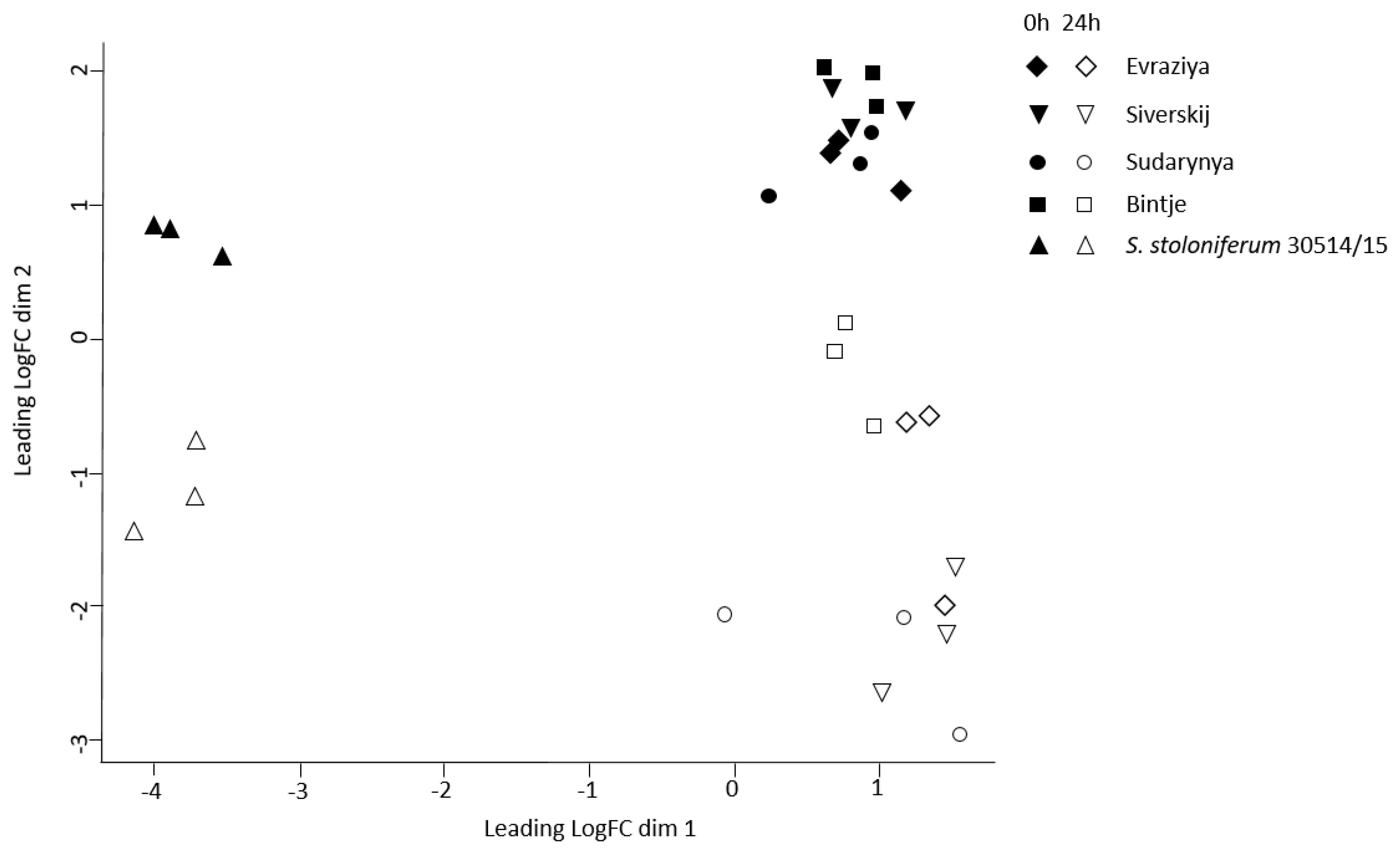
3.3. De novo Transcriptome Assembly and Reconstruction of the Orthologous Groups
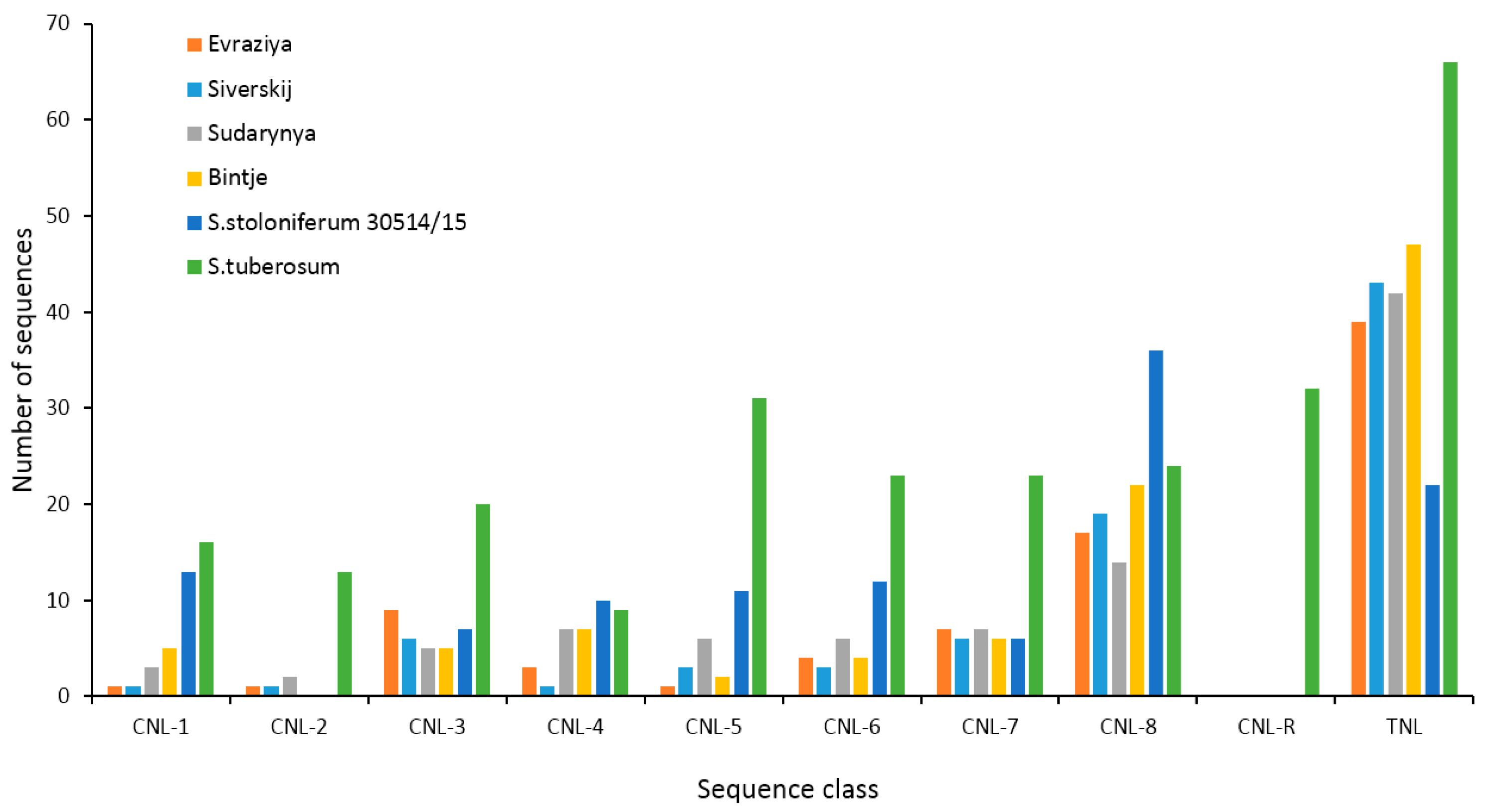
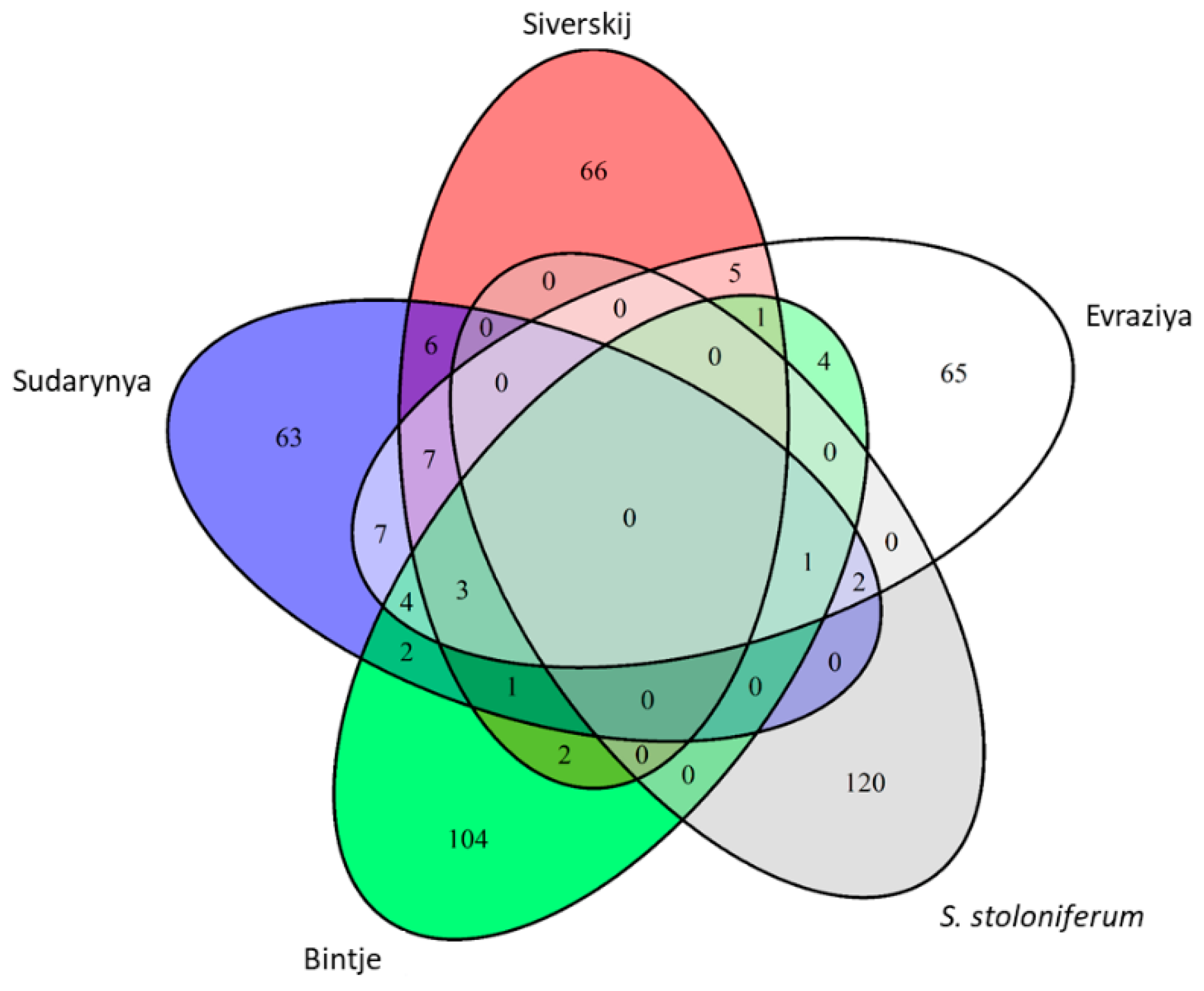
3.4. Identification and Analysis of the NBS-LRR Proteins
3.5. Novel Transcripts Encoding Proteins with NBS-LRR Domains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caruana, B.; Rodoni, B.; Constable, F.; Slater, A.; Cogan, N. Genome Enhanced Marker Improvement for Potato Virus Y Disease Resistance in Potato. Agronomy 2021, 11, 832. [Google Scholar] [CrossRef]
- Stefańczyk, E.; Plich, J.; Janiszewska, M.; Smyda-Dajmund, P.; Sobkowiak, S.; Śliwka, J. Marker-assisted pyramiding of potato late blight resistance genes Rpi-rzc1 and Rpi-phu1 on di- and tetraploid levels. Mol. Breed. 2020, 40, 1–12. [Google Scholar] [CrossRef]
- Rakosy-Tican, E.; Thieme, R.; König, J.; Nachtigall, M.; Hammann, T.; Denes, T.-E.; Kruppa, K.; Molnár-Láng, M. Introgression of Two Broad-Spectrum Late Blight Resistance Genes, Rpi-Blb1 and Rpi-Blb3, From Solanum bulbocastanum Dun Plus Race-Specific R Genes Into Potato Pre-breeding Lines. Front. Plant Sci. 2020, 11, 699. [Google Scholar] [CrossRef]
- Shao, Z.-Q.; Wang, B.; Chen, J.-Q. Tracking ancestral lineages and recent expansions of NBS-LRR genes in angiosperms. Plant Signal. Behav. 2016, 11, e1197470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, R.; Ponce, O.P.; Ramírez, M.; Mostajo, N.; Orjeda, G. Genome-Wide Identification and Mapping of NBS-Encoding Resistance Genes in Solanum tuberosum Group Phureja. PLoS ONE 2012, 7, e34775. [Google Scholar] [CrossRef] [PubMed]
- Sekhwal, M.K.; Li, P.; Lam, I.; Wang, X.; Cloutier, S.; You, F.M. Disease Resistance Gene Analogs (RGAs) in Plants. Int. J. Mol. Sci. 2015, 16, 19248–19290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Weng, Q.; Song, J.; Ma, H.; Yuan, J.; Dong, Z.; Liu, Y. Bioinformatics Analysis of NBS-LRR Encoding Resistance Genes in Setaria italica. Biochem. Genet. 2016, 54, 232–248. [Google Scholar] [CrossRef]
- Pal, T.; Jaiswal, V.; Chauhan, R.S. DRPPP: A machine learning based tool for prediction of disease resistance proteins in plants. Comput. Biol. Med. 2016, 78, 42–48. [Google Scholar] [CrossRef]
- Toda, N.; Rustenholz, C.; Baud, A.; Le Paslier, M.-C.; Amselem, J.; Merdinoglu, D.; Faivre-Rampant, P. NLGenomeSweeper: A Tool for Genome-Wide NBS-LRR Resistance Gene Identification. Genes 2020, 11, 333. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ghouri, F.; Yu, H.; Li, X.; Yu, S.; Shahid, M.Q.; Liu, X. Genome wide re-sequencing of newly developed Rice Lines from common wild rice (Oryza rufipogon Griff.) for the identification of NBS-LRR genes. PLoS ONE 2017, 12, e0180662. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Silverstein, K.A.T.; Ramaraj, T.; Guhlin, J.; Denny, R.; Liu, J.; Farmer, A.D.; Steele, K.P.; Stupar, R.M.; Miller, J.R.; et al. Exploring structural variation and gene family architecture with De Novo assemblies of 15 Medicago genomes. BMC Genom. 2017, 18, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Jiang, X.-M.; Shao, Z.-Q. Genome-Wide Analysis of NLR Disease Resistance Genes in an Updated Reference Genome of Barley. Front. Genet. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Jupe, F.; Witek, K.; Verweij, W.; Śliwka, J.; Pritchard, L.; Etherington, G.J.; Maclean, D.; Cock, P.J.; Leggett, R.M.; Bryan, G.J.; et al. Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J. 2013, 76, 530–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirin, E.A.; Mann, H.; Meyer, R.S.; Traini, A.; Chiusano, M.L.; Litt, A.; Bradeen, J. Evolutionary Meta-Analysis of Solanaceous Resistance Gene and Solanum Resistance Gene Analog Sequences and a Practical Framework for Cross-Species Comparisons. Mol. Plant-Microbe Interact. 2012, 25, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Morata, J.; Puigdomènech, P. Variability among Cucurbitaceae species (melon, cucumber and watermelon) in a genomic region containing a cluster of NBS-LRR genes. BMC Genom. 2017, 18, 138. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Gan, L.; Shang, K.; Wang, C.; Song, Y.; Liu, H.; Zhou, S.; Zhu, C. Global transcriptome analyses reveal the molecular signatures in the early response of potato (Solanum tuberosum L.) to Phytophthora infestans, Ralstonia solanacearum, and Potato virus Y infection. Planta 2020, 252, 1–13. [Google Scholar] [CrossRef]
- Kochetov, A.V.; Glagoleva, A.Y.; Strygina, K.V.; Khlestkina, E.K.; Gerasimova, S.; Ibragimova, S.M.; Shatskaya, N.V.; Vasilyev, G.V.; Afonnikov, D.A.; Shmakov, N.A.; et al. Differential expression of NBS-LRR-encoding genes in the root transcriptomes of two Solanum phureja genotypes with contrasting resistance to Globodera rostochiensis. BMC Plant Biol. 2017, 17, 251. [Google Scholar] [CrossRef]
- Kochetov, A.V.; Egorova, A.A.; Glagoleva, A.Y.; Strygina, K.V.; Khlestkina, E.K.; Gerasimova, S.V.; Shatskaya, N.V.; Vasilyev, G.V.; Afonnikov, D.A.; Shmakov, N.A.; et al. The mechanism of potato resistance to Globodera rostochiensis: Comparison of root transcriptomes of resistant and susceptible Solanum phureja genotypes. BMC Plant Biol. 2020, 20, 350. [Google Scholar] [CrossRef]
- Sagi, M.S.; Deokar, A.A.; Tar’An, B. Genetic Analysis of NBS-LRR Gene Family in Chickpea and Their Expression Profiles in Response to Ascochyta Blight Infection. Front. Plant Sci. 2017, 8, 838. [Google Scholar] [CrossRef] [Green Version]
- Vleeshouwers, V.G.A.A.; Oliver, R.P. Effectors as Tools in Disease Resistance Breeding Against Biotrophic, Hemibiotrophic, and Necrotrophic Plant Pathogens. Mol. Plant-Microbe Interact. 2015, 2015, 17–27. [Google Scholar] [CrossRef]
- Brendolise, C.; Montefiori, M.; Dinis, R.; Peeters, N.; Storey, R.D.; Rikkerink, E.H. A novel hairpin library-based approach to identify NBS–LRR genes required for effector-triggered hypersensitive response in Nicotiana benthamiana. Plant Methods 2017, 13, 32. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.-T.; Choi, H.; Kim, S.-B.; Lee, H.-A.; Choi, D.; Kim, K.-H. A simple method for screening of plant NBS-LRR genes that confer a hypersensitive response to plant viruses and its application for screening candidate pepper genes against Pepper mottle virus. J. Virol. Methods 2014, 201, 57–64. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, H.; Cai, T.; Deng, Y.; Zhuang, R.; Zhang, N.; Zeng, Y.; Zheng, Y.; Tang, R.; Pan, R.; et al. Overexpression of a novel peanut NBS-LRR geneAhRRS5enhances disease resistance toRalstonia solanacearumin tobacco. Plant Biotechnol. J. 2017, 15, 39–55. [Google Scholar] [CrossRef]
- Wei, C.; Chen, J.; Kuang, H. Dramatic Number Variation of R Genes in Solanaceae Species Accounted for by a Few R Gene Subfamilies. PLoS ONE 2016, 11, e0148708. [Google Scholar] [CrossRef]
- Qian, L.-H.; Zhou, G.-C.; Sun, X.-Q.; Lei, Z.; Zhang, Y.-M.; Xue, J.-Y.; Hang, Y.-Y. Distinct Patterns of Gene Gain and Loss: Diverse Evolutionary Modes of NBS-Encoding Genes in Three Solanaceae Crop Species. G3 Genes Genomes Genet. 2017, 7, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Chae, E.; Bomblies, K.; Kim, S.-T.; Karelina, D.; Zaidem, M.; Ossowski, S.; Martín-Pizarro, C.; Laitinen, R.A.; Rowan, B.A.; Tenenboim, H.; et al. Species-wide Genetic Incompatibility Analysis Identifies Immune Genes as Hot Spots of Deleterious Epistasis. Cell 2014, 159, 1341–1351. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Xue, S.; Lei, L.; Fan, M.; Peng, S.; Li, T.; Nagarajan, R.; Carver, B.; Ma, Z.; Deng, J.; et al. A semi-dominant NLR allele causes whole-seedling necrosis in wheat. Plant Physiol. 2021, 186, 483–496. [Google Scholar] [CrossRef]
- Barragan, A.C.; Collenberg, M.; Wang, J.; Lee, R.R.Q.; Cher, W.Y.; Rabanal, F.A.; Ashkenazy, H.; Weigel, D.; Chae, E. A Truncated Singleton NLR Causes Hybrid Necrosis in Arabidopsis thaliana. Mol. Biol. Evol. 2021, 38, 557–574. [Google Scholar] [CrossRef]
- Ma, J.; Hancock, W.G.; Nifong, J.M.; Kernodle, S.P.; Lewis, R.S. Identification and editing of a hybrid lethality gene expands the range of interspecific hybridization potential in Nicotiana. Theor. Appl. Genet. 2020, 133, 2915–2925. [Google Scholar] [CrossRef]
- Deng, J.; Fang, L.; Zhu, X.; Zhou, B.; Zhang, T. A CC-NBS-LRR gene induces hybrid lethality in cotton. J. Exp. Bot. 2019, 70, 5145–5156. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Hou, X.; Yang, G.; Xiao, Y.; Han, L.; Meng, J.; Luan, Y. Sl-lncRNA15492 interacts with Sl-miR482a and affects Solanum lycopersicum immunity against Phytophthora infestans. Plant J. 2020, 103, 1561–1574. [Google Scholar] [CrossRef]
- Jiang, N.; Cui, J.; Shi, Y.; Yang, G.; Zhou, X.; Hou, X.; Meng, J.; Luan, Y. Tomato lncRNA23468 functions as a competing endogenous RNA to modulate NBS-LRR genes by decoying miR482b in the tomato-Phytophthora infestans interaction. Hortic. Res. 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.-H.; Meng, J.; Zhang, M.; Luan, Y.-S. Identification of tomato circular RNAs responsive to Phytophthora infestans. Gene 2020, 746, 144652. [Google Scholar] [CrossRef]
- Parker, M.T.; Knop, K.; Zacharaki, V.; Sherwood, A.V.; Tome, D.; Yu, X.; Martin, P.G.; Beynon, J.; Michaels, S.; Barton, G.J.; et al. Widespread premature transcription termination of Arabidopsis thaliana NLR genes by the spen protein FPA. eLife 2021, 10, 65537. [Google Scholar] [CrossRef]
- Cantalapiedra, C.; Contreras-Moreira, B.; Silvar, C.; Perovic, D.; Ordon, F.; Gracia, M.P.; Igartua, E.; Casas, A.M. A Cluster of Nucleotide-Binding Site–Leucine-Rich Repeat Genes Resides in a Barley Powdery Mildew Resistance Quantitative Trait Loci on 7HL. Plant Genome 2016, 9. [Google Scholar] [CrossRef]
- Lee, R.R.; Chae, E. Variation Patterns of NLR Clusters in Arabidopsis thaliana Genomes. Plant Commun. 2020, 1, 100089. [Google Scholar] [CrossRef]
- Göktay, M.; Fulgione, A.; Hancock, A.M. A New Catalog of Structural Variants in 1301 A. thaliana Lines from Africa, Eurasia, and North America Reveals a Signature of Balancing Selection at Defense Response Genes. Mol. Biol. Evol. 2021, 38, 1498–1511. [Google Scholar] [CrossRef]
- Prakash, C.; Trognitz, F.C.; Venhuizen, P.; Von Haeseler, A.; Trognitz, B. A compendium of genome-wide sequence reads from NBS (nucleotide binding site) domains of resistance genes in the common potato. Sci. Rep. 2020, 10, 1–21. [Google Scholar] [CrossRef]
- Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Gavrilenko, T.A.; Klimenko, N.S.; Antonova, O.Y.; Lebedeva, V.A.; Evdokimova, Z.Z.; Gadjiyev, N.M.; Apalikova, O.V.; Alpatyeva, N.V.; Kostina, L.I.; Zoteyeva, N.M.; et al. Molecular screening of potato varieties bred in the northwestern zone of the Russian Federation. Vavilovskii Zhurnal Genet. I Sel. Vavilov J. Genet. Breed. 2018, 22, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Antonova, O.Y.; Klimenko, N.S.; Evdokimova, Z.Z.; Kostina, L.I.; Gavrilenko, T.A. Finding RB/Rpi-blb1/Rpi-sto1-like sequences in conventionally bred potato varieties. Vavilov J. Genet. Breed. 2018, 22, 693–702. [Google Scholar] [CrossRef]
- Naess, S.K.; Bradeen, J.M.; Wielgus, S.M.; Haberlach, G.T.; McGrath, J.M.; Helgeson, J.P. Resistance to late blight in Solanum bulbocastanum is mapped to chromosome 8. Theor. Appl. Genet. 2000, 101, 697–704. [Google Scholar] [CrossRef]
- Song, J.; Bradeen, J.M.; Naess, S.K.; Raasch, J.A.; Wielgus, S.M.; Haberlach, G.T.; Liu, J.; Kuang, H.; Austin-Phillips, S.; Buell, C.R.; et al. Gene RB cloned from Solanum bulbocastanum confers broad spectrum resistance to po-tato late blight. Proc. Natl. Acad. Sci. USA 2003, 100, 9128–9133. [Google Scholar] [CrossRef] [Green Version]
- Van Der Vossen, E.; Sikkema, A.; Hekkert, B.T.L.; Gros, J.; Stevens, P.; Muskens, M.; Wouters, D.; Pereira, A.; Stiekema, W.; Allefs, S. An ancientRgene from the wild potato speciesSolanum bulbocastanumconfers broad-spectrum resistance toPhytophthora infestansin cultivated potato and tomato. Plant J. 2003, 36, 867–882. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.A.A.; Rietman, H.; Krenek, P.; Champouret, N.; Young, C.; Oh, S.-K.; Wang, M.; Bouwmeester, K.; Vosman, B.; Visser, R.G.F.; et al. Effector Genomics Accelerates Discovery and Functional Profiling of Potato Disease Resistance and Phytophthora Infestans Avirulence Genes. PLoS ONE 2008, 3, e2875. [Google Scholar] [CrossRef]
- Wang, M.; Allefs, S.; Berg, R.G.V.D.; Vleeshouwers, V.G.A.A.; van der Vossen, E.A.G.; Vosman, B. Allele mining in Solanum: Conserved homologues of Rpi-blb1 are identified in Solanum stoloniferum. Theor. Appl. Genet. 2008, 116, 933–943. [Google Scholar] [CrossRef]
- Lokossou, A.A.; Rietman, H.; Wang, M.; Krenek, P.; van der Schoot, H.; Henken, B.; Hoekstra, R.; Vleeshouwers, V.G.A.A.; van der Vossen, E.A.G.; Visser, R.G.F.; et al. Diversity, Distribution, and Evolution ofSolanum bulbocastanumLate Blight Resistance Genes. Mol. Plant-Microbe Interact. 2010, 23, 1206–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Vossen, J.H.; Visser, R.G.F. Durable Late Blight Resistance in Potato Through Dynamic Varieties Obtained by Cisgenesis: Scientific and Societal Advances in the DuRPh Project. Potato Res. 2016, 59, 35–66. [Google Scholar] [CrossRef] [Green Version]
- Khiutti, A.; Spooner, D.M.; Jansky, S.H.; Halterman, D.A. Testing Taxonomic Predictivity of Foliar and Tuber Resistance to Phytophthora infestans in Wild Relatives of Potato. Phytopathology 2015, 105, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Cruickshank, G.; Stewart, H.E.; Wastie, R.L. An illustrated assessment key for foliage blight of potatoes. Potato Res. 1982, 25, 213–214. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Lin, H.-N.; Hsu, W.-L. DART: A fast and accurate RNA-seq mapper with a partitioning strategy. Bioinformatics 2017, 34, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.L.; Contreras-Moreira, B.; De Silva, N.; Maslen, G.; Akanni, W.; Allen, J.; Alvarez-Jarreta, J.; Barba, M.; Bolser, D.M.; Cambell, L.; et al. Ensembl Genomes 2020—enabling non-vertebrate genomic research. Nucleic Acids Res. 2020, 48, D689–D695. [Google Scholar] [CrossRef] [Green Version]
- Hardigan, M.A.; Laimbeer, F.P.E.; Newton, L.; Crisovan, E.; Hamilton, J.; Vaillancourt, B.; Wiegert-Rininger, K.; Wood, J.; Douches, D.S.; Farré, E.M.; et al. Genome diversity of tuber-bearing Solanum uncovers complex evolutionary history and targets of domestication in the cultivated potato. Proc. Natl. Acad. Sci. USA 2017, 114, E9999–E10008. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Roulin, A.C.; Wu, M.; Pichon, S.; Arbore, R.; Kühn-Bühlmann, S.; Kölliker, M.; Walser, J.-C. De Novo Transcriptome Hybrid Assembly and Validation in the European Earwig (Dermaptera, Forficula auricularia). PLoS ONE 2014, 9, e94098. [Google Scholar] [CrossRef]
- Shmakov, N.A. Improving the quality of barley transcriptome de novo assembling by using a hybrid approach for lines with varying spike and stem coloration. Vavilov J. Genet. Breed. 2021, 25, 30–38. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Robertson, G.; Schein, J.; Chiu, R.; Corbett, R.; Field, M.; Jackman, S.D.; Mungall, K.; Lee, S.; Okada, H.M.; Qian, J.Q.; et al. De novo assembly and analysis of RNA-seq data. Nat. Methods 2010, 7, 909–912. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.D.; Watanabe, C.K. GMAP: A genomic mapping and alignment program for mRNA and EST sequences. Bioinformatics 2005, 21, 1859–1875. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic orthology inference for comparative genomics. Genome Biol. 2019, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From reads to genes to pathways: Differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [CrossRef] [Green Version]
- Steuernagel, B.; Jupe, F.; Witek, K.; Jones, J.D.; Wulff, B.B. NLR-parser: Rapid annotation of plant NLR complements. Bioinformatics 2015, 31, 1665–1667. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A Cross-Platform and Ultrafast Toolkit for FASTA/Q File Manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef] [PubMed]
- Pronozin, A.Y.; Bragina, M.K.; Salina, E.A. Crop pangenomes. Vavilov J. Genet. Breed. 2021, 25, 57–63. [Google Scholar] [CrossRef]
- Golicz, A.; Batley, J.; Edwards, D. Towards plant pangenomics. Plant Biotechnol. J. 2015, 14, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhou, Y.; Ye, G.; Zheng, J.; Meng, Y.; Wang, J.; Shan, W. Serial Transcriptome Analysis Reveals Genes Associated with Late Blight Resistance in Potato Cultivar Qingshu. Agronomy 2021, 11, 1919. [Google Scholar] [CrossRef]
- Bradeen, J.M.; Iorizzo, M.; Mollov, D.S.; Raasch, J.; Kramer, L.C.; Millett, B.P.; Austin-Phillips, S.; Jiang, J.; Carputo, D. Higher Copy Numbers of the PotatoRBTransgene Correspond to Enhanced Transcript and Late Blight Resistance Levels. Mol. Plant-Microbe Interact. 2009, 22, 437–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, L.C.; Choudoir, M.J.; Wielgus, S.M.; Bhaskar, P.B.; Jiang, J. Correlation Between Transcript Abundance of the RB Gene and the Level of the RB-Mediated Late Blight Resistance in Potato. Mol. Plant-Microbe Interact. 2009, 22, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochetov, A.V.; Titov, S.; Kolodyazhnaya, Y.S.; Komarova, M.L.; Koval, V.S.; Makarova, N.N.; Il’Yinskyi, Y.Y.; Trifonova, E.A.; Shumny, V. Tobacco Transformants Bearing Antisense Suppressor of Proline Dehydrogenase Gene, Are Characterized by Higher Proline Content and Cytoplasm Osmotic Pressure. Russ. J. Genet. 2004, 40, 216–218. [Google Scholar] [CrossRef]
- Trifonova, E.A.; Sapotsky, M.V.; Komarova, M.L.; Scherban, A.B.; Shumny, V.K.; Polyakova, A.M.; Lapshina, L.A.; Kochetov, A.V.; Malinovsky, V.I. Protection of transgenic tobacco plants expressing bovine pancreatic ribonuclease against tobacco mosaic virus. Plant Cell Rep. 2007, 26, 1121–1126. [Google Scholar] [CrossRef]
- Sugawara, T.; Trifonova, E.A.; Kochetov, A.V.; Kanayama, Y. Expression of an extracellular ribonuclease gene increases resistance to Cucumber mosaic virus in tobacco. BMC Plant Biol. 2016, 16, 246. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Y.; Yin, L.; Lu, J. Overexpression of pathogen-induced grapevine TIR-NB-LRR gene VaRGA1 enhances disease resistance and drought and salt tolerance in Nicotiana benthamiana. Protoplasma 2017, 254, 957–969. [Google Scholar] [CrossRef]
- Ghislain, M.; Byarugaba, A.A.; Magembe, E.; Njoroge, A.; Rivera, C.; Román, M.L.; Tovar, J.C.; Gamboa, S.; Forbes, G.A.; Kreuze, J.; et al. Stacking three late blight resistance genes from wild species directly into African highland potato varieties confers complete field resistance to local blight races. Plant Biotechnol. J. 2018, 17, 1119–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, J.G.; Gaviria, A.E.; Gilchrist, E. Allelic Variation and Selection in Effector Genes of Phytophthora infestans (Mont.) de Bary. Pathogens 2020, 9, 551. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, X.; Han, T.; Li, K.; Qu, Y.; Gao, Z. Differential Potential of Phytophthora capsici Resistance Mechanisms to the Fungicide Metalaxyl in Peppers. Microorganisms 2020, 8, 278. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Duan, S.; Armstrong, M.R.; Duan, Y.; Xu, J.; Chen, X.; Hein, I.; Jin, L.; Li, G. New Findings on the Resistance Mechanism of an Elite Diploid Wild Potato Species JAM1-4 in Response to a Super Race Strain of Phytophthora infestans. Phytopathology 2020, 110, 1375–1387. [Google Scholar] [CrossRef]
- Frolova, T.S.; Cherenko, V.A.; Sinitsyna, O.I.; Kochetov, A.V. Genetic aspects of potato resistance to phytophthorosis. Vavilov J. Genet. Breed. 2021, 25, 164–170. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potato Cultivar | Disease Assessment (9-Point Scale) * | |||||
|---|---|---|---|---|---|---|
| Replicate 1 | Replicate 2 | |||||
| Days after Inoculation | ||||||
| 7 | 10 | 13 | 7 | 10 | 13 | |
| Sudarynya | 8.2 ± 0.23 | 8.0 ± 0.23 | 8.0 ± 0.22 | 9.0 ± 0.00 | 6.5 ± 0.09 | 7.0 ± 0.08 |
| Siverskij | 6.9 ± 0.15 | 6.7 ± 0.13 | 6.2 ± 0.13 | 8.0 ± 0.00 | 7.0 ± 0.20 | 4.7 ± 0.09 |
| Evraziya | 4.8 ± 0.27 | 4.2 ± 0.19 | 1.8 ± 0.11 | 3.4 ± 0.17 | 2.1 ± 0.17 | 1.1 ± 0.05 |
| Bintje (susceptible control) | 4.5 ± 0.15 | 2.5 ± 0.14 | 1.0 ± 0.00 | 1.7 ± 0.14 | 1.0 ± 0.00 | 1.0 ± 0.00 |
| S. stoloniferum, clone 30514/15 (resistant control) | 9.0 ± 0.00 | 9.0 ± 0.00 | 9.0 ± 0.00 | - | - | - |
| Potato Genotype | Number of Proteins with NBS-LRR Domains | Number of Proteins with Complete NBS-LRR Domains | Number of Proteins with Complete NBS-LRR Domains and TPM > 1 |
|---|---|---|---|
| Siverskij | 1188 | 409 | 358 (88%) |
| Evraziya | 1331 | 449 | 376 (84%) |
| Sudarynya | 960 | 334 | 310 (93%) |
| Bintje | 1185 | 444 | 384 (86%) |
| S. stoloniferum, 30514/15 | 1333 | 504 | 417 (83%) |
| Genotype | Number of Transcripts | Differential Expression (FDR < 0.05) | |||
|---|---|---|---|---|---|
| Total | CNL | TNL | Upregulated | Downregulated | |
| Evraziya | 104 | 66 | 38 | 3 | 1 |
| Siverskij | 103 | 67 | 36 | 7 | 5 |
| Sudarynya | 109 | 70 | 39 | 11 | 3 |
| Bintje | 133 | 94 | 39 | 7 | 9 |
| S. stoloniferum 30514/15 | 160 | 138 | 22 | 41 | 16 |
| Potato Cultivars | Transcript 1 | Homolog/Identity | Abundance at 0 hpi, TPM | Abundance at 24 hpi, TPM | Log2(FC) 0 vs. 24 hpi | FDR |
|---|---|---|---|---|---|---|
| Siverskij | R450380 | R3b/99.9% | 7.42 | 5.82 | −0.52 | 0.21 |
| R488392 | Rpi-blb1/99.8% | 2.04 | 7.43 | 2.14 | 0.004 | |
| Evraziya | R595896 | R3b/99.9% | 5.66 | 5.75 | 0.49 | 0.55 |
| Sudarynya | R489301 | R3b/99.8% | 2.52 | 3.05 | 0.09 | 0.95 |
| R443838 | Rpi-blb1/99.8% | 0.90 | 5.43 | 4.21 | 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kochetov, A.V.; Afonnikov, D.A.; Shmakov, N.; Vasiliev, G.V.; Antonova, O.Y.; Shatskaya, N.V.; Glagoleva, A.Y.; Ibragimova, S.M.; Khiutti, A.; Afanasenko, O.S.; et al. NLR Genes Related Transcript Sets in Potato Cultivars Bearing Genetic Material of Wild Mexican Solanum Species. Agronomy 2021, 11, 2426. https://doi.org/10.3390/agronomy11122426
Kochetov AV, Afonnikov DA, Shmakov N, Vasiliev GV, Antonova OY, Shatskaya NV, Glagoleva AY, Ibragimova SM, Khiutti A, Afanasenko OS, et al. NLR Genes Related Transcript Sets in Potato Cultivars Bearing Genetic Material of Wild Mexican Solanum Species. Agronomy. 2021; 11(12):2426. https://doi.org/10.3390/agronomy11122426
Chicago/Turabian StyleKochetov, Alex V., Dmitry A. Afonnikov, Nikolay Shmakov, Gennady V. Vasiliev, Olga Y. Antonova, Natalja V. Shatskaya, Anastasiya Y. Glagoleva, Salmaz M. Ibragimova, Aleksander Khiutti, Olga S. Afanasenko, and et al. 2021. "NLR Genes Related Transcript Sets in Potato Cultivars Bearing Genetic Material of Wild Mexican Solanum Species" Agronomy 11, no. 12: 2426. https://doi.org/10.3390/agronomy11122426