
Metabolomics Analysis Reveals Dynamic Accumulation of Sugar and Acid during Stem Development of Brassica juncea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Methods for Determination of Morphological Indexes, Sugar and Acid Contents, and Related Enzymes
2.3. Metabolomics Analysis
2.3.1. Sample Preparation and UHPLC-MS/MS
2.3.2. Data Processing
2.3.3. Identification and Screening of Metabolites
2.4. Validation of Gene Expression Levels
3. Results
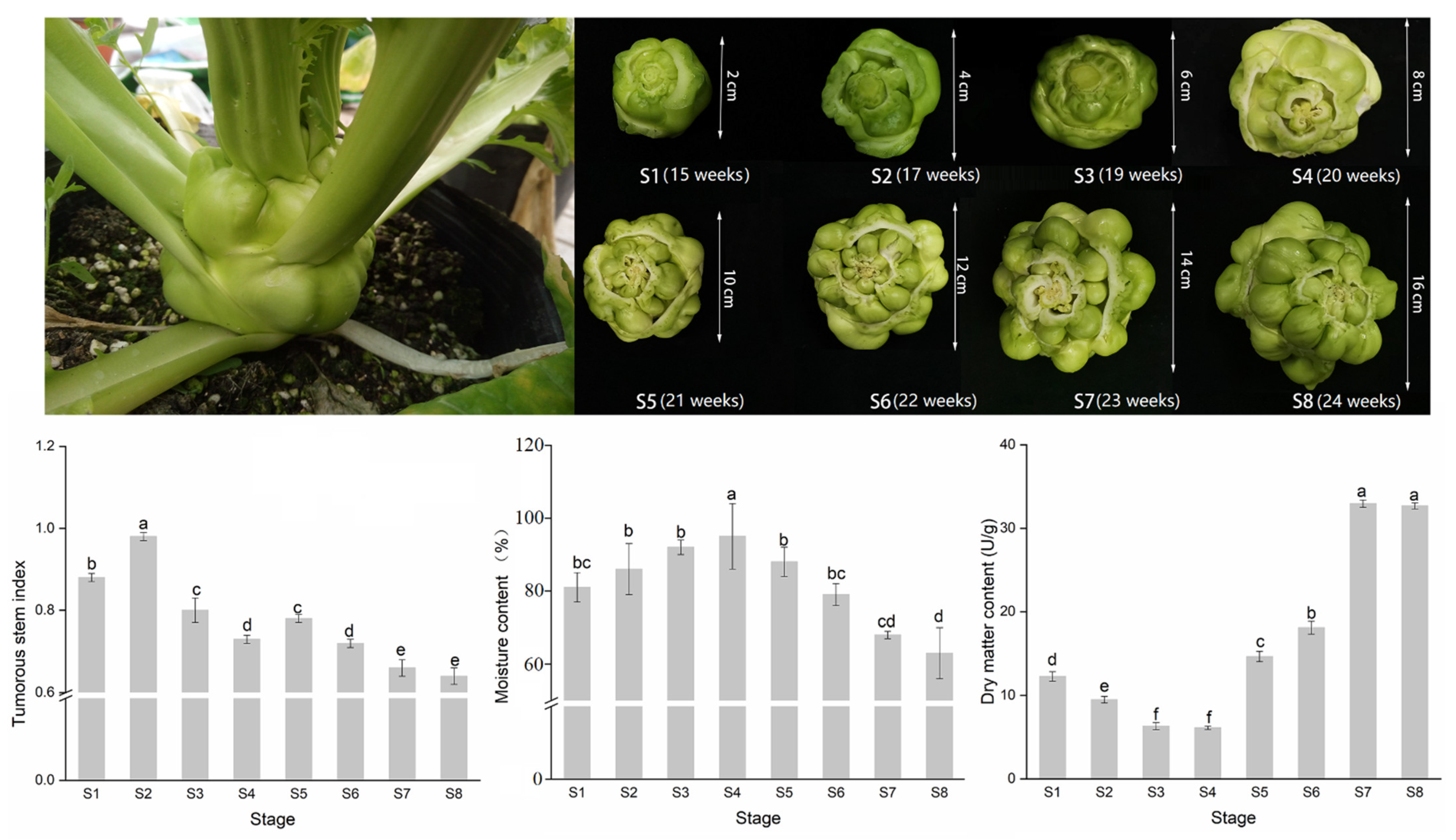
3.1. Morphological and Growth Index Changes during Stem Expansion of Stem Mustard
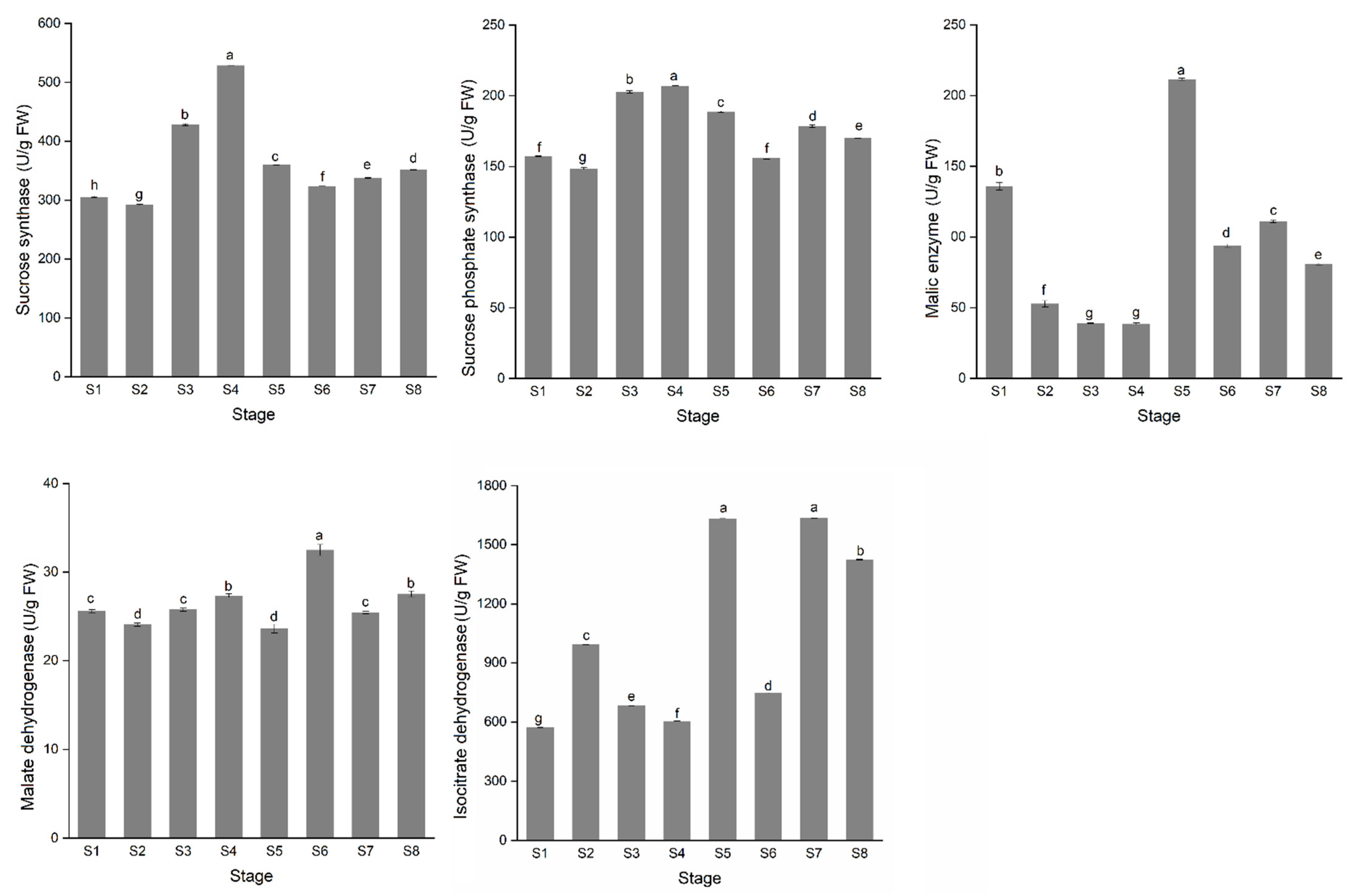
3.2. The Variation of Sugar and Acid Content during Stem Development
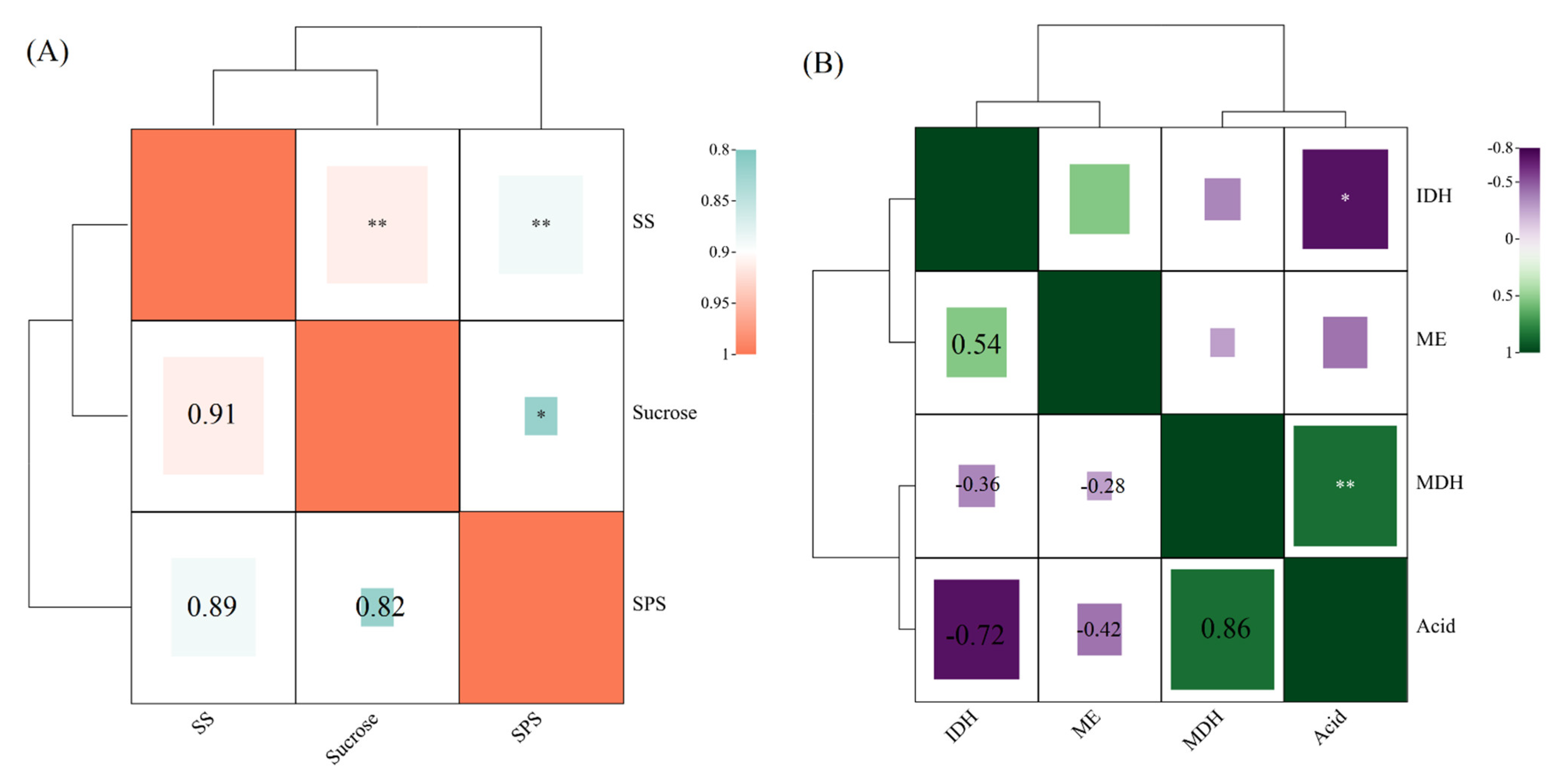
3.3. Analysis of the Correlation between Sugar and Acid Metabolizing Enzymes during the Development of Stem
3.4. Analysis of Metabolite Levels
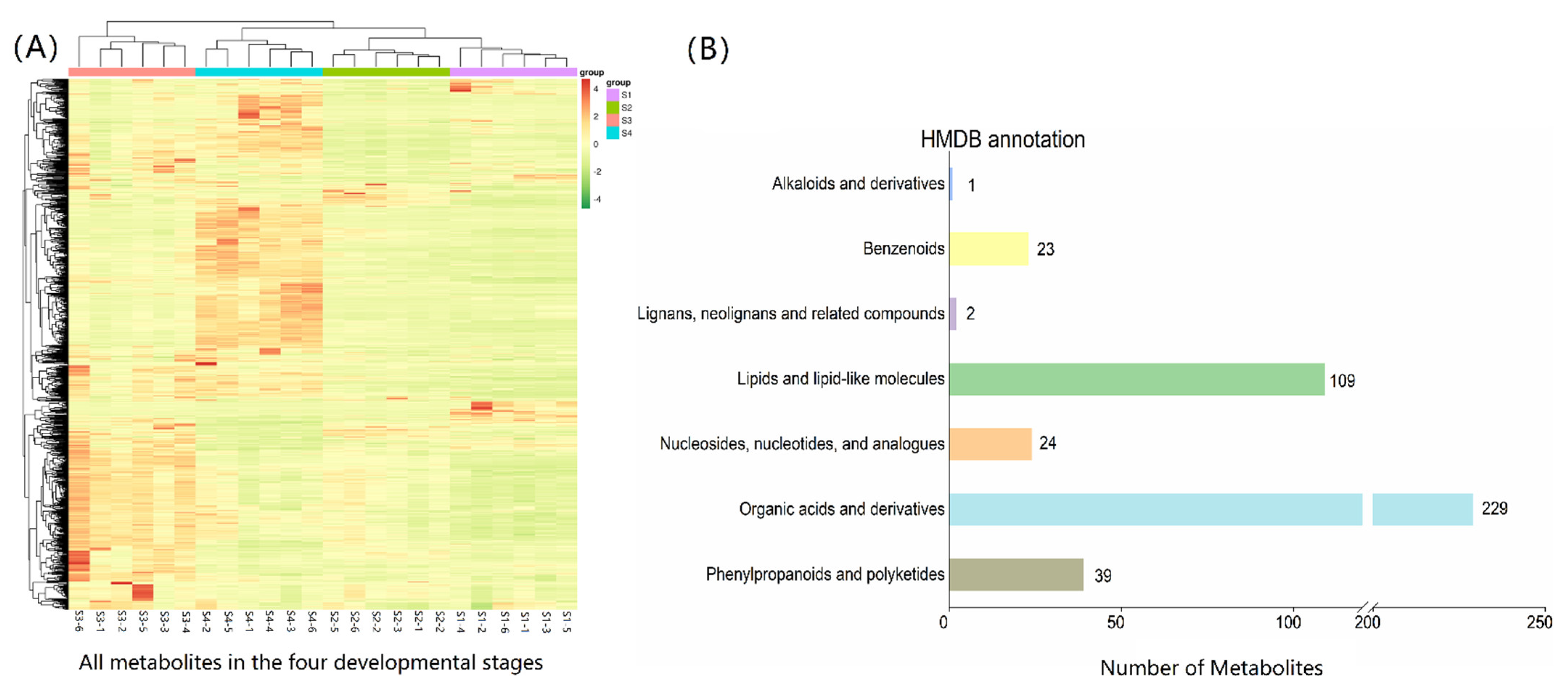
3.4.1. Metabolic Data Evaluation
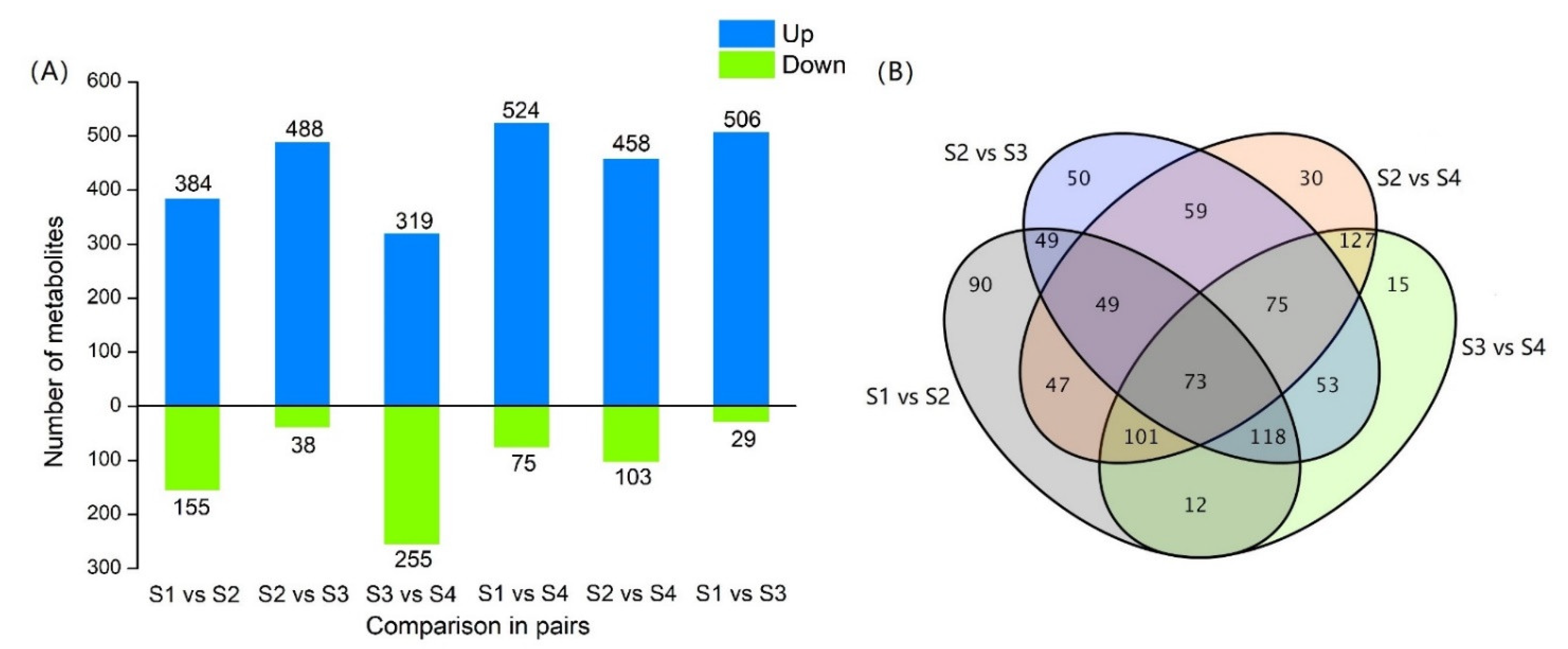
3.4.2. Analysis of Differential Metabolites
3.4.3. Analysis of Differential Metabolites
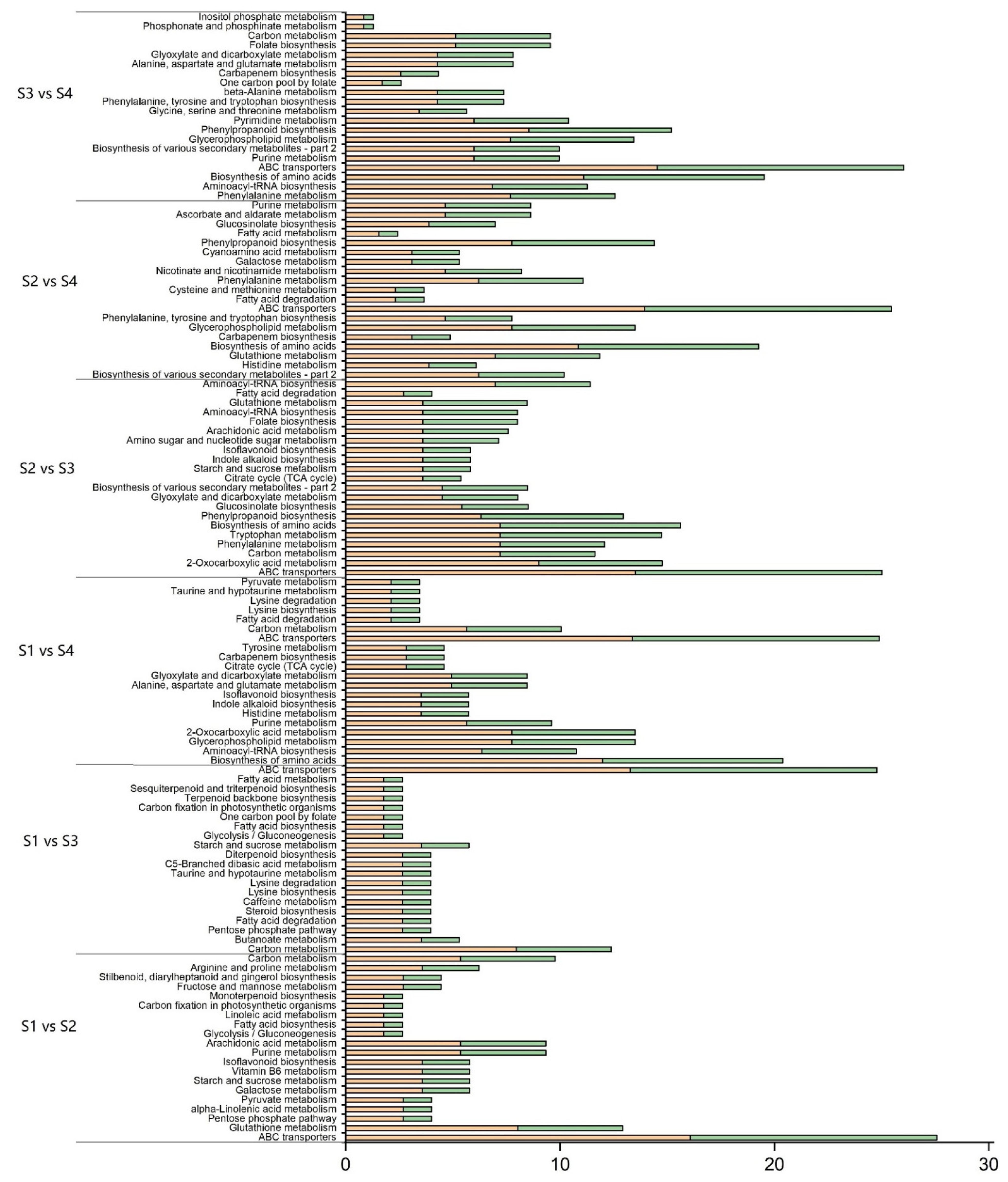
3.4.4. Analysis of Differential Metabolite Pathways
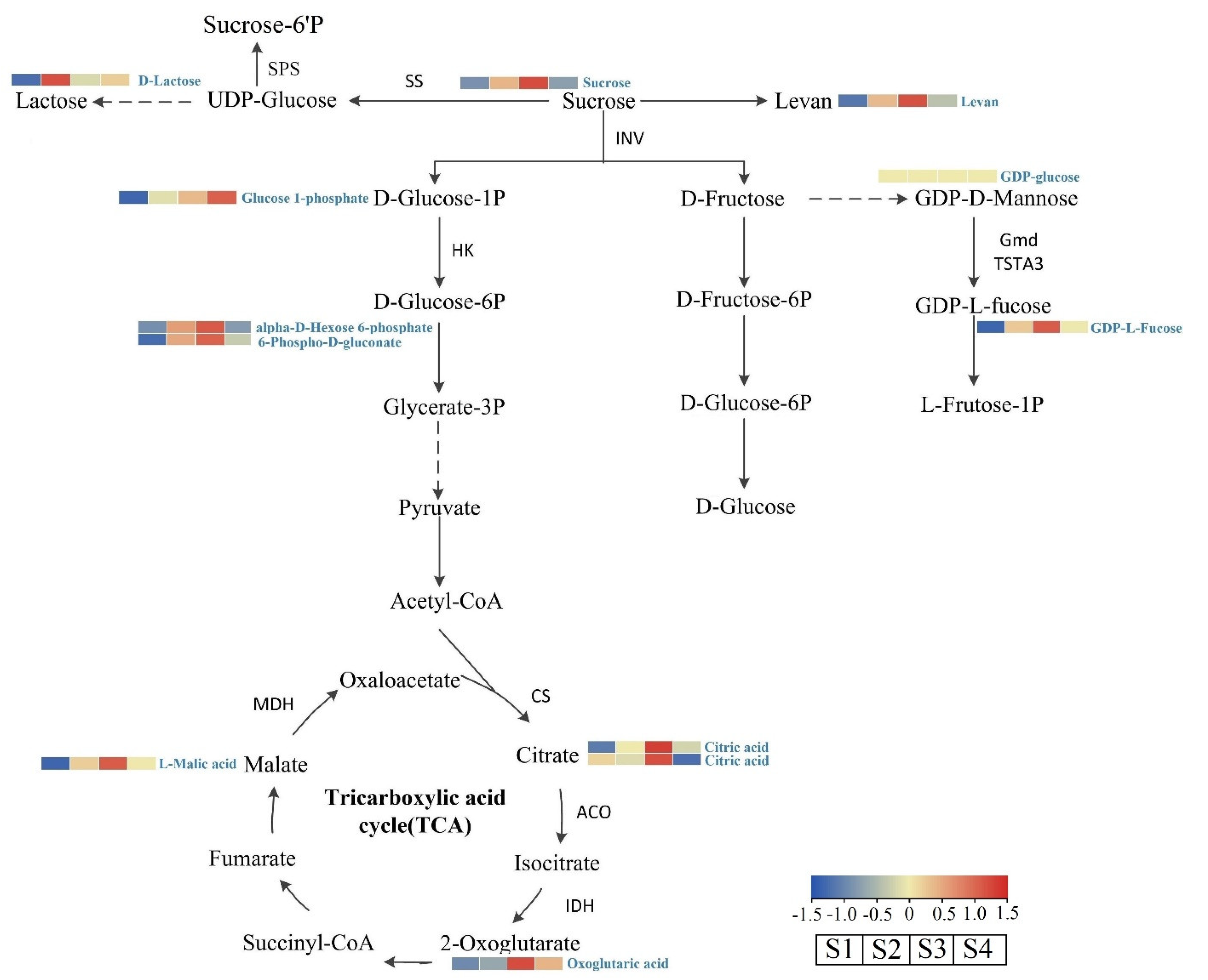
3.4.5. Analysis of Metabolites Related to Sugar and Acid Biosynthetic Pathways
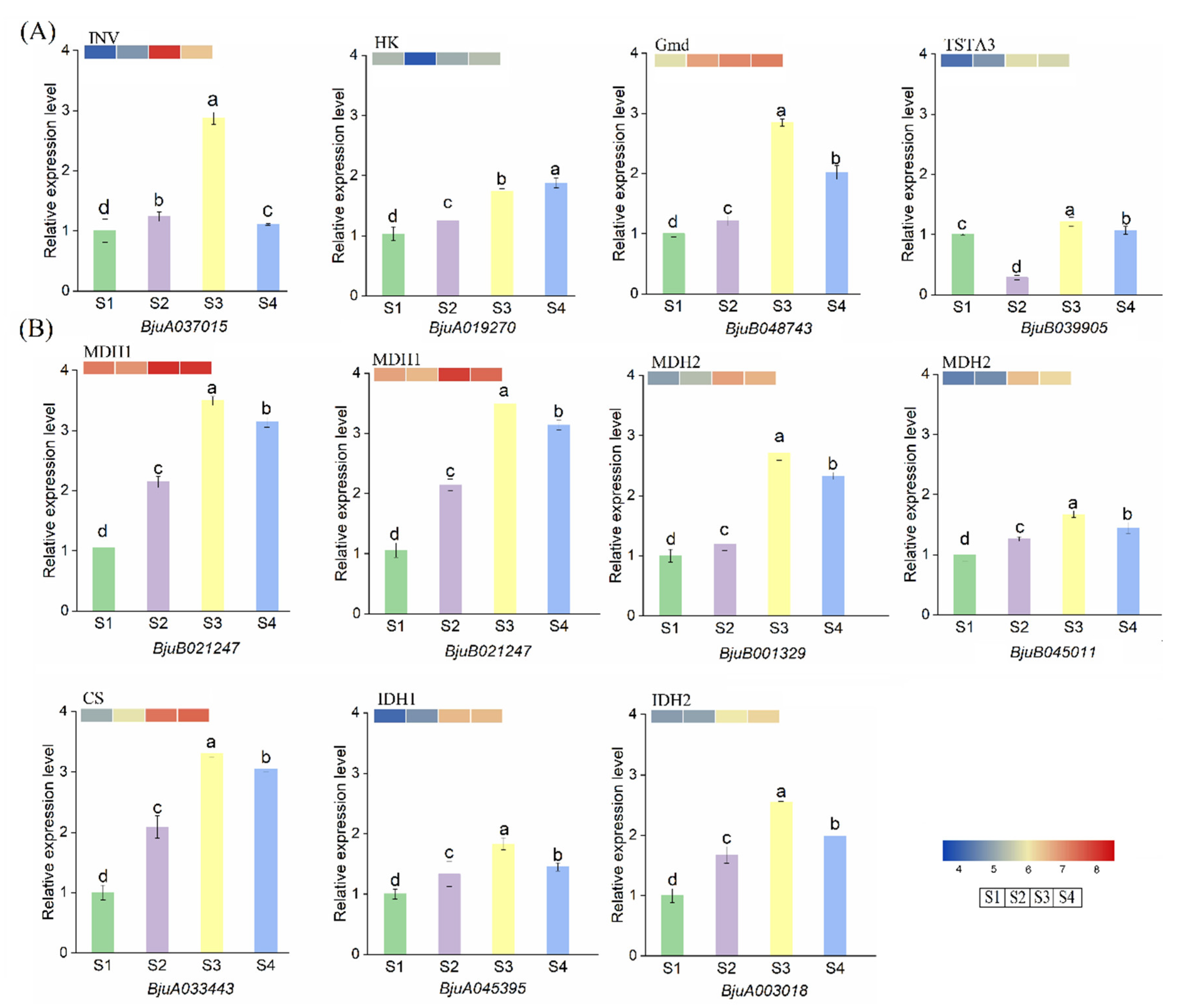
3.5. The Expression Level of Genes Related to Sugar and Acid Biosynthesis Pathway by qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, M.; Sun, B.; Xie, F.; Gong, R.; Luo, Y.; Zhang, F.; Yan, Z.; Tang, H. Identification of the GRAS gene family in the Brassica juncea genome provides insight into its role in stem swelling in stem mustard. PeerJ 2019, 7, e6682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.H.; Zhang, M.F.; Yang, J.H. Molecular phylogeny of Chinese vegetable mustard (Brassica juncea) based on the internal transcribed spacers (ITS) of nuclear ribosomal DNA. Genet. Resour. Crop Evol. 2007, 54, 1709–1716. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, C.; Zhao, N.; Zhang, L.; Hu, Z.; Chen, S.; Zhang, M. Chinese root-type mustard provides phylogenomic insights into the evolution of the multi-use diversified allopolyploid Brassica juncea. Mol. Plant. 2018, 11, 512–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Cao, J.; Hao, N.; Wu, T. Genetic and molecular regulation mechanisms in the formation and development of vegetable fruit shape. Appl. Sci. 2022, 12, 1514. [Google Scholar] [CrossRef]
- Seymour, G.B.; Østergaard, L.; Chapman, N.H.; Knapp, S.; Martin, C. Fruit development and ripening. Annu. Rev. Plant Biol. 2013, 64, 219–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Nie, J.Y.; Li, J.; Zhang, H.; Ye, L.I.; Farooq, S.; Bacha, S.A.S.; Wang, J. Evaluation of sugar and organic acid composition and their levels in highbush blueberries from two regions of China. J. Integr. Agric. 2020, 19, 2352–2361. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Loannidou, S.; Nikoloudakis, N.; Seraphides, N.; Papayiannis, L.C.; Kyratzis, A.C. Physicochemical characterization and trait stability in a genetically diverse ex situ collection of pomegranate (Punica granatum L.) germplasm from Cyprus. Sci. Hortic. 2020, 263, 109116. [Google Scholar] [CrossRef]
- Aslam, M.M.; Deng, L.; Wang, X.B.; Wang, Y.; Pan, L.; Liu, H.; Niu, L.; Lu, Z.H.; Cui, G.C.; Zeng, W.F.; et al. Expression patterns of genes involved in sugar metabolism and accumulation during peach fruit development and ripening. Sci. Hortic. 2019, 257, 108633. [Google Scholar] [CrossRef]
- Jia, H.F.; Jiu, S.T.; Zhang, C.; Wang, C.; Tariq, P.; Liu, Z.J.; Wang, B.J.; Cui, L.W.; Fang, J.G. Abscisic acid and sucrose regulate tomato and strawberry fruit ripening through the abscisic acid-stress-ripening transcription factor. Plant Biotechnol. J. 2016, 14, 2045–2065. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Lin, L.; Yang, L.; Liao, M.; Wang, X.; Wang, J.; Lv, X.; Deng, H.; Liang, D.; Xia, H.; et al. Exogenous melatonin promotes growth and sucrose metabolism of grape seedlings. PLoS ONE 2020, 15, e0232033. [Google Scholar] [CrossRef]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Mirajkar, S.J.; Suprasanna, P.; Vaidya, E.R. Spatial distribution and dynamics of sucrose metabolising enzymes in radiation induced mutants of sugarcane. Plant Physiol. Biochem. 2016, 100, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Shao, X.F.; Wei, Y.Y.; Xu, F.; Wang, H.F. Sucrose degradation is regulated by 1-methycyclopropene treatment and is related to chilling tolerance in two peach cultivars. Postharvest Biol. Technol. 2017, 124, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Xu, J.F.; Duan, S.G.; Bian, C.S.; Li, G.C.; Jin, L.P. Transcriptome analysis of potato tuber expansion stage. Potato Ind. Green Dev. 2021, 261–262. [Google Scholar] [CrossRef]
- Zhang, X.M.; Liu, S.H.; Du, L.Q.; Yao, Y.L.; Wu, J.Y. Activities, transcript levels, and subcellular localizations of sucrose phosphate synthase, sucrose synthase, and neutral invertase and change in sucrose content during fruit development in pineapple (Ananas comosus). J. Hortic. Sci. Biotechnol. 2019, 94, 573–579. [Google Scholar] [CrossRef]
- Liu, H.F.; Génard, M.; Guichard, S.; Bertin, N. Model-assisted analysis of tomato fruit growth in relation to carbon and water fluxes. J. Exp. Bot. 2007, 58, 3567–3580. [Google Scholar] [CrossRef] [Green Version]
- Sweetlove, L.J.; Beard, K.F.; Nunes-Nesi, A.; Fernie, A.R.; Ratcliffe, R.G. Not just a circle: Flux modes in the plant TCA cycle. Trends Plant Sci. 2010, 15, 462–470. [Google Scholar] [CrossRef]
- Li, Q.; Qiao, X.; Jia, L.; Zhang, Y.; Zhang, S. Transcriptome and resequencing analyses provide insight into differences in organic acid accumulation in two pear varieties. Int. J. Mol. Sci. 2021, 22, 9622. [Google Scholar] [CrossRef]
- Liu, X.C.; Lin, X.H.; Liu, S.C.; Zhu, C.Q.; Grierson, D.; Li, S.J.; Chen, K.S. The effect of NH4+ on phosphoenolpyruvate carboxykinase gene expression, metabolic flux and citrate content of citrus juice sacs. Plant Physiol. Biochem. 2021, 167, 123–131. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, C.W.; Bae, D.W.; Do, H.; Jeong, C.S.; Hwang, J.; Cha, S.S.; Lee, J.H. Structural basis of the cooperative activation of type II citrate synthase (HyCS) from Hymenobacter sp. PAMC 26554. Int. J. Biol. Macromol. 2021, 31, 213–221. [Google Scholar] [CrossRef]
- Shukla, R.; Shukla, H.; Tripathi, T. Structure-based discovery of phenyl-diketo acids derivatives as Mycobacterium tuberculosis malate synthase inhibitors. J. Biomol. Struct. Dyn. 2021, 39, 2945–2958. [Google Scholar] [CrossRef] [PubMed]
- Cirilli, M.; Bassi, D.; Ciacciulli, A. Sugars in peach fruit: A breeding perspective. Hortic. Res. 2016, 20, 15067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.B.; Zhao, L.; Jiang, X.H.; Cherono, S.; Liu, J.J.; Ogutu, C.; Ntini, C.; Zhang, X.; Han, Y. Assessment of organic acid accumulation and its related genes in peach. Food Chem. 2021, 334, 127567. [Google Scholar] [CrossRef] [PubMed]
- Castro-Moretti, F.R.; Gentzel, I.N.; Mackey, D.; Alonso, A.P. Metabolomics as an Emerging Tool for the Study of Plant-Pathogen Interactions. Metabolites 2020, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Liu, M.; Qin, N.; Li, S.; Yu, M.; Wang, C.; Ma, Q. Lipidomics coupled with pathway analysis characterizes serum metabolic changes in response to potassium oxonate induced hyperuricemic rats. Lipids Health Dis. 2019, 18, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Liang, X.; Wei, T.; Liu, N.; Wang, Y.; Zou, L.; Bai, C.; Yao, Y.; Wu, T.; Kong, L.; et al. A metabolomics study: CdTe/ZnS quantum dots induce polarization in mice microglia. Chemosphere 2020, 246, 125629. [Google Scholar] [CrossRef]
- Xu, J.D.; Yan, J.J.; Li, W.J.; Wang, Q.Y.; Wang, C.X.; Guo, J.X.; Geng, D.L.; Guan, Q.M.; Ma, F.W. Integrative Analyses of Widely Targeted Metabolic Profiling and Transcriptome Data Reveals Molecular Insight into Metabolomic Variations during Apple (Malus domestica) Fruit Development and Ripening. Int. J. Mol. Sci. 2020, 21, 4797. [Google Scholar] [CrossRef]
- Gong, C.; Diao, W.; Zhu, H.; Umer, M.J.; Zhao, S.; He, N.; Lu, X.; Yuan, P.; Anees, M.; Yang, D.; et al. Metabolome and transcriptome integration reveals insights into flavor formation of ‘Crimson’ watermelon flesh during fruit development. Front. Plant Sci. 2021, 12, 629361. [Google Scholar] [CrossRef]
- Wang, S.S.; Liu, L.; Mi, X.Z.; Zhao, S.Q.; An, Y.L.; Xia, X.B.; Guo, R.; Wei, C.L. Multi-omics analysis to visualize the dynamic roles of defense genes in the response of tea plants to gray blight. Plant J. 2021, 106, 862–875. [Google Scholar] [CrossRef]
- Slade, W.O.; Werth, E.G.; McConnell, E.W.; Alvarez, S.; Hicks, L.M. Quantifying reversible oxidation of protein thiols in photosynthetic organisms. J. Am. Soc. Mass Spectrom. 2015, 26, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, C.; Tautenhahn, R.; Böttcher, C.; Larson, T.R.; Neumann, S. CAMERA: An integrated strategy for compound spectra extraction and annotation of liquid chromatography/mass spectrometry data sets. Anal. Chem. 2012, 84, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.Y.; Xie, F.J.; He, Q.; Li, J.; Liu, J.; Sun, B.; Luo, Y.; Zhang, Y.; Chen, Q.; Zhang, F.; et al. Expression analysis of XTH in stem swelling of stem mustard and selection of reference genes. Genes 2020, 20, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucl. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.S.; Xie, C.H.; Yu, X.C.; Hu, X.Y.; Zhu, S.H.; Li, J. Effect of Sowing Date and Date of Harvest on Yield and Shape of Tumorous Stem in Brassica juncea var. tumida Tsen et Lee. North. Hortic. 2012, 22, 33–35. [Google Scholar]
- Yang, S.W.; Rao, L.; Hu, H.D.; Zhao, S.Z.; Lin, H.Q. Comparison of differences among different tumorous stem mustard cultivars under direct seeding cultivation. J. Chang. Veg. 2022, 8, 53–55. [Google Scholar] [CrossRef]
- Zhang, D.; Tian, J.Q.; Cheng, Y.J.; Ma, X.H.; Huang, K.; Deng, J.; Li, C.S.; Liu, X. Desalting process optimization and quality analysis of tuber mustard. Food Ferment. Ind. 2017, 43, 167–172. [Google Scholar] [CrossRef]
- Kortstee, A.J.; Appeldoorn, N.J.; Oortwijn, M.E.; Visser, R.G. Differences in regulation of carbohydrate metabolism during early fruit development between domesticated tomato and two wild relatives. Planta 2007, 226, 929–939. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.W.; Li, M.; Zhang, C.M.; Tan, Q.L.; Yang, X.Z.; Sun, X.C.; Pan, Z.Y.; Deng, X.X.; Hu, C.X. Effects of potassium on fruit soluble sugar and citrate accumulations in Cara Cara navel orange (Citrus sinensis L. Osbeck). Sci. Hortic. 2021, 283, 110057. [Google Scholar] [CrossRef]
- Li, M.; Feng, F.; Cheng, L. Expression patterns of genes involved in sugar metabolism and accumulation during apple fruit development. PLoS ONE 2012, 7, e33055. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.Z.; Lin, X.K.; Du, J.J.; Peng, J.J.; Zhou, K.B. Transcriptomic analysis reveals key genes regulating organic acid synthesis and accumulation in the pulp of Litchi chinensis Sonn. cv. Feizixiao. Sci. Hortic. 2022, 303, 111220. [Google Scholar] [CrossRef]
- Zhang, Q.; Dong, X.Q.; Lin, X.; Peng, J.J.; Luo, D.C.; Huang, S.A.; Zhu, S.L. Analysis of primary metabolites in three plum varieties using ultra-high performance liquid chromatography tandem mass spectrometry. Food Sci. 2022, 43, 1–13. [Google Scholar] [CrossRef]
- Yang, J.J.; Zhu, L.C.; Cui, W.F.; Zhang, C.; Li, D.X.; Ma, B.Q.; Cheng, L.L.; Ruan, Y.L.; Ma, F.W.; Li, M.J. Increased activity of MdFRK2, a high-affinity fructokinase, leads to upregulation of sorbitol metabolism and downregulation of sucrose metabolism in apple leaves. Hortic. Res. 2018, 5, 54–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.C.; Li, B.Y.; Wu, L.M.; Li, H.X.; Wang, Z.Y.; Wei, X.Y.; Ma, B.Q.; Zhang, Y.F.; Ma, F.W.; Ruan, Y.L.; et al. MdERDL6-mediated glucose efflux to the cytosol promotes sugar accumulation in the vacuole through upregulating TSTs in apple and tomato. Proc. Natl. Acad. Sci. USA 2021, 118, e2022788118. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Shen, D.; Yang, W.; Zhang, M.; Zeng, Y.; Xu, J.; Deng, X.; Cheng, Y. Gaba pathway rate-limit citrate degradation in postharvest citrus fruit evidence from hb pumelo (citrus grandis) × fairchild (citrus reticulata) hybrid population. J. Agric. Food Chem. 2017, 65, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Xu, C.; Chen, J.L.; Cao, B.S. Changes of carbohydrates during lotus rhizome development of nelumbo nucifera gaertn. J. Yangzhou Univ. (Agric. Life Sci. Ed.) 2003, 24, 72–74+78. [Google Scholar] [CrossRef]
- Radchuk, V.; Riewe, D.; Peukert, M.; Matros, A.; Strickert, M.; Radchuk, R.; Weier, D.; Steinbiß, H.H.; Sreenivasulu, N.; Weschke, W.; et al. Down-regulation of the sucrose transporters HvSUT1 and HvSUT2 affects sucrose homeostasis along its delivery path in barley grains. J. Exp. Bot. 2017, 68, 4595–4612. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Xie, F.J.; Li, J.; Sun, B.; Luo, Y.; Zhang, Y.; Chen, Q.; Wang, Y.; Zhang, F.; Zhang, Y.T.; et al. Tumorous stem development of Brassica Juncea: A complex regulatory network of stem formation and identification of key genes in glucosinolate biosynthesis. Plants 2020, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Umer, M.J.; Bin Safdar, L.; Gebremeskel, H.; Zhao, S.J.; Yuan, P.L.; Zhu, H.J.; Kaseb, M.O.; Anees, M.; Lu, X.; He, N.; et al. Identification of key gene networks controlling organic acid and sugar metabolism during watermelon fruit development by integrating metabolic phenotypes and gene expression profiles. Hortic. Res. 2020, 7, 193. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, F.; Xuan, Z.; Chen, W.; Du, J.; Wang, Y.; Huo, E.; Guo, L.; Li, M. Metabolomics Analysis Reveals Dynamic Accumulation of Sugar and Acid during Stem Development of Brassica juncea. Agronomy 2022, 12, 3227. https://doi.org/10.3390/agronomy12123227
Xie F, Xuan Z, Chen W, Du J, Wang Y, Huo E, Guo L, Li M. Metabolomics Analysis Reveals Dynamic Accumulation of Sugar and Acid during Stem Development of Brassica juncea. Agronomy. 2022; 12(12):3227. https://doi.org/10.3390/agronomy12123227
Chicago/Turabian StyleXie, Fangjie, Zhengying Xuan, Wanqin Chen, Jiageng Du, Yalin Wang, Entong Huo, Ling Guo, and Mengyao Li. 2022. "Metabolomics Analysis Reveals Dynamic Accumulation of Sugar and Acid during Stem Development of Brassica juncea" Agronomy 12, no. 12: 3227. https://doi.org/10.3390/agronomy12123227