
Beneficial Effect of Exogenously Applied Calcium Chloride on the Anatomy and Fast Chlorophyll Fluorescence in Rhododendron × pulchrum Leaves Following Short-Term Heat Stress Treatment

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Microstructure of the Leaves and Ultrastructure of Leaf Epidermal Cells
2.3. Measurement of PF, DF and MR Kinetics
2.4. Statistical Analysis
3. Results
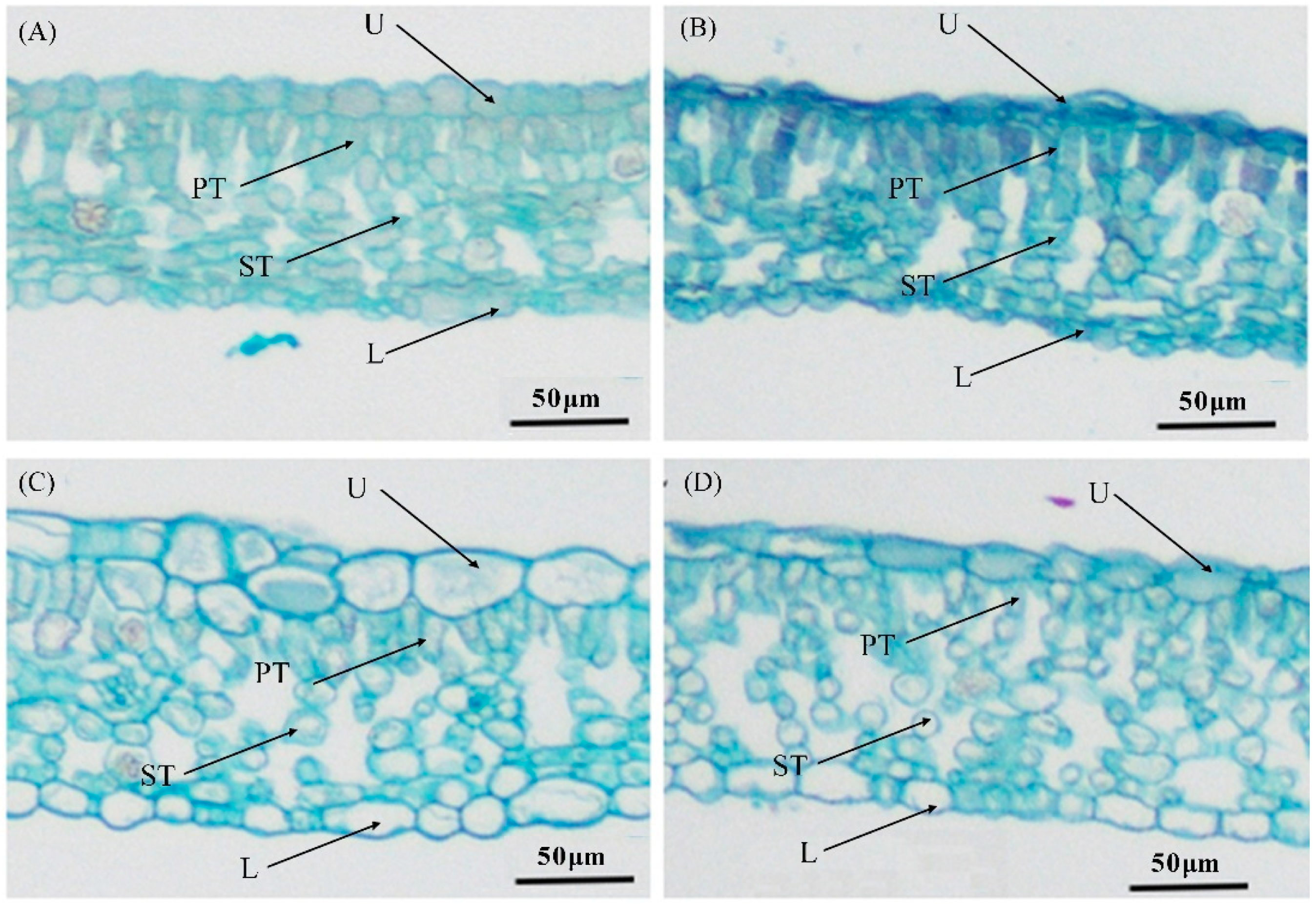
3.1. Phenotype and Microstructure of the Leaf
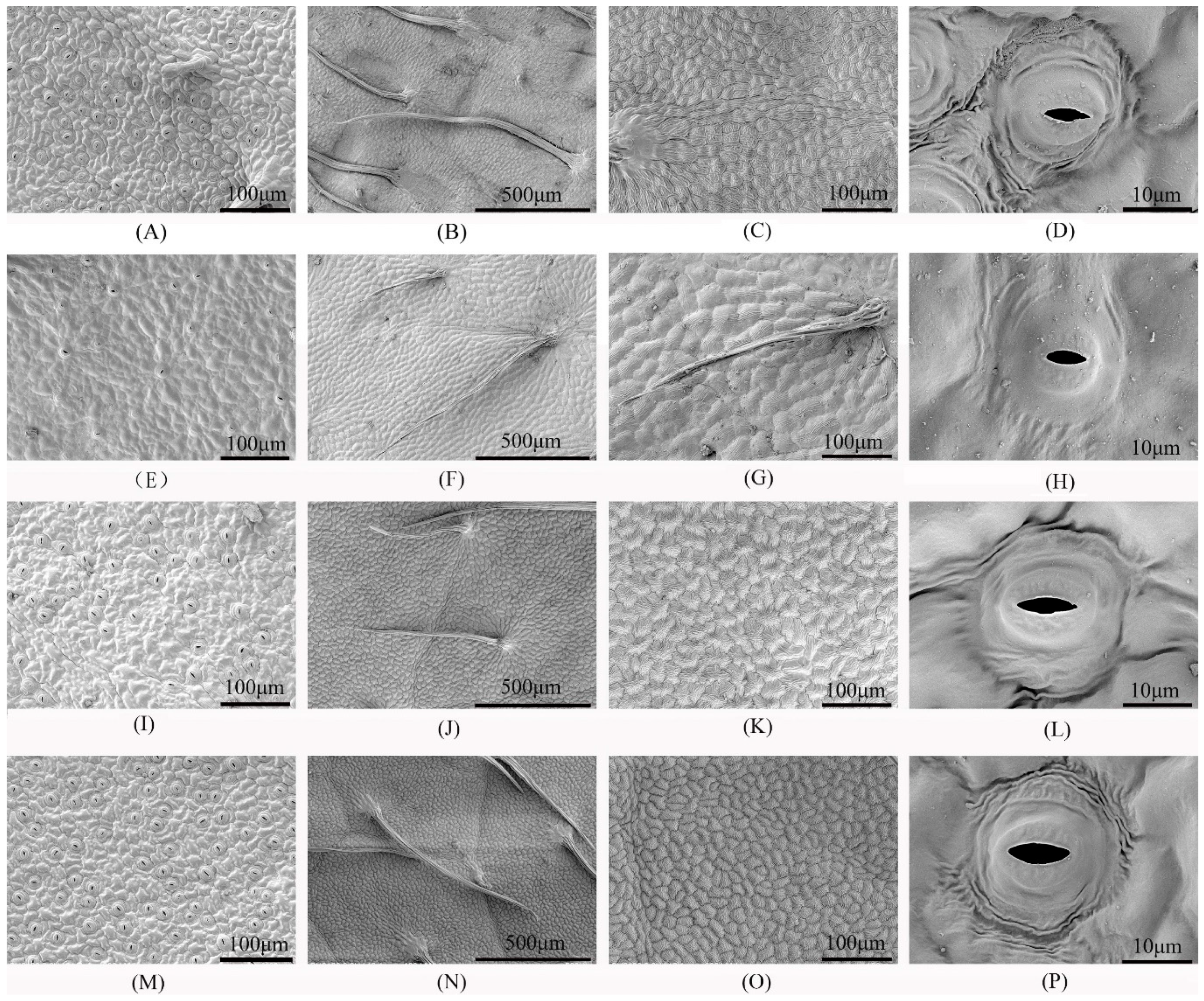
3.2. Ultrastructure of Leaf Epidermal Cells and Stomata
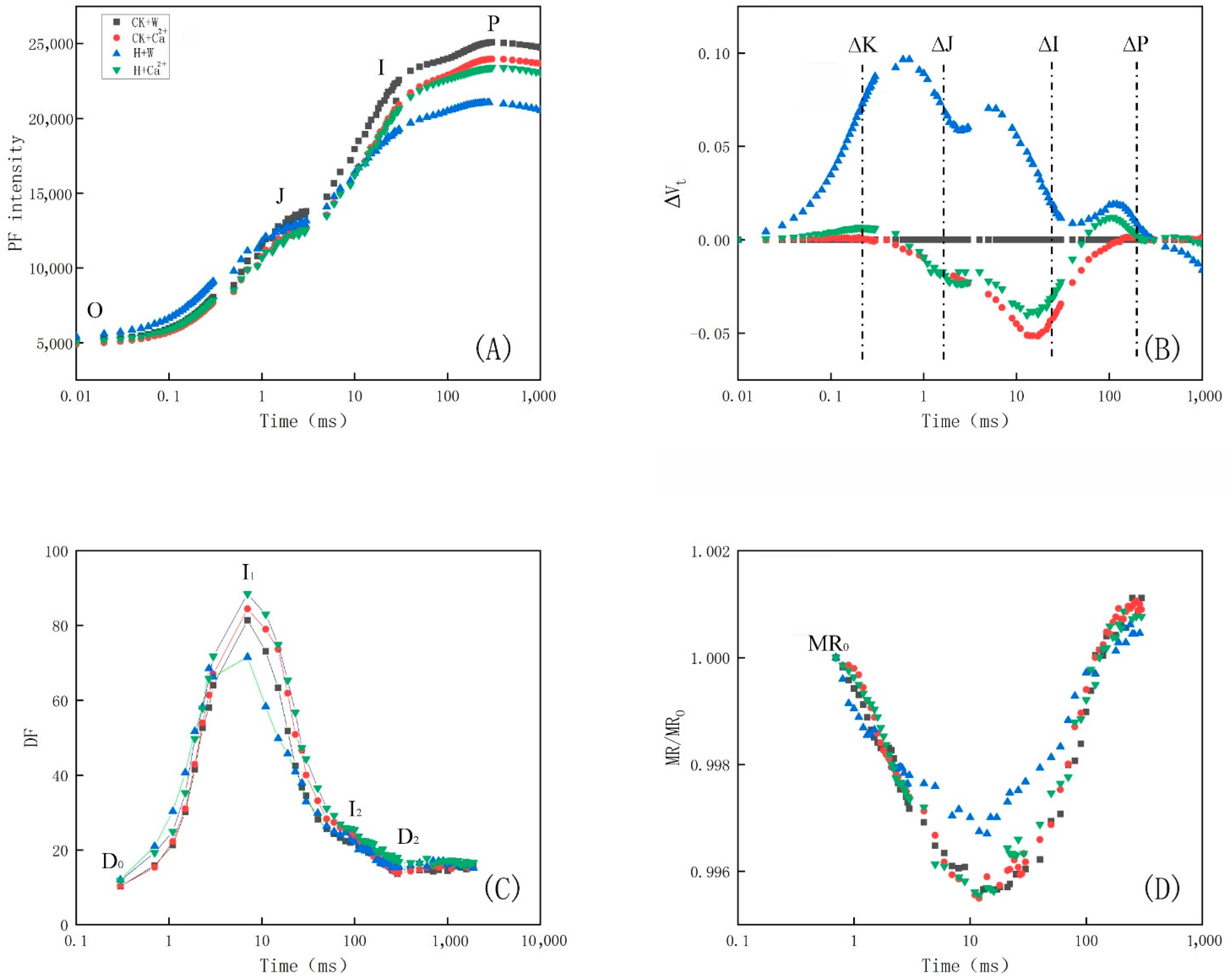
3.3. Analysis of PF, DF and MR Kinetics
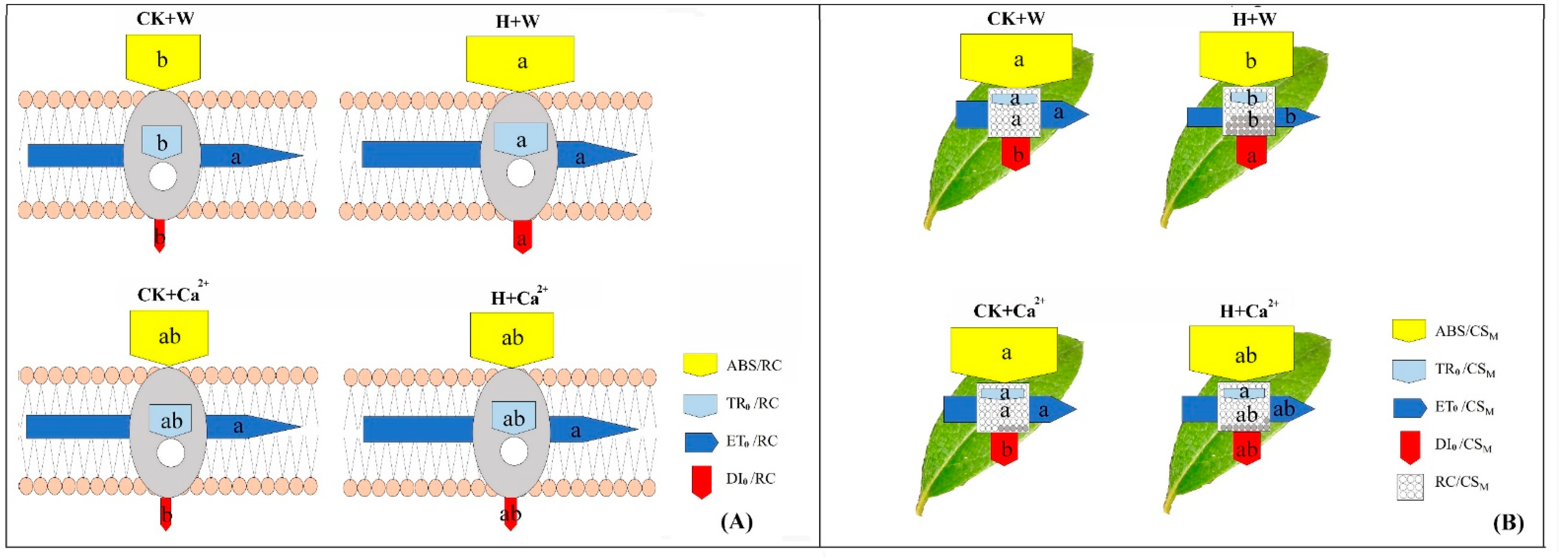
4. Discussion
4.1. Exogenous CaCl2 Influences Leaf Structure under Heat Stress
4.2. Exogenous CaCl2 Influences Leaf Chlorophyll Fluorescence in R. × pulchrum under Heat Stress
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Lavania, D.; Dhingra, A.; Siddiqui, M.H.; Al-Whaibi, M.H.; Grover, A. Current status of the production of high temperature tolerant transgenic crops for cultivation in warmer climates. Plant Physiol. Biochem. 2015, 86, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Grover, A.; Mittal, D.; Negi, M.; Lavania, D. Generating high temperature tolerant transgenic plants: Achievements and challenges. Plant Sci. 2013, 205, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Laanemets, K.; Brandt, B.; Li, J.L.; Merilo, E.; Wang, Y.F.; Keshwani, M.M.; Taylor, S.S.; Kollist, H.; Schroeder, J.I. Calcium dependent and independent stomatal signaling network and compensatory feedback control of stomatal opening via Ca2+ sensitivity priming. Plant Physiol. 2013, 163, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janda, M.; Ruellan, E. Magical mystery tour: Salicylic acid signalling. Environ. Expt. Bot. 2015, 114, 117–128. [Google Scholar] [CrossRef]
- Gao, Y. Molecular and genetic evidence for the key role of atcam3 in heat-shock signal transduction in Arabidopsis. Plant Physiol. 2009, 149, 1773. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Xu, X.; Li, Y.; Wang, H.; Li, Z. Comparative transcriptome analysis reveals differential transcription in heat-susceptible and heat-tolerant pepper (Capsicum annum L.) cultivars under heat stress. J. Plant Biol. 2015, 58, 411–424. [Google Scholar] [CrossRef]
- Anireddy, S.N.R.; Gul, S.A.; Helena, C.; Irene, S.D. Coping with Stresses: Roles of calcium- and calcium/calmodulin-regulated gene expression. The Plant Cell 2011, 23, 2010–2032. [Google Scholar] [CrossRef] [Green Version]
- Snedden, W.A.; Fromm, H. Calmodulin as a versatile calcium signal transducer in plants. New Phytol. 2001, 151, 35–66. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Bing, Z.; Xu, J.; Zheng, X.; Huang, W. Effects of salicylic acid and calcium chloride on heat tolerance in Rhododendron ’fen zhen zhu’. J. Am. Soc. Hortic. Sci. 2016, 141, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Mishkind, M.; Vermeer, J.E.M.; Darwish, E.; Munnik, T. Heat stress activates phospholipase D and triggers PIP accumulation at the plasma membrane and nucleus. The Plant J. 2009, 60, 10–21. [Google Scholar] [CrossRef]
- Perez, D.E.; Hoyer, J.S.; Johnson, A.I.; Moody, Z.R.; Lopez, J.; Kaplinsky, N.J. BOBBER1 is a noncanonical Arabidopsis small heat shock protein required for both development and thermotolerance. Plant Physiol. 2009, 151, 241–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.L.; Yang, W.J.; Lo, H.F.; Yeh, D.M. Physiology, anatomy, and cell membrane thermostability selection of leafy radish (Raphanus sativus var. Oleiformis Pers.) with different tolerance under heat stress. Sci. Hortic. 2014, 179, 367–375. [Google Scholar] [CrossRef]
- Wang, J.; Lv, J.H.; Liu, Z.B.; Liu, Y.H.; Song, J.S.; Ma, Y.Q. Integration of transcriptomics and metabolomics for Pepper (Capsicum annuum L.) in response to heat stress. Int. J. Mol. Sci. 2020, 20, 5042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, Q.; Xi, X.; He, Y.; Jiang, A. Transcriptomic analysis of the leaves of two grapevine cultivars under high-temperature stress. Sci. Hortic. 2020, 265, 109265. [Google Scholar] [CrossRef]
- Salem-Fnayou, A.B.; Bouamama, B.; Ghorbel, A.; Mliki, A. Investigations on the leaf anatomy and ultrastructure of grapevine (Vitis vinifera) under heat stress. Microsc. Res. Tech. 2011, 74, 756–762. [Google Scholar] [CrossRef]
- Shen, H.F.; Zhao, B.; Xu, J.J.; Liang, W.; Huang, W.M.; Li, H.H. Effects of heat stress on changes in physiology and anatomy in two cultivars of Rhododendron. S. Afr. J. Bot. 2017, 112, 338–345. [Google Scholar] [CrossRef]
- Feng, J.Q.; Wang, J.H.; Zhang, S.B. Leaf physiological and anatomical responses of two sympatric paphiopedilum species to temperature. Plant Divers. 2022, 44, 101e108. [Google Scholar] [CrossRef]
- Mathur, S.; Jajoo, A. Photosynthesis: Limitations in response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Oba, E.G.; Hu, J.; Zhang, Z.; Li, S.; Xie, W. Physiological response of three micropropagated plant lets of Rhododendron hybrids for heat stress. Eur. J. Hortic. Sci. 2021, 86, 300–307. [Google Scholar] [CrossRef]
- Ishikawa, T.; Shigeoka, S. Recent advances in ascorbate biosynthesis and the physiological significance of ascorbate peroxidase in photosynthesizing organisms. Biosci. Biotechnol. Biochem. 2008, 72, 1143–1154. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Y.; Yu, W.; Hänninen, H.; Song, L.; Du, X.; Zhang, R.; Wu, J. Novel insights into the influence of seed sarcotesta photosynthesis on accumulation of seed dry matter and oil content in Torreya grandis cv. ‘Merrillii’. Front. Plant Sci. 2018, 8, 2179. [Google Scholar] [CrossRef] [Green Version]
- Oukarroum, A.; Goltsev, V.; Strasser, R.J. Temperature effects on pea plants probed by simultaneous measurements of the kinetics of prompt fluorescence, delayed fluorescence and modulated 820 nm reflection. PLoS ONE 2013, 8, e59433. [Google Scholar] [CrossRef] [PubMed]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta 2007, 1767, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zushi, K.; Kajiwara, S.; Matsuzoe, N. Chlorophyll a fluorescence OJIP transient as a tool to characterize and evaluate response to heat and chilling stress in tomato leaf and fruit. Sci. Hortic. 2012, 148, 39–46. [Google Scholar] [CrossRef]
- Sharma, D.K.; Fernández, J.O.; Rosenqvist, E.; Ottosen, C.O.; Andersen, S.B. Genotypic response of detached leaves versus intact plants for chlorophyll fluorescence parameters under high temperature stress in wheat. Plant Physiol. 2014, 171, 576–586. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.D.; Maria, T.A.; Eva, R.; Carl-Otto, O.; Andersen, S.B. QTLs and potential candidate genes for heat stress tolerance identified from the mapping populations specifically segregating for Fv/Fm in wheat. Front. Plant Sci. 2017, 8, 1668. [Google Scholar] [CrossRef] [Green Version]
- Jedmowski, C.; Brüggemann, W. Imaging of fast chlorophyll fluorescence induction curve (OJIP) parameters, applied in a screening study with wild barley (Hordeum spontaneum) genotypes under heat stress. J. Photochem. Photobiol. B Biol. 2015, 151, 153–160. [Google Scholar] [CrossRef]
- Sohn, S.I.; Yong, H.L.; Sun, H.H.; Chang, S.K.; Oh, Y.J. Characterization and evaluation of response to heat and chilling stress in exotic weeds using chlorophyll a fluorescence OJIP transient. Environ. Biol. Res. 2020, 38, 450–460. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, M.; Xu, J.; Zhou, W.; Cao, L. Transcriptomic analysis of short-term heat stress response in Pinellia ternata provided novel insights into the improved thermotolerance by spermidine and melatonin. Ecotoxicol. Environ. Safe 2020, 202, 110877. [Google Scholar] [CrossRef]
- Essemine, J.; Qu, M.; Lyu, M.; Song, Q.; Zhu, X.G. Photosynthetic and transcriptomic responses of two C4 grass species with different nacl tolerance. J. Plant Physiol. 2020, 253, 153244. [Google Scholar] [CrossRef]
- Wang, X.; Dingxuan, Q.; Shi, M. Calcium amendment for improved germination, plant growth, and leaf photosynthetic electron transport in oat (Avena sativa) under NaCl stress. PLoS ONE 2021, 16, e0256529. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Kluwer Academic Publishers Press: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Wang, J.W.; Liu, Y.; Xu, Y.X.; Chen, W.J.; Han, Y.N.; Wang, G.G.; Jin, S. Sexual diferences in gas exchange and chlorophyll fuorescence of Torreya grandis under drought stress. Trees 2022, 36, 283–294. [Google Scholar] [CrossRef]
- Ballantyne, D.J.; Argent, G.; Mcfarlane, M. Dormancy, flowering and chlorophyll fluorescence of a rhododendron and azaleas in the greenhouse. In Rhododendrons in Horticulture & Science Papers Presented at the International Rhododendron Conference, Proceedings of the International Rhododendron Conference, Edinburgh, UK, 17–19 May 2002; Royal Botanic Garden Edinburgh: Edinburgh, UK, 2003. [Google Scholar]
- Fang, L.; Tong, J.; Dong, Y.; Xu, D.; Jing, M.; Yuan, Z. De novo RNA sequencing transcriptome of Rhododendron obtusum identified the early heat response genes involved in the transcriptional regulation of photosynthesis. PLoS ONE 2017, 12, e0186376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, J. Hardy Rhododendron Species: A Guide to Identification; Timber Press Inc.: Portland, OR, USA, 2005. [Google Scholar]
- Krebs, S.L. Rhododendron. In Ornamental Crops, Handbook of Plant Breeding; Van Huylenbroeck, J., Ed.; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Xia, X.M.; Yang, M.Q.; Li, C.L.; Huang, S.X.; Jin, W.T.; Shen, T.T. Spatiotemporal evolution of the global species diversity of rhododendron. Mol. Biol. Evol. 2021, 39, msab314. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Nielsen, J.; Chamberlain, D.F.; Li, X.; Sun, W. The conservation of Rhododendrons is of greater urgency than has been previously acknowledged in China. Biodivers. Conserv. 2014, 23, 3149–3154. [Google Scholar] [CrossRef]
- Rennenberg, H.; Loreto, F.; Polle, A.; Brilli, F.; Fares, S.; Beniwal, R.S.; Gessler, A.J. Physiological responses of forest trees to heat and drought. Plant Biol. 2006, 8, 556–571. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, H.; Peng, S.M.; Korpelainen, H.; Li, C.Y. Sex-related differences in morphological, physiological, and ultrastructural responses of Populus cathayana to chilling. J. Exp. Bot. 2011, 62, 675–686. [Google Scholar] [CrossRef] [Green Version]
- Ruppel, N.J.; Logsdon, C.A.; Whippo, C.W. A mutation in Arabidopsis seedling plastid development affects plastid differentiation in embryo-derived tissues during seedling growth. Plant Physiol. 2011, 155, 342–353. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.X.; Li, Y.Q.; Wang, M.L.; Bai, M.; Zou, R.; Tang, H.; Wu, H. Effects of light intensity on leaf photosynthetic characteristics, chloroplast structure, and alkaloid content of Mahonia bodinieri (Gagnep.) Laferr. Acta Physiol. Plant. 2016, 38, 120. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, L.; Yang, R.; Han, Y.Y.; Hao, J.H.; Liu, C.J.; Fan, S. Effects of exogenous putrescine on the ultrastructure of and calcium ion flow rate in lettuce leaf epidermal cells under drought stress. Hortic. Environ. Biotechnol. 2019, 60, 479–490. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820 nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Kouzmanova, M.; Kalaji, H.M.; Yordanov, I.; Krasteva, V.; Alexandrov, V.; Stefanov, D.; Allakhverdiev, S.I.; et al. Drought induced modifications of photosynthetic electron transport in intact leaves: Analysis and use of neural networks as a tool for a rapid non-invasive estimation. Biochim. Biophys. Acta 2012, 1817, 1490–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Li, X.; Zhu, X.; Ding, Z.; Huang, X.; Chen, X.; Jin, S. Molecular and photosynthetic performance in the yellow leaf mutant of Torreya grandis according to transcriptome sequencing, chlorophyll a fluorescence, and modulated 820 nm reflection. Cells 2022, 11, 431. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.J.; Zhang, Q.X.; Pan, H.T.; Dong, L.L. Preliminary study on physiological indexes of heat-resistance of Primula. Chin. Agric. Sci. Bull. 2010, 26, 158–163. [Google Scholar]
- Bian, A.N.; Lu, Y.L.; Pan, D.M. Preliminary study on heat resistant physiological indexes of daffodils. J. Minnan Norm. Univ. (Nat. Sci.) 2016, 29, 70–75. [Google Scholar] [CrossRef]
- Banon, S.; Fernandez, J.A.; Franco, J.A.; Torrecillas, A.; Alarcón, J.J.; Sánchez-Blanco, M.J. Effects of water stress and night temperature preconditioning on water relations and morphological and anatomical changes of Lotus creticusplants. Sci. Hortic. 2004, 101, 333–342. [Google Scholar] [CrossRef]
- El-Fiki, M.A.; El-Taher, A.M.; EL-Gendy, A.G.; Lila, M.I. Morphological and anatomical studies on some taxa of family Apocynaceae. Al-Azhar J. Agric. Res. 2019, 44, 136–147. [Google Scholar]
- Faralli, M.; Bontempo, L.; Bianchedi, P.L.; Moser, C.; Bertamini, M.; Lawson, T.; Camin, F.; Stefanini, M.; Varotto, C. Natural variation in stomatal dynamics drives divergence in heat stress tolerance and contributes to seasonal intrinsic water-use efficiency in Vitis vinifera (subsp. sativa and sylvestris). J. Exp. Bot. 2022, 73, 3238–3250. [Google Scholar] [CrossRef]
- El-Taher, A.M.; Abd El-Raouf, H.S.; Osman, N.A.; Azoz, S.N.; Omar, M.A.; Elkelish, A.; Abd El-Hady, M.A. Effect of salt stress and foliar application of salicylic acid on morphological, biochemical, anatomical, and productivity characteristics of Cowpea (Vigna unguiculata L.) Plants. Plants 2021, 11, 115. [Google Scholar] [CrossRef]
- Strauss, A.J.; Krüger, G.H.J.; Strasser, R.J.; Heerden, P.D.R.V. Ranking of dark chilling tolerance in soybean genotypes probed by the chlorophyll a fluorescence transient O-J-I-P. Environ. Exp. Bot. 2006, 56, 147–157. [Google Scholar] [CrossRef]
- Oster, U.; Tanaka, R.; Tanaka, A.; Rüdigger, W. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. The Plant J. 2000, 21, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Momchil, P.; Lyubka, K.; Andon, V.; Jaco, V.; Vasilij, G. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Brestic, M.; Tan, D.X.; Zivcak, M.; Zhu, X.; Liu, S.; Song, F.; Reiter, R.J.; Liu, F. Melatonin alleviates low PS I-limited carbon assimilation under elevated CO2 and enhances the cold tolerance of offspring in chlorophyll b-deficient mutant wheat. J. Pineal Res. 2018, 64, e12453. [Google Scholar] [CrossRef]
- Tongra, T.; Mehta, P.; Mathur, S.; Agrawal, D.; Bharti, S.; Los, D.A.; Allakhverdiev, S.I.; Jajoo, A. Computational analysis of fluorescence induction curves in intact spinach leaves treated at different pH. BioSystems 2011, 103, 158–163. [Google Scholar] [CrossRef]
- Mathur, S.; Allakhverdiev, S.I.; Jajoo, A. Analysis of high temperature stress on the dynamics of antenna size and reducing side heterogeneity of photosystem ii in wheat leaves (Triticum aestivum). Biochim. Biophys. Acta 2011, 1807, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Ahammed, G.J.; Xu, W.; Liu, A.; Chen, S. Comt1 silencing aggravates heat stress-induced reduction in photosynthesis by decreasing chlorophyll content, photosystem ii activity and electron transport efficiency in tomato. Front. Plant Sci. 2018, 9, 998. [Google Scholar] [CrossRef]
- Gao, J.; Li, P.; Ma, F.; Goltsev, V. Photosynthetic performance during leaf expansion in malus micromalus probed by chlorophyll a fluorescence and modulated 820 nm reflection. J. Photochem. Photobiol. B Biol. 2014, 137, 144–150. [Google Scholar] [CrossRef]
- Dąbrowski, P.; Kalaji, M.H.; Baczewska, A.H.; Pawluśkiewicz, B.; Mastalerczuk, G.; Borawska-Jarmułowicz, B.; Paunov, M.; Goltsev, V. Delayed chlorophyll a fluorescence, mr 820, and gas exchange changes in perennial ryegrass under salt stress. J. Lumin. 2017, 183, 322–333. [Google Scholar] [CrossRef]
- Adams, W.W.; Demming-Adams, B. Chlorophyll Fluorescence as a Tool to Monitor Plant Response to the Environment; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar] [CrossRef]
- Song, Y.; Chen, Q.; Ci, D.; Shao, X.; Zhang, D. Effects of high temperature on photosynthesis and related gene expression in poplar. BMC Plant Biol. 2014, 14, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitko, K.; Rusinowski, S.; Pogrzeba, M.; Daszkowska-Golec, A.; Makowski, E. Development and aging of photosynthetic apparatus of Vitis vinifera L. during growing season. Photosynthetica 2020, 58, 186–193. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Upper Epidermis Thickness (μm) | Palisade Thickness (μm) | Spongy Thickness (μm) | Lower Epidermis (μm) | Leaf Thickness (μm) | Ratio of Palisade Thickness to Spongy Thickness | Ratio of Palisade Thickness to Leaf Thickness |
|---|---|---|---|---|---|---|---|
| CK + W | 14.96 ± 1.79 b 1 | 33.47 ± 2.51 a | 39.58 ± 1.76 c | 12.27 ± 1.39 b | 100.28 ± 3.23 b | 0.85 ± 0.09 a | 0.33 ± 0.02 a |
| H + W | 11.82 ± 1.70 c | 32.53 ± 3.53 a | 43.69 ± 6.36 c | 10.78 ± 2.25 b | 98.82 ± 9.84 b | 0.75 ± 0.08 b | 0.33 ± 0.02 a |
| CK + Ca2+ | 22.62 ± 3.43 a | 31.81 ± 4.25 a | 53.78 ± 3.32 b | 17.29 ± 3.31 a | 125.53 ± 5.36 a | 0.59 ± 0.10 c | 0.25 ± 0.03 b |
| H + Ca2+ | 15.40 ± 1.53 b | 22.67 ± 6.17 b | 65.91 ± 3.13 a | 13.25 ± 2.90 b | 117.23 ± 6.80 a | 0.34 ± 0.09 d | 0.19 ± 0.04 c |
| Group | Cell Density (No. of Upper Epidermises/mm2) | Stomatal Density (No. of Stomata/mm2) | Stomatal Transverse Diameter/μm | Stomatal Longitudinal Diameter/μm | Stomatal Aperture/μm2 | Open Stomatal Density (No. of Opened Stomata/mm2) | Closed Stomatal Density (No. of Closed Stomata/mm2) | Ratio of Closed/Open Stomata |
|---|---|---|---|---|---|---|---|---|
| CK + W | 217 ± 5 b 1 | 71 ± 3 a | 5.56 ± 0.46 b | 1.43 ± 0.21 b | 6.23 ± 0.99 b | 57 ± 5 a | 14 ± 3 a | 0.25 ± 0.06 b |
| H + W | 81 ± 6 d | 27 ± 3 c | 4.44 ± 0.71 c | 1.44 ± 0.27 b | 4.95 ± 0.78 b | 19 ± 3 c | 8 ± 1 b | 0.43 ± 0.07 a |
| CK + Ca2+ | 104 ± 8 c | 46 ± 6 b | 7.38 ± 1.23 a | 2.31 ± 0.37 a | 13.55 ± 3.85 a | 45 ± 5 b | 1 ± 1 c | 0.03 ± 0.01 c |
| H + Ca2+ | 251 ± 8 a | 57 ± 5 b | 8.02 ± 0.56 a | 2.39 ± 0.24 a | 15.03 ± 1.71 a | 57 ± 4 a | 1 ± 0 c | 0.02 ± 0.00 c |
| Group | Fv/Fm 1 | PIABS 2 | ΨEo 3 | σRo 4 | γRC 5 | φEo 6 | φRo 7 | φDo 8 |
|---|---|---|---|---|---|---|---|---|
| CK + W | 0.80 ± 0.01 a 9 | 5.16 ± 0.44 a | 0.61 ± 0.06 a | 0.20 ± 0.04 a | 0.38 ± 0.03 a | 0.49 ± 0.06 a | 0.10 ± 0.02 bc | 0.20 ± 0.01 b |
| CK + Ca2+ | 0.80 ± 0.02 a | 5.44 ± 0.16 a | 0.63 ± 0.07 a | 0.25 ± 0.02 a | 0.37 ± 0.03 ab | 0.50 ± 0.06 a | 0.13 ± 0.01 a | 0.20 ± 0.02 b |
| H + W | 0.75 ± 0.03 b | 2.57 ± 0.47 b | 0.55 ± 0.12 a | 0.22 ± 0.09 a | 0.30 ± 0.04 b | 0.41 ± 0.10 a | 0.08 ± 0.02 c | 0.25 ± 0.03 a |
| H + Ca2+ | 0.79 ± 0.02 ab | 4.72 ± 0.97 a | 0.63 ± 0.04 a | 0.24 ± 0.02 a | 0.36 ± 0.04 ab | 0.50 ± 0.05 a | 0.12 ± 0.00 ab | 0.21 ± 0.02 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, J.; Cheng, H.; Li, X.; Pan, X.; Hu, Y.; Jin, S. Beneficial Effect of Exogenously Applied Calcium Chloride on the Anatomy and Fast Chlorophyll Fluorescence in Rhododendron × pulchrum Leaves Following Short-Term Heat Stress Treatment. Agronomy 2022, 12, 3226. https://doi.org/10.3390/agronomy12123226
Shen J, Cheng H, Li X, Pan X, Hu Y, Jin S. Beneficial Effect of Exogenously Applied Calcium Chloride on the Anatomy and Fast Chlorophyll Fluorescence in Rhododendron × pulchrum Leaves Following Short-Term Heat Stress Treatment. Agronomy. 2022; 12(12):3226. https://doi.org/10.3390/agronomy12123226
Chicago/Turabian StyleShen, Jianshuang, Hefeng Cheng, Xueqin Li, Xiangdong Pan, Yue Hu, and Songheng Jin. 2022. "Beneficial Effect of Exogenously Applied Calcium Chloride on the Anatomy and Fast Chlorophyll Fluorescence in Rhododendron × pulchrum Leaves Following Short-Term Heat Stress Treatment" Agronomy 12, no. 12: 3226. https://doi.org/10.3390/agronomy12123226