Genotype × Environment Interaction Influence Secondary Metabolite in Cowpea Infested by Flower Bud Thrips

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Sites and Experimental Procedure
2.2. Data Collection
2.2.1. Determination of Total Flavonoid Content
2.2.2. Determination of Total Phenolic Content
2.2.3. Determination of Proteins
2.2.4. Determination of Total Reducing Sugars
2.2.5. Determination of Total Tannins
2.2.6. Determination of Total Antioxidants
2.2.7. Determination of Lignin Content
2.3. Statistical Analysis
3. Results
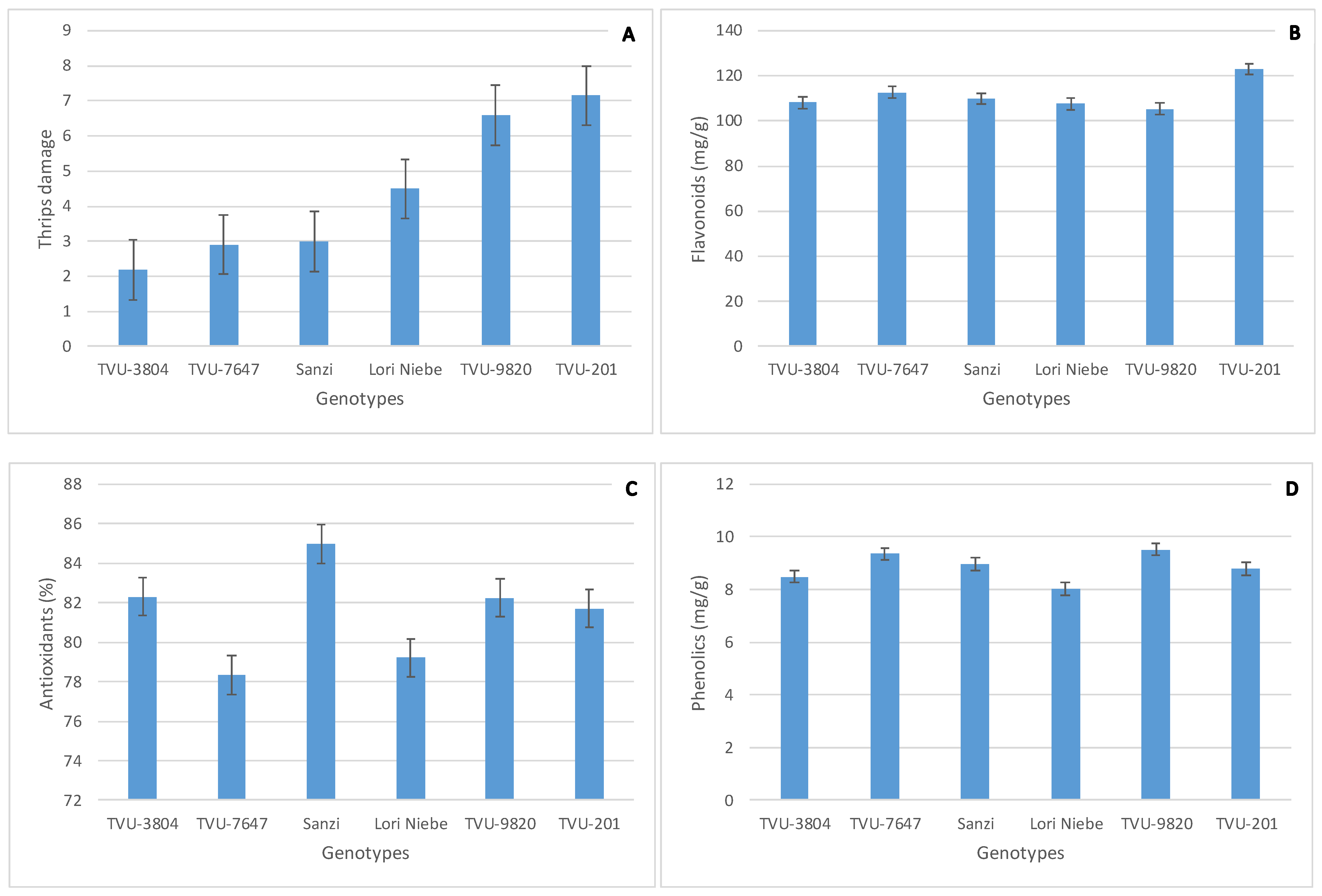
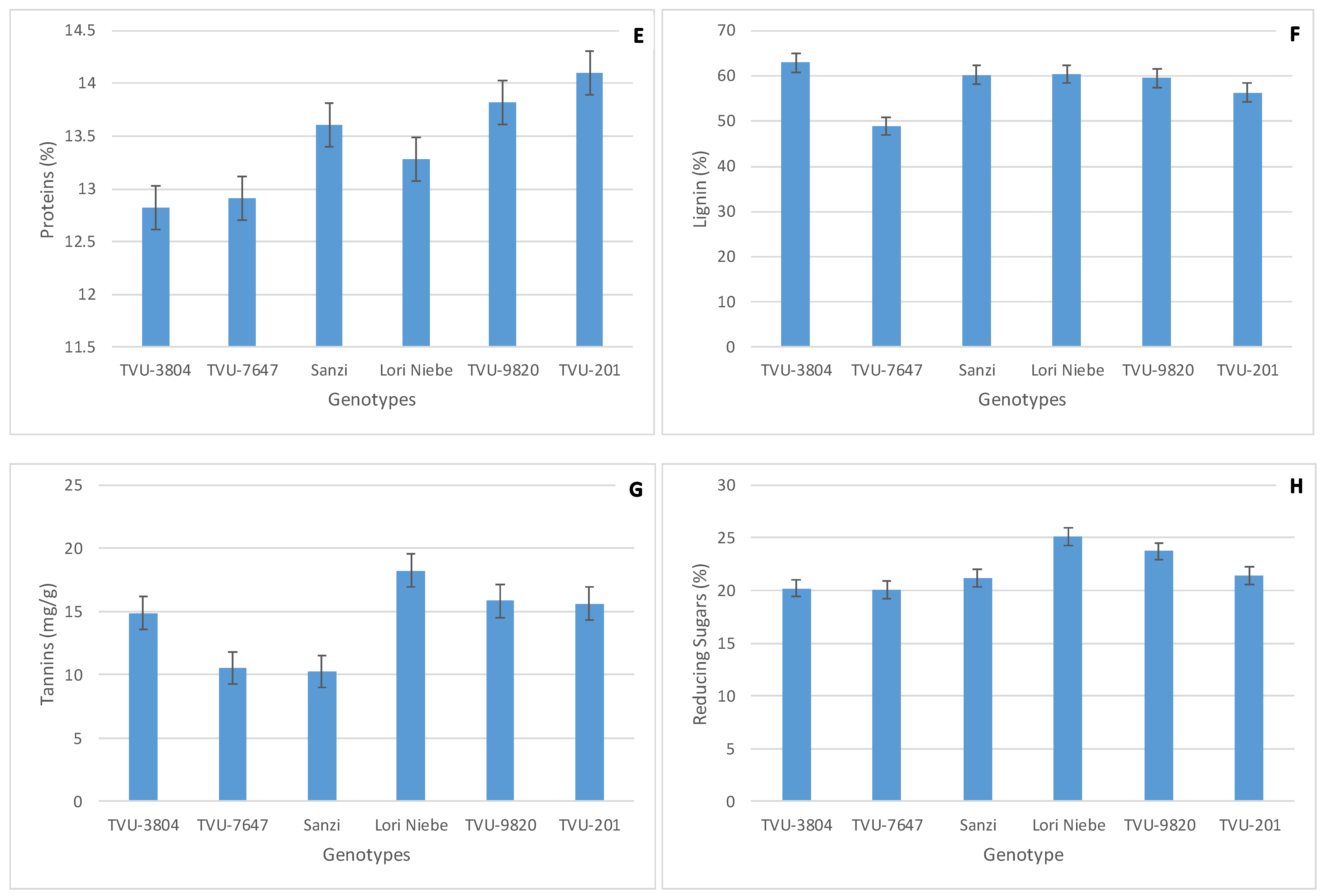
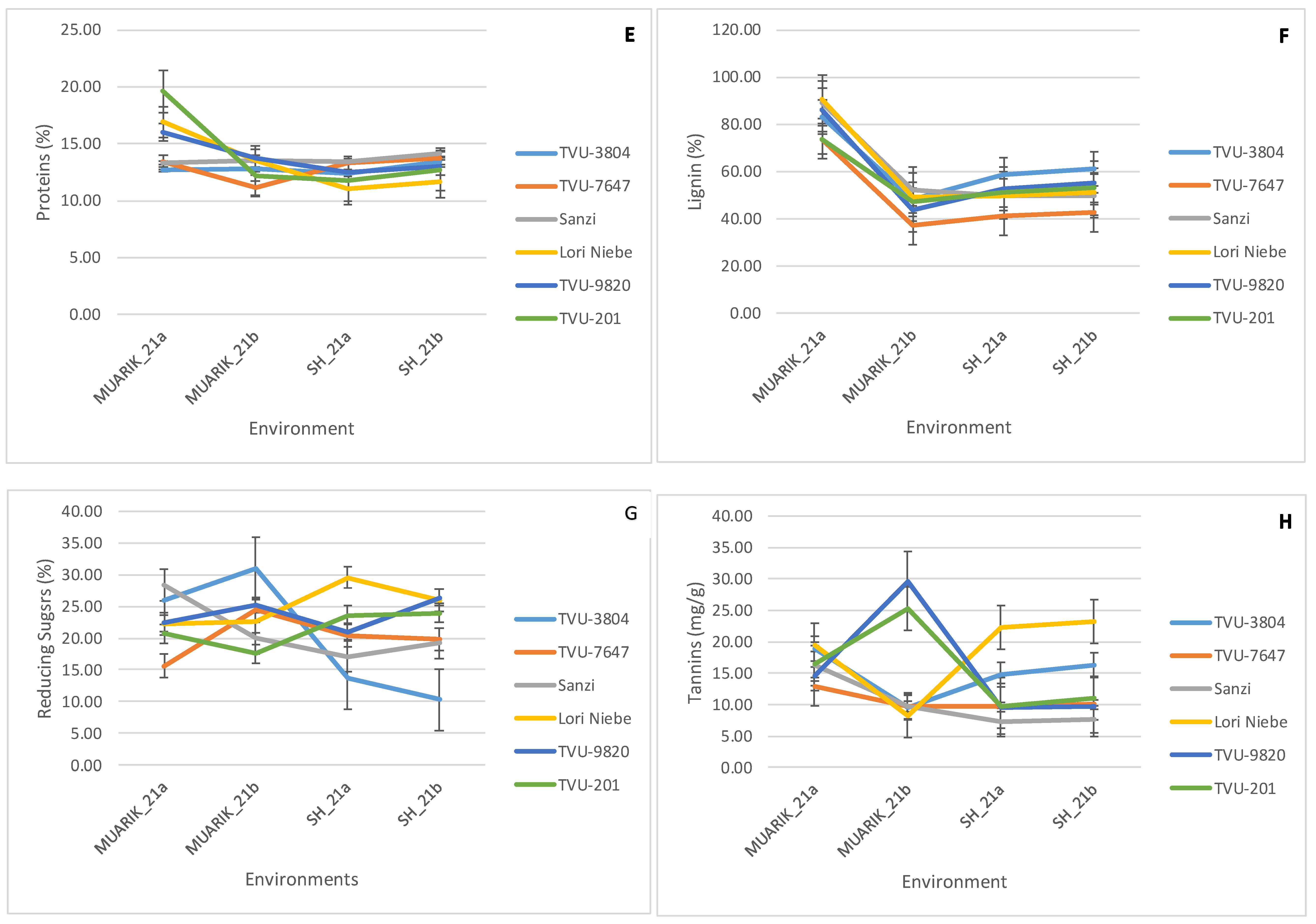
3.1. Performance of Cowpea Secondary Metabolites and Flower Thrips Damage across Environments
3.2. Correlation of Traits
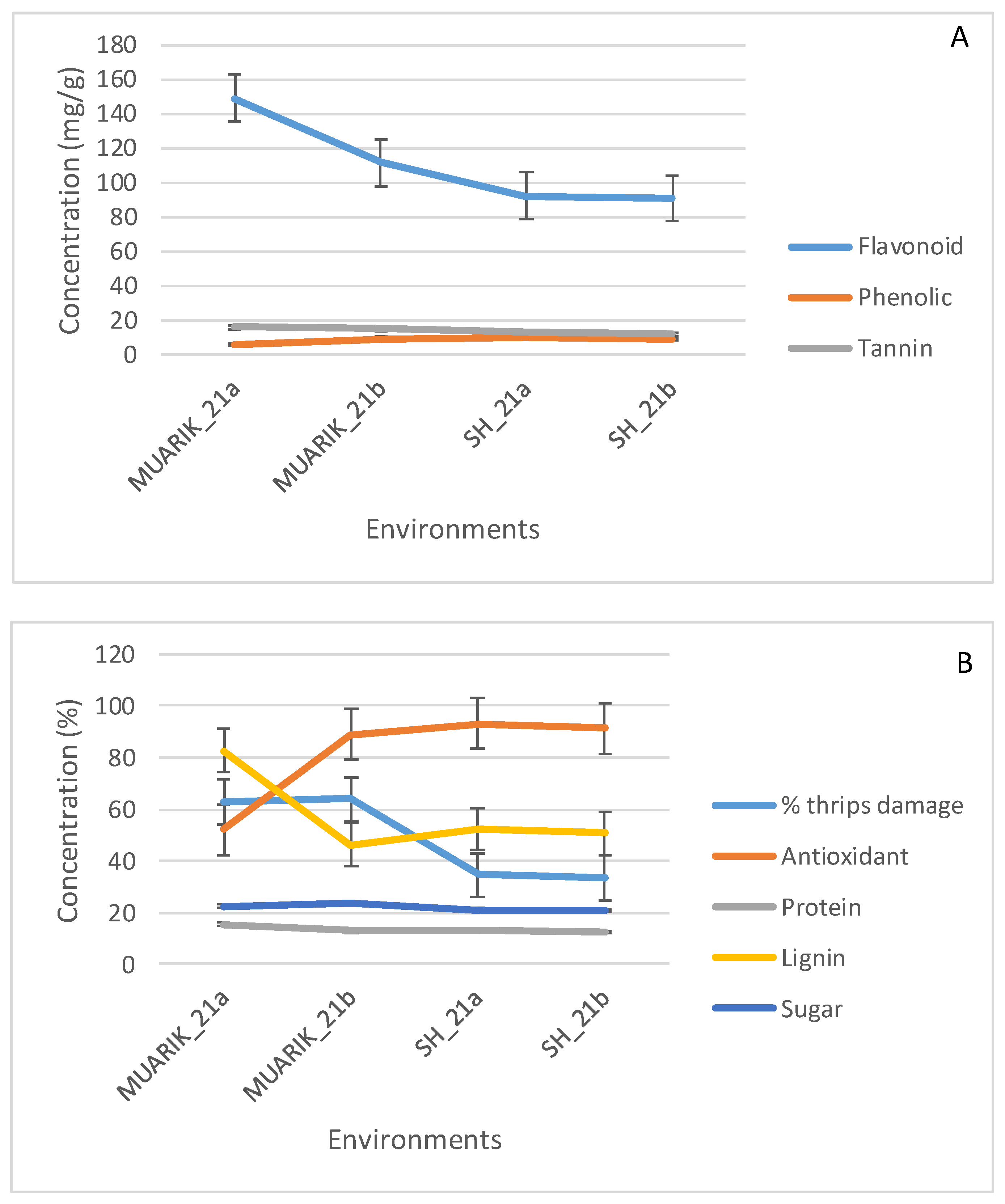
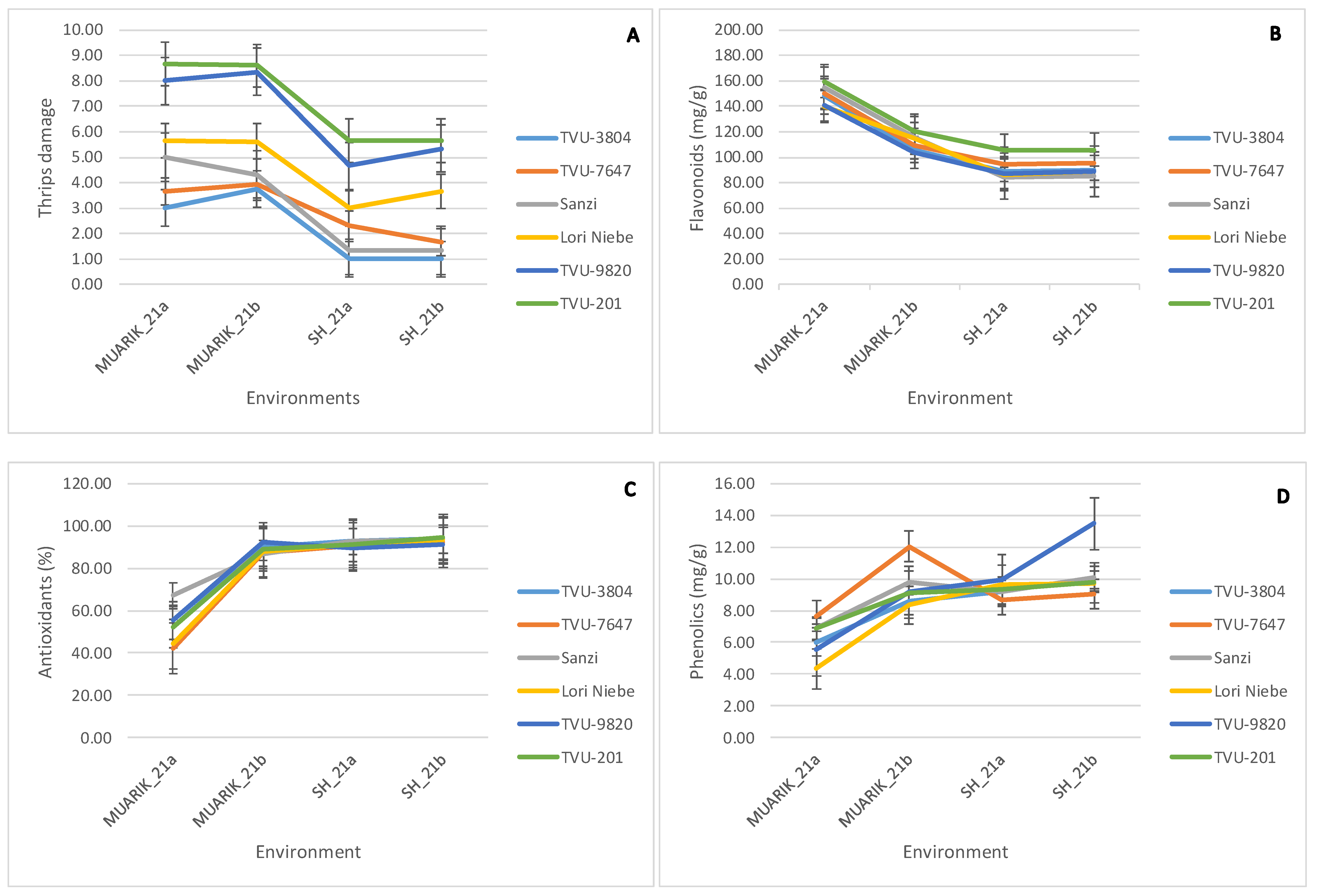
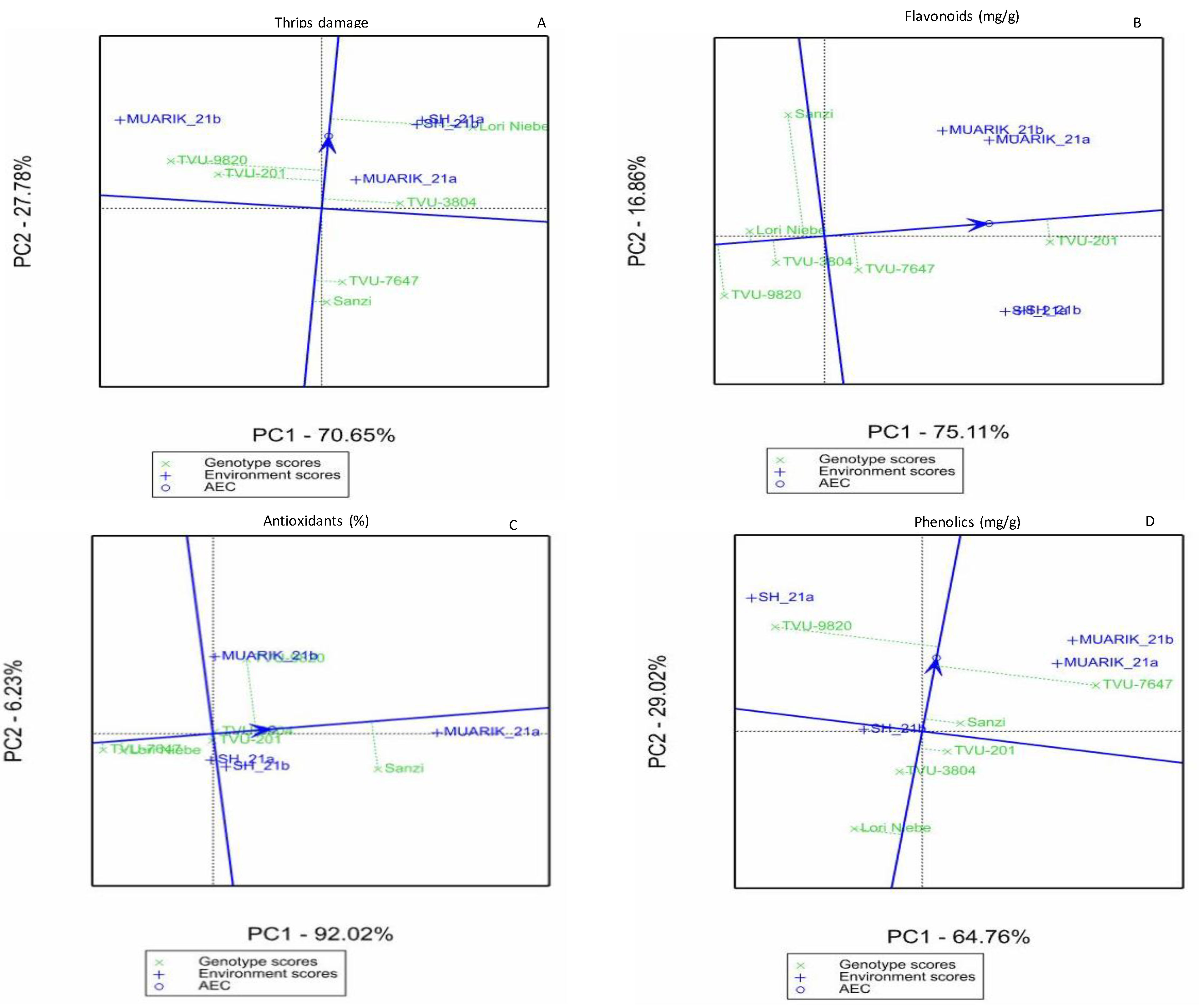
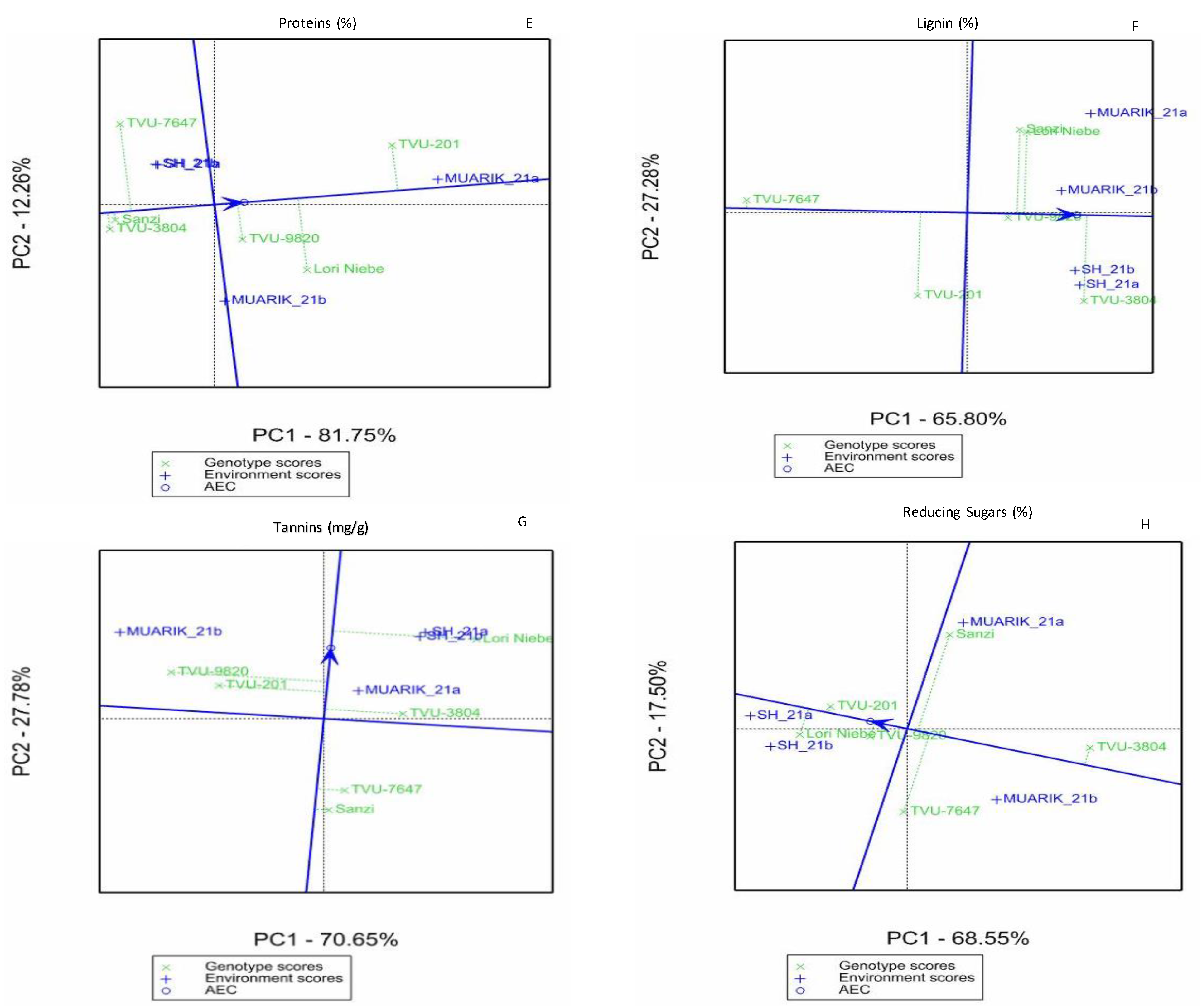
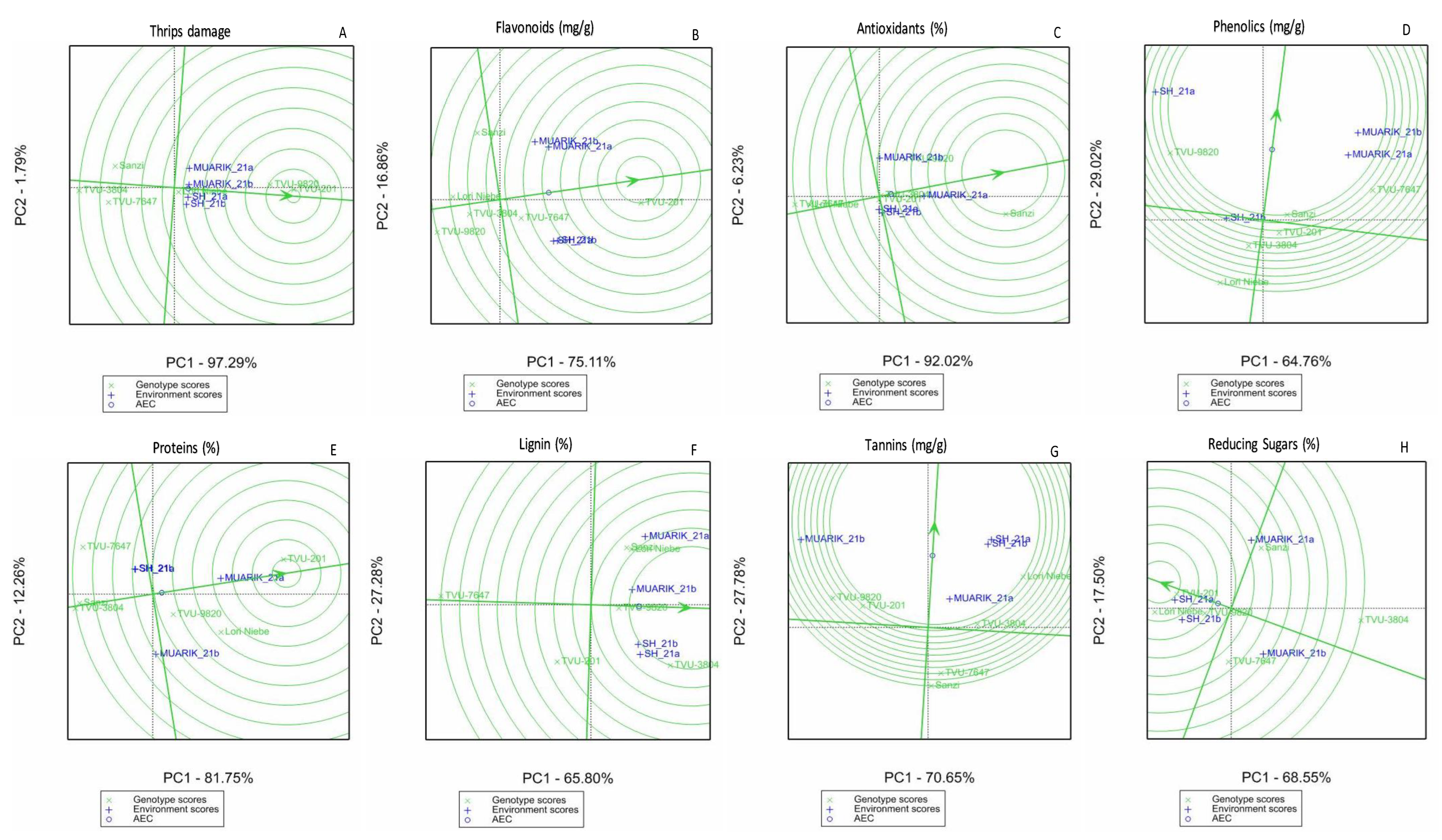
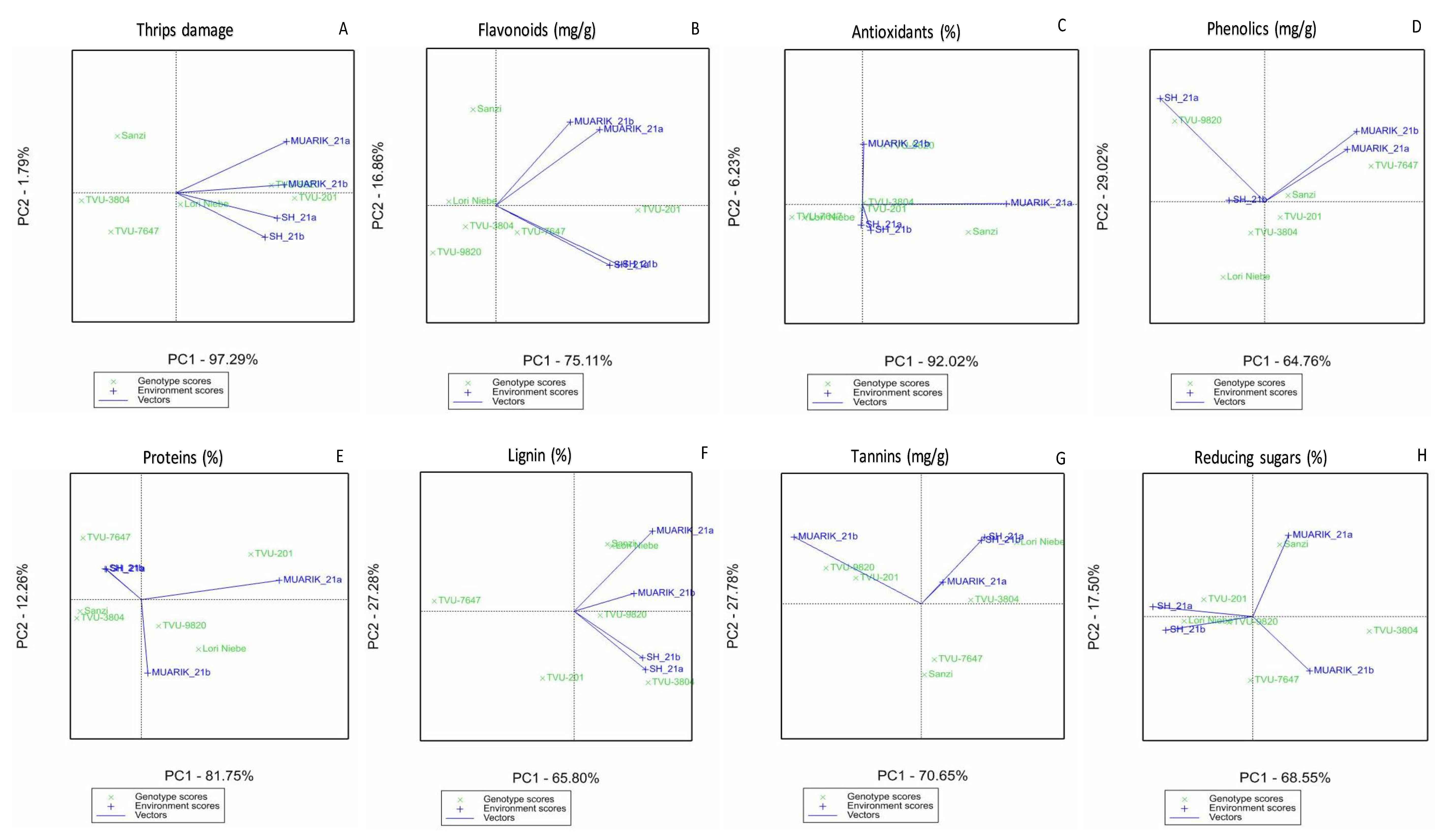
3.3. Stability of Secondary Metabolites across Environments
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Food and Agriculture Organization of the United Nations Statistical Information. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 20 September 2022).
- Santamaria, M.E.; Martínez, M.; Cambra, I.; Grbic, V.; Diaz, I. Understanding Plant Defence Responses against Herbivore Attacks: An Essential First Step towards the Development of Sustainable Resistance against Pests. Transgenic Res. 2013, 22, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Muchero, W.; Diop, N.N.; Bhat, P.R.; Fenton, R.D.; Wanamaker, S.; Pottorff, M.; Hearne, S.; Cisse, N.; Fatokun, C.; Ehlers, J.D.; et al. A Consensus Genetic Map of Cowpea [Vigna Unguiculata (L) Walp.] and Synteny Based on EST-Derived SNPs. Proc. Natl. Acad. Sci. USA 2009, 106, 18159–18164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agbahoungba, S.; Karungi, J.; Odong, T.L.; Badji, A.; Sadik, K.; Rubaihayo, P.R. Stability and Extent of Resistance of Cowpea Line to Flower Bud Thrips in Uganda. Afr. Crop. Sci. J. 2019, 3, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Alabi, A.; Omoloye, A.A.; Omoloye, A.A. Preliminary Evaluation of Selected Cowpea Varieties for Resistance to Cowpea Preliminary Evaluation of Selected Cowpea Varieties for Resistance to Cowpea Aphid, Aphis Craccivora. Niger. J. Ecol. 2012, 12, 45–55. [Google Scholar]
- Abudulai, M.; Salifu, A.B.; Haruna, M. Screening of Cowpea for Resistance to the Flower Bud Thrips, Megalurothrips Sjostedti Trybom (Thysanoptera’Thripidae). J. Appl. Sci. 2006, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Togola, A.; Boukar, O.; Chamarthi, S.; Belko, N.; Tamò, M.; Oigiangbe, N.; Ojo, J.; Ibikunle, M.; Fatokun, C. Evaluation of Cowpea Mini Core Accessions for Resistance to Flower Bud Thrips Megalurothrips Sjostedti Trybom (Thysanoptera: Thripidae). J. Appl. Entomol. 2019, 143, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Agbahoungba, S.; Karungi, J.; Odong, T.L.; Badji, A.; Kumi, F.; Mwila, N.; Rubaihayo, P.R. Biochemical Constituents Influencing the Resistance to Flower Bud Thrips in Cowpea [Vigna Unguiculata (L.) Walp] Germplasm. J. Anim. Plant Sci. 2018, 28, 128–137. [Google Scholar]
- Erb, M.; Reymond, P. Molecular Interactions between Plants and Insect Herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alabi, O.Y.; Odebiyi, J.A.; Jackai, L.E.N. Field Evaluation of Cowpea Cultivars (Vigna Unguiculata [L.] Walp.) for Resistance to Flower Bud Thrips (Megalurothrips Sjostedti Trybom) (Thysanoptera:Thripidae). Int. J. Pest. Manag. ISSN 2010, 4, 287–291. [Google Scholar] [CrossRef]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.J.; Villarroel, C.A.; Ataide, L.M.S.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and Ecological Consequences of Plant Defence Induction and Suppression in Herbivore Communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, M.A.; Burkle, L.A.; Manson, J.S.; Runyon, J.B.; Trowbridge, A.M.; Zientek, J. Global Change Effects on Plant–Insect Interactions: The Role of Phytochemistry. Curr. Opin. Insect. Sci. 2017, 23, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Barah, P.; Bones, A.M. Multidimensional Approaches for Studying Plant Defence against Insects: From Ecology to Omics and Synthetic Biology. J. Exp. Bot. 2015, 66, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Agbahoungba, S.; Karungi, J.; Odong, T.L.; Badji, A.; Sadik, K.; Rubaihayo, P.R.; Zonal, A. Stability and Extent of Resistance of Cowpea Lines to Flower Thrips. Afr. Crop. Sci. J. 2017, 25, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Alabi, O.Y.; Odebiyi, J.A.; Tamo, M.; Omoloye, A.A. The Roles of Plant Secondary Metabolites from Cowpea Floral Structures in Resistance to the Flower Bud Thrips. J. Agric. Sci. Technol. 2011, 262–269. [Google Scholar]
- Shrivastava, G.; Rogers, M.; Wszelaki, A.; Panthee, D.R.; Chen, F. Plant Volatiles-Based Insect Pest Management in Organic Farming. CRC Crit. Rev. Plant Sci. 2010, 29, 123–133. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant Immunity to Insect Herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazid, M.; Khan, T.A.; Mohammad, F. Role of Secondary Metabolites in Defense Mechanisms of Plants. Biol. Med. 2011, 3, 232–249. [Google Scholar]
- Stevens, M.T.; Waller, D.M.; Lindroth, R.L. Resistance and Tolerance in Populus Tremuloides: Genetic Variation, Costs, and Environmental Dependency. Evol. Ecol. 2007, 21, 829–847. [Google Scholar] [CrossRef]
- Ncube, B.; Finnie, J.F.; van Staden, J. Quality from the Field: The Impact of Environmental Factors as Quality Determinants in Medicinal Plants. S. Afr. J. Bot. 2012, 82, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, N.P.; Burkepile, D.E.; Parker, J.D. Variable Effects of Temperature on Insect Herbivory. PeerJ 2014, 2014, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Shukla, G.K. Some Statistical Aspects of Partitioning Genotype-Environmental Components of Variability. Heredity 1972, 29, 237–245. [Google Scholar] [CrossRef]
- Finlay, K.; Wilkinson, G. The Analysis of Adaptation in a Plant-Breeding Programme. Aust J. Agric. Res. 1963, 14, 742. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.S.; Binns, M.R. A Superiority Measure of Cultivar Performance for Cultivar × Location Data. Can. J. Plant Sci. 1988, 68, 193–198. [Google Scholar] [CrossRef]
- Ruswandi, D.; Syafii, M.; Maulana, H.; Ariyanti, M.; Indriani, N.P.; Yuwariah, Y. GGE Biplot Analysis for Stability and Adaptability of Maize Hybrids in Western Region of Indonesia. Int. J. Agron. 2021, 2021. [Google Scholar] [CrossRef]
- Horn, L.; Shimelis, H.; Sarsu, F.; Mwadzingeni, L.; Laing, M.D. Genotype-by-Environment Interaction for Grain Yield among Novel Cowpea (Vigna Unguiculata L.) Selections Derived by Gamma Irradiation. Crop. J. 2018, 6, 306–313. [Google Scholar] [CrossRef]
- Gitonga, W.H.; Ojwang, P.O.P.; Macharia, K.G.; Njau, N.P. Evaluation of Advanced Bread Wheat Genotypes for Resistance to Stem Rust and Yield Stability. Afr. J. Plant Sci. 2016, 10, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Adjei, E.A.; Esuma, W.; Alicai, T.; Chamba, E.B.; Edema, R.; Dramadri, I.O.; Ozimati, A.A.; Agaba, R.; Odong, T.L. Genotype-by-Environment Interaction of Yam (Dioscorea Species) for Yam Mosaic Virus Resistance, Dry Matter Content and Yield in Uganda. Agronomy 2022, 12, 1984. [Google Scholar] [CrossRef]
- Mbeyagala, E.K.; Ariko, J.B.; Atimango, A.O.; Amuge, E.S. Yield Stability among Cowpea Genotypes Evaluated in Different Environments in Uganda. Cogent Food Agric. 2021, 7, 1914368. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S.; Ma, B.; Woods, S.; Cornelius, P.L. GGE Biplot vs. AMMI Analysis of Genotype-by-Environment Data. Crop. Sci. 2007, 47, 643–653. [Google Scholar] [CrossRef] [Green Version]
- Odeseye, A.O.; Amusa, N.A.; Ijagbone, I.F.; Aladele, S.E.; Ogunkanmi, L.A. Genotype by Environment Interactions of Twenty Accessions of Cowpea [Vigna Unguiculata (L.) Walp.] across Two Locations in Nigeria. Ann. Agrar. Sci. 2018, 16, 481–489. [Google Scholar] [CrossRef]
- Simion, T.; Mohammed, W.; Amsalu, B. Genotype by Environment Interaction and Stability Analysis of Cowpea (Vigna Unguiculata (L) Walp) Genotypes for Yield in Ethiopia. J. Plant Breed Crop. Sci. 2018, 10, 249–257. [Google Scholar]
- Padi, F.K. Genotype × Environment Interaction and Yield Stability in a Cowpea-Based Cropping System. Euphytica 2007, 158, 11–25. [Google Scholar] [CrossRef]
- Mwila, N.; Nuwamanya, E.; Odong, T.L.; Badji, A.; Agbahoungba, S.; Ibanda, P.A.; Mwala, M.; Sohati, P.; Kyamanywa, S.; Rubaihayo, P.R. Genotype by Environment Interaction Unravels Influence on Secondary Metabolite Quality in Cassava Infested by Bemisia Tabaci. J. Agric. Sci. 2018, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Sserumaga, J.P.; Oikeh, S.O.; Mugo, S.; Otim, G.A.M.; Beyene, Y.; Abalo, G.; Kikafunda, J. Genotype by Environment Interactions and Agronomic Performance of Doubled Haploids Testcross Maize (Zea Mays L.) Hybrids. Euphytica 2015, 205, 1–15. [Google Scholar]
- Jackai, L.E.N.; Adalla, C.B. Pest Management Practices in Cowpea: A Review; IITA: Ibadan, Nigeria, 1997; pp. 240–258. [Google Scholar]
- Michalska, A.; Ceglinska, A.; Amarowicz, R.; Piskula, M.K.; Szawara-Nowak, D.; Zielinski, H. Antioxidant Contents and Antioxidative Properties of Traditional Rye Breads. J. Agric. Food Chem 2007, 55, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Mwila, N.; Rubaihayo, S.; Kyamanywa, S.; Odong, T.L.; Nuwamanya, E.; Mwala, M.; Agbahoungba, S.; Badji, A. Biochemical Factors Associated with Cassava Resistance to Whitefly Infestation. Afr Crop. Sci J. 2017, 25, 365. [Google Scholar] [CrossRef]
- Harbourne, J.B. A Guide to Modern Techniques of Plant Analysis; Kluwer Academic Publishers: Norwell, MA, USA, 1998. [Google Scholar]
- Kandakoor, S.B.; Khan, H.K.; Chakravarthy, A.K.; Kumar, C.T.A.; Venkataravana, P. Biochemical Constituents Influencing Thrips Resistance in Groundnut Germplasm. J. Environ. Biol. 2014, 35, 675–681. [Google Scholar] [PubMed]
- Ahmed, D.; Khan, M.; Saeed, R. Comparative Analysis of Phenolics, Flavonoids, and Antioxidant and Antibacterial Potential of Methanolic, Hexanic and Aqueous Extracts from Adiantum Caudatum Leaves. Antioxidants 2015, 4, 394–409. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Vilar, F.C.; Siqueira-Soares, R.D.C.; Finger-Teixeira, A.; de Oliveira, D.M.; Ferro, A.P.; da Rocha, G.J.; Ferrarese, M.D.L.L.; dos Santos, W.D.; Ferrarese-Filho, O. The Acetyl Bromide Method Is Faster, Simpler and Presents Best Recovery of Lignin in Different Herbaceous Tissues than Klason and Thioglycolic Acid Methods. PLoS ONE 2014, 9, e110000. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.L.; Felton, G.W.; Murphy, J.B.; Howles, P.A.; Dixon, R.A.; Lamb, C.J. Do Plant Phenolics Confer Resistance to Specialist and Generalist Insect Herbivores? J. Agric. Food Chem. 1997, 45, 4500–4504. [Google Scholar] [CrossRef]
- Lone, R.; Shuab, R.; Kamili, A.N. (Eds.) Plant Phenolics in Sustainable Agriculture; Springer: Singapore, 2020; ISBN 978-981-15-4889-5. [Google Scholar]
- Oomah, B.D.; Cardador-Martínez, A.; Loarca-Piña, G. Phenolics and Antioxidative Activities in Common Beans (Phaseolus Vulgaris L.). J. Sci. Food Agric. 2005, 85, 935–942. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Okoth, S.; Rose, L.J.; Ouko, A.; Netshifhefhe, N.E.I.; Sila, H.; Viljoen, A. Assessing Genotype-by-Environment Interactions in Aspergillus Ear Rot and Pre-Harvest Aflatoxin Accumulation in Maize Inbred Lines. Agronomy 2017, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Omo-ikerodah, E.; Fatokun, C.; Fawole, I. Genetic Analysis of Resistance to Flower Bud Thrips (Megalurothrips Sjostedti) Genetic Analysis of Resistance to Flower Bud Thrips (Megalurothrips Sjostedti) in Cowpea (Vigna Unguiculata L.] Walp.). Euphytica 2009, 165, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Yang, B.; Ruusunen, V.; Laaksonen, O.; Tahvonen, R.; Hellsten, J.; Kallio, H. Compositional Differences of Phenolic Compounds between Black Currant (Ribes Nigrum L.) Cultivars and Their Response to Latitude and Weather Conditions. J. Agric. Food Chem. 2012, 60, 6581–6593. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rating | Appearance |
|---|---|
| 1 | No browning/drying of stipules, leaves or flower buds; no bud abscission. |
| 3 | Initiation of browning of stipules, leaves or flower buds; no bud abscission. |
| 5 | Distinct browning/drying of stipules and leaves or flower buds; some bud abscission. |
| 7 | Serious bud abscission accompanied by browning/drying of stipules and buds; non-elongation of peduncles. |
| 9 | Very severe bud abscission, heavy browning, drying of stipules and buds; distinct non-elongation of (most or all) peduncles. |
| Environment | Temp. Max (°C) | Temp. Min (°C) | Rainfall (mm) |
|---|---|---|---|
| MUARIK_21a | 29.95 | 16.28 | 94.98 |
| MUARIK_21b | 27.19 | 15.41 | 38.85 |
| SH_21a | 33.92 | 21.34 | 99.56 |
| SH_21b | 31.03 | 20.54 | 99.56 |
| SOV | df | FT Damage | Flavonoids (mg/g) | Antioxidants (%) | Phenolics (mg/g) | Proteins (%) | Lignin (%) | Tannins (mg/g) | Reducing Sugars (%) |
|---|---|---|---|---|---|---|---|---|---|
| Rep | 2 | 0.18 | 4.00 | 7.00 | 8.50 | 4.84 | 4.00 | 4.40 | 0.80 |
| Geno | 5 | 51.58 *** | 474.00 *** | 68.00 *** | 3.70 ns | 3.11 ns | 295.00 *** | 120.42 *** | 48.64 *** |
| Env | 3 | 42.45 *** | 13164.00 *** | 6864.00 *** | 59.71 *** | 30.69 *** | 5013.00 *** | 70.38 *** | 28.35 * |
| Geno × Env | 15 | 0.59 *** | 58.00 *** | 62.00 *** | 4.57 * | 8.85 *** | 52.00 *** | 119.51 *** | 90.93 *** |
| Residuals | 46 | 0.33 | 17.00 | 13.00 | 2.07 | 1.44 | 4.00 | 2.56 | 7.52 |
| CV | 13.00 | 3.72 | 4.49 | 16.22 | 8.93 | 3.59 | 11.23 | 12.49 | |
| LSD | 0.47 | 3.40 | 3.01 | 1.18 | 0.98 | 1.71 | 1.31 | 2.25 |
| FT Damage | Flavonoids (mg/g) | Antioxidants (%) | Phenolics (mg/g) | Proteins (%) | Lignin (%) | Tannins (mg/g) | Sugars (%) | Temp.max (°C) | Temp.min (°C) | Rainfall (mm) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| FT damage | - | ||||||||||
| Flavonoids (mg/g) | 0.81 | - | |||||||||
| Antioxidants (%) | −0.62 | −0.96 ** | - | ||||||||
| Phenolics (mg/g) | −0.60 | −0.93 ** | 0.98 ** | - | |||||||
| Proteins (%) | 0.57 | 0.93 * | −0.97 ** | −0.90 * | - | ||||||
| Lignin (%) | 0.43 | 0.88 | −0.97 ** | −0.93 * | 0.97 ** | - | |||||
| Tannins (mg/g) | 0.96 ** | 0.92 * | −0.78 | −0.73 | 0.77 | 0.64 | - | ||||
| Sugars (%) | 0.96 ** | 0.62 | −0.39 | −0.38 | 0.33 | 0.17 | 0.86 | - | |||
| Temp.max (°C) | −0.81 | −0.43 | 0.23 | 0.31 | −0.06 | 0.01 | −0.62 | −0.89 | - | ||
| Temp.min (°C) | −0.99 ** | −0.74 | 0.55 | 0.56 | −0.46 | −0.34 | −0.91 * | −0.98 * | 0.89 * | - | |
| Rainfall (mm) | −0.65 | −0.09 | −0.18 | −0.17 | 0.23 | 0.40 | −0.44 | −0.84 | 0.83 | 0.72 | - |
| Rainfall (mm) | −0.65 | −0.09 | −0.18 | −0.17 | 0.23 | 0.40 | −0.44 | −0.84 | 0.83 | 0.72 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gitonga, H.W.; Kyamanywa, S.; Arusei, P.; Lukanda, M.M.; Edema, R.; Dramadri, I.O. Genotype × Environment Interaction Influence Secondary Metabolite in Cowpea Infested by Flower Bud Thrips. Agronomy 2022, 12, 3210. https://doi.org/10.3390/agronomy12123210
Gitonga HW, Kyamanywa S, Arusei P, Lukanda MM, Edema R, Dramadri IO. Genotype × Environment Interaction Influence Secondary Metabolite in Cowpea Infested by Flower Bud Thrips. Agronomy. 2022; 12(12):3210. https://doi.org/10.3390/agronomy12123210
Chicago/Turabian StyleGitonga, Hellen Wairimu, Samuel Kyamanywa, Perpetua Arusei, Musondolya Mathe Lukanda, Richard Edema, and Isaac O. Dramadri. 2022. "Genotype × Environment Interaction Influence Secondary Metabolite in Cowpea Infested by Flower Bud Thrips" Agronomy 12, no. 12: 3210. https://doi.org/10.3390/agronomy12123210