

Evaluation of In Vitro Morphogenic Response of Triticum urartu, a Donor of Au Genome of Modern Wheat

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Donor Plant Growth
2.2. Culture Media and Culture Conditions
2.3. Effect of Auxins
2.4. Initiation of Embryogenic Callus in Winter T. urartu
2.5. Statistical Analysis
3. Results
3.1. Effect of Auxins
3.1.1. The Effect of 2,4-D Concentrations
3.1.2. The Effect of Dicamba Concentrations
3.1.3. The Effect of Picloram Concentrations
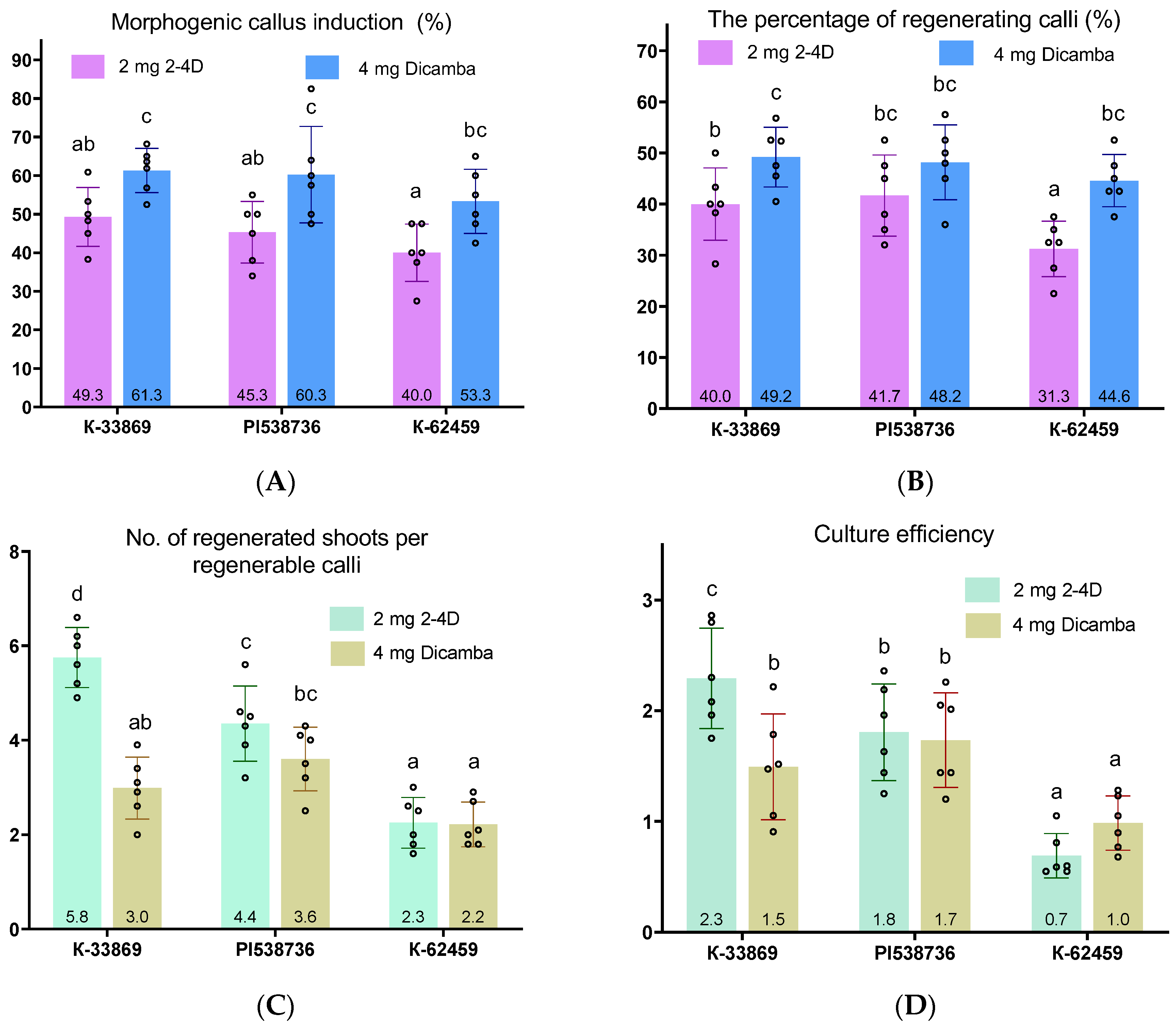
3.2. Initiation of Morphogenic Response in Winter Type of T. urartu
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dvorák, J.; Terlizzi, P.; Zhang, H.B.; Resta, P. The evolution of polyploid wheats: Identification of the A genome donor species. Genome 1993, 36, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Goncharov, N.P. Genus Triticum L. taxonomy: The present and the future. Plant Syst. Evol. 2011, 295, 1–11. [Google Scholar] [CrossRef]
- Peng, J.H.; Sun, D.; Nevo, E. Domestication evolution, genetics and genomics in wheat. Mol. Breeding 2011, 28, 281–301. [Google Scholar] [CrossRef]
- Grewal, S.; Hubbart-Edwards, S.; Yang, C.; Scholefield, D.; Ashling, S.; Burridge, A.; Wilkinson, P.A.; King, I.P.; King, J. Detection of T. urartu Introgressions in Wheat and Development of a Panel of Interspecific Introgression Lines. Front. Plant. Sci. 2018, 9, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.-Q.; Ma, B.; Shi, X.; Liu, H.; Dong, L.; Sun, H.; Cao, Y.; Gao, Q.; Zheng, S.; Li, Y.; et al. Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 2018, 557, 424–428. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hu, Y.; He, W.; Yu, K.; Zhang, C.; Li, Y.; Yang, W.; Sun, J.; Li, X.; Zheng, F.; et al. Whole-genome resequencing of the wheat A subgenome progenitor Triticum urartu provides insights into its demographic history and geographic adaptation. Plant. Commun. 2022, 3, 100345. [Google Scholar] [CrossRef]
- Dorofeev, V.F. The variability and breeding value of Armenian wheats. Euphytica 1968, 17, 451–461. [Google Scholar] [CrossRef]
- Heun, M.; Haldorsen, S.; Vollan, K. Reassessing domestication events in the Near East: Einkorn and Triticum urartu. Genome 2008, 51, 444–451. [Google Scholar] [CrossRef]
- Wang, X.; Luo, G.; Yang, W.; Li, Y.; Sun, J.; Zhan, K.; Liu, D.; Zhang, A. Genetic diversity, population structure and marker-trait associations for agronomic and grain traits in wild diploid wheat Triticum urartu. BMC Plant. Biol. 2017, 17, 112. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, C.; Alvarez, J.B. Molecular characterization of a novel waxy allele (Wx-A u 1a) from Triticum urartu Thum. ex Gandil. Genet. Resour. Crop. Evol. 2012, 59, 971–979. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, G.; Liu, D.; Wang, D.; Yang, W.; Sun, J.; Zhang, A.; Zhan, K. Genome-, Transcriptome- and Proteome-Wide Analyses of the Gliadin Gene Families in Triticum urartu. PLoS ONE 2015, 10, e0131559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talini, R.F.; Brandolini, A.; Miculan, M.; Brunazzi, A.; Vaccino, P.; Pè, M.E.; Dell’Acqua, M. Genome-wide association study of agronomic and quality traits in a world collection of the wild wheat relative Triticum urartu. Plant J. 2020, 102, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Miroshnichenko, D.; Ashin, D.; Pushin, A.; Dolgov, S. Genetic transformation of einkorn (Triticum monococcum L. ssp. monococcum L.), a diploid cultivated wheat species. BMC Biotechnol. 2018, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shi, L.; Liang, X.; Zhao, P.; Wang, W.; Liu, J.; Chang, Y.; Hiei, Y.; Yanagihara, C.; Du, L.; et al. The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 2022, 8, 110–117. [Google Scholar] [CrossRef]
- Eudes, F.; Acharya, S.; Laroche, A.; Selinger, L.B.; Cheng, K.-J. A novel method to induce direct somatic embryogenesis, secondary embryogenesis and regeneration of fertile green cereal plants. Plant. Cell Tissue Organ. Cult. 2003, 73, 147–157. [Google Scholar] [CrossRef]
- Chauhan, H.; Desai, S.A.; Khurana, P. Comparative analysis of the differential regeneration response of various genotypes of Triticum aestivum, Triticum durum and Triticum dicoccum. Plant. Cell Tissue Organ. Cult. 2007, 91, 191–199. [Google Scholar] [CrossRef]
- Alikina, O.; Chernobrovkina, M.; Dolgov, S.; Miroshnichenko, D. Tissue culture efficiency of wheat species with different genomic formulas. Crop. Breed. Appl. Biotechnol. 2016, 16, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Xu, K.; Wang, Y.; Bu, B.; Huang, W.; Sun, F.; Liu, S.; Xi, Y. Analysis of biochemical and physiological changes in wheat tissue culture using different germplasms and explant types. Acta Physiol Plant. 2015, 37, 16128. [Google Scholar] [CrossRef]
- Pellegrineschi, A.; Noguera, L.M.; Skovmand, B.; Brito, R.M.; Velazquez, L.; Salgado, M.M.; Hernandez, R.; Warburton, M.; Hoisington, D. Identification of highly transformable wheat genotypes for mass production of fertile transgenic plants. Genome 2002, 45, 421–430. [Google Scholar] [CrossRef]
- Rasco-Gaunt, S.; Riley, A.; Cannell, M.; Barcelo, P.; Lazzeri, P.A. Procedures allowing the transformation of a range of European elite wheat (Triticum aestivum L.) varieties via particle bombardment. J. Exp. Bot. 2001, 52, 865–874. [Google Scholar] [CrossRef]
- He, G.Y.; Lazzeri, P.A. Improvement of somatic embryogenesis and plant regeneration from durum wheat (Triticum turgidum var. durum Desf.) scutellum and inflorescence cultures. Euphytica 2001, 119, 369–376. [Google Scholar] [CrossRef]
- León, E.; Marín, S.; Barro, F. Improvement of in vitro culture response of elite wheat cultivars by selecting the source spike, the scutellum size and the temperature for the induction of embryogenesis. Plant. Breeding 2006, 125, 580–583. [Google Scholar] [CrossRef]
- Tao, L.-L.; Yin, G.-X.; Du, L.-P.; Shi, Z.-Y.; She, M.-Y.; Xu, H.-J.; Ye, X.-G. Improvement of Plant Regeneration from Immature Embryos of Wheat Infected by Agrobacterium tumefaciens. Agric. Sci. 2011, 10, 317–326. [Google Scholar] [CrossRef]
- Sabetta, W.; Crosatti, C.; Soltész, A.; Di Rienzo, V.; Montemurro, C. Screening Auxin Response, In Vitro Culture Aptitude and Susceptibility to Agrobacterium-Mediated Transformation of Italian Commercial Durum Wheat Varieties. Molecules 2016, 21, 1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miroshnichenko, D.; Chernobrovkina, M.; Dolgov, S. Somatic embryogenesis and plant regeneration from immature embryos of Triticum timopheevii Zhuk. and Triticum kiharae Dorof. et Migusch, wheat species with G genome. Plant. Cell Tissue Organ. Cult. 2016, 125, 495–508. [Google Scholar] [CrossRef]
- Miroshnichenko, D.; Klementyeva, A.; Dolgov, S. The Effect of Daminozide, Dark/Light Schedule and Copper Sulphate in Tissue Culture of Triticum timopheevii. Plants 2021, 10, 2620. [Google Scholar] [CrossRef]
- Wijerathna-Yapa, A.; Ramtekey, V.; Ranawaka, B.; Basnet, B.R. Applications of In Vitro Tissue Culture Technologies in Breeding and Genetic Improvement of Wheat. Plants 2022, 11, 2273. [Google Scholar] [CrossRef]
- Miroshnichenko, D.; Klementyeva, A.; Pushin, A.; Dolgov, S. A competence of embryo-derived tissues of tetraploid cultivated wheat species Triticum dicoccum and Triticum timopheevii for efficient and stable transgenesis mediated by particle inflow gun. BMC Plant. Biol. 2020, 20, 442. [Google Scholar] [CrossRef]
- Shrawat, A.K.; Armstrong, C.L. Development and Application of Genetic Engineering for Wheat Improvement. Crit. Rev. Plant Sci. 2018, 37, 335–421. [Google Scholar] [CrossRef]
- Elhiti, M.; Stasolla, C. Transduction of Signals during Somatic Embryogenesis. Plants 2022, 11, 178. [Google Scholar] [CrossRef]
- Bińka, A.; Orczyk, W.; Nadolska-Orczyk, A. The Agrobacterium-mediated transformation of common wheat (Triticum aestivum L.) and triticale (x Triticosecale Wittmack): Role of the binary vector system and selection cassettes. J. Appl. Genet. 2012, 53, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensel, G.; Marthe, C.; Kumlehn, J. Agrobacterium-Mediated Transformation of Wheat Using Immature embryos. Methods Mol. Biol. 2017, 1679, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Tsunashima, M.; Hiei, Y.; Komari, T. Wheat (Triticum aestivum L.) transformation using immature embryos. Methods Mol. Biol. 2015, 1223, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hayta, S.; Smedley, M.A.; Demir, S.U.; Blundell, R.; Hinchliffe, A.; Atkinson, N.; Harwood, W.A. An efficient and reproducible Agrobacterium-mediated transformation method for hexaploid wheat (Triticum aestivum L.). Plant Methods 2019, 15, 121. [Google Scholar] [CrossRef]
- Abe, F.; Nakamura, S.; Mori, M.; Ashikawa, I. Low-temperature pretreatment of explants and high maltose concentration during callus culture improves particle-bombardment-mediated stable transgene expression in common wheat. Plant. Biotechnol. 2020, 37, 177–184. [Google Scholar] [CrossRef]
- Qin, J.B.; Wang, Y.; Zhu, C.Q. Biolistic transformation of wheat using the HMW-GS 1Dx5 gene without selectable markers. Genet. Mol. Res. 2014, 13, 4361–4371. [Google Scholar] [CrossRef]
- Ding, L.; Li, S.; Gao, J.; Wang, Y.; Yang, G.; He, G. Optimization of Agrobacterium-mediated transformation conditions in mature embryos of elite wheat. Mol. Biol. Rep. 2009, 36, 29–36. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Concentration (mg·L−1) | Callus Induction (%) | Morphogenic Callus Formation (%) | Percentage of Regenerating Calli (%) | No. of Plantlets per Regenerable Calli | Regeneration Coefficient * |
|---|---|---|---|---|---|
| 2 | 99.7 ns | 63.3 b | 53.2 ns | 9.3 ns | 5.1 b |
| 3 | 98.6 ns | 58.3 ab | 43.9 ns | 8.6 ns | 3.9 ab |
| 4 | 99.2 ns | 47.7 a | 39.54 ns | 8.8 ns | 3.6 ab |
| 5 | 96.5 ns | 51.8 ab | 42.6 ns | 6.9 ns | 2.9 a |
| Concentration (mg·L−1) | Callus Induction (%) | Embryogenic Callus Formation (%) | Percentage of Regenerating Calli (%) | No. of Plantlets per Regenerable Calli | Regeneration Coefficient * |
|---|---|---|---|---|---|
| 2 | 100.0 ns | 69.1 a | 39.9 a | 4.0 a | 1.3 a |
| 3 | 100.0 ns | 75.5 ab | 59.0 ab | 4.6 ab | 2.7 b |
| 4 | 100.0 ns | 79.4 ab | 65.8 b | 5.7 ab | 4.0 c |
| 5 | 99.0 ns | 81.0 b | 59.9 ab | 5.9 b | 3.4 bc |
| Concentration (mg·L−1) | Callus Induction (%) | Morphogenic Callus Formation (%) | Percentage of Regenerating Calli (%) | No. of Plantlets per Regenerable Calli | Regeneration Coefficient * |
|---|---|---|---|---|---|
| 2 | 98.9 ns | 45.9 a | 30.6 a | 2.9 a | 0.9 a |
| 3 | 98.9 ns | 59.6 b | 55.4 ab | 4.9 b | 2.8 b |
| 4 | 98.9 ns | 65.4 b | 57.5 b | 5.1 b | 3.0 b |
| 5 | 100.0 ns | 59.0 b | 50.9 b | 4.8 b | 2.7 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miroshnichenko, D.; Sidorova, T.; Dolgov, S. Evaluation of In Vitro Morphogenic Response of Triticum urartu, a Donor of Au Genome of Modern Wheat. Agronomy 2022, 12, 3123. https://doi.org/10.3390/agronomy12123123
Miroshnichenko D, Sidorova T, Dolgov S. Evaluation of In Vitro Morphogenic Response of Triticum urartu, a Donor of Au Genome of Modern Wheat. Agronomy. 2022; 12(12):3123. https://doi.org/10.3390/agronomy12123123
Chicago/Turabian StyleMiroshnichenko, Dmitry, Tatiana Sidorova, and Sergey Dolgov. 2022. "Evaluation of In Vitro Morphogenic Response of Triticum urartu, a Donor of Au Genome of Modern Wheat" Agronomy 12, no. 12: 3123. https://doi.org/10.3390/agronomy12123123