
Quantitation of Multipartite Banana Bunchy Top Virus Genomic Components and Their Transcripts in Infected Tissues of Banana (Musa acuminata)

 , ,
, ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Acquisition
2.2. Primer Designing
2.3. DNA Extraction and PCR Amplification
2.4. Standard Curve Establishment
2.5. Absolute Quantitative Analysis of BBTV Components
2.6. RNA Extraction and First Strand cDNA Synthesis
2.7. Absolute Quantification of BBTV Components
2.8. Statistical Analysis
3. Results
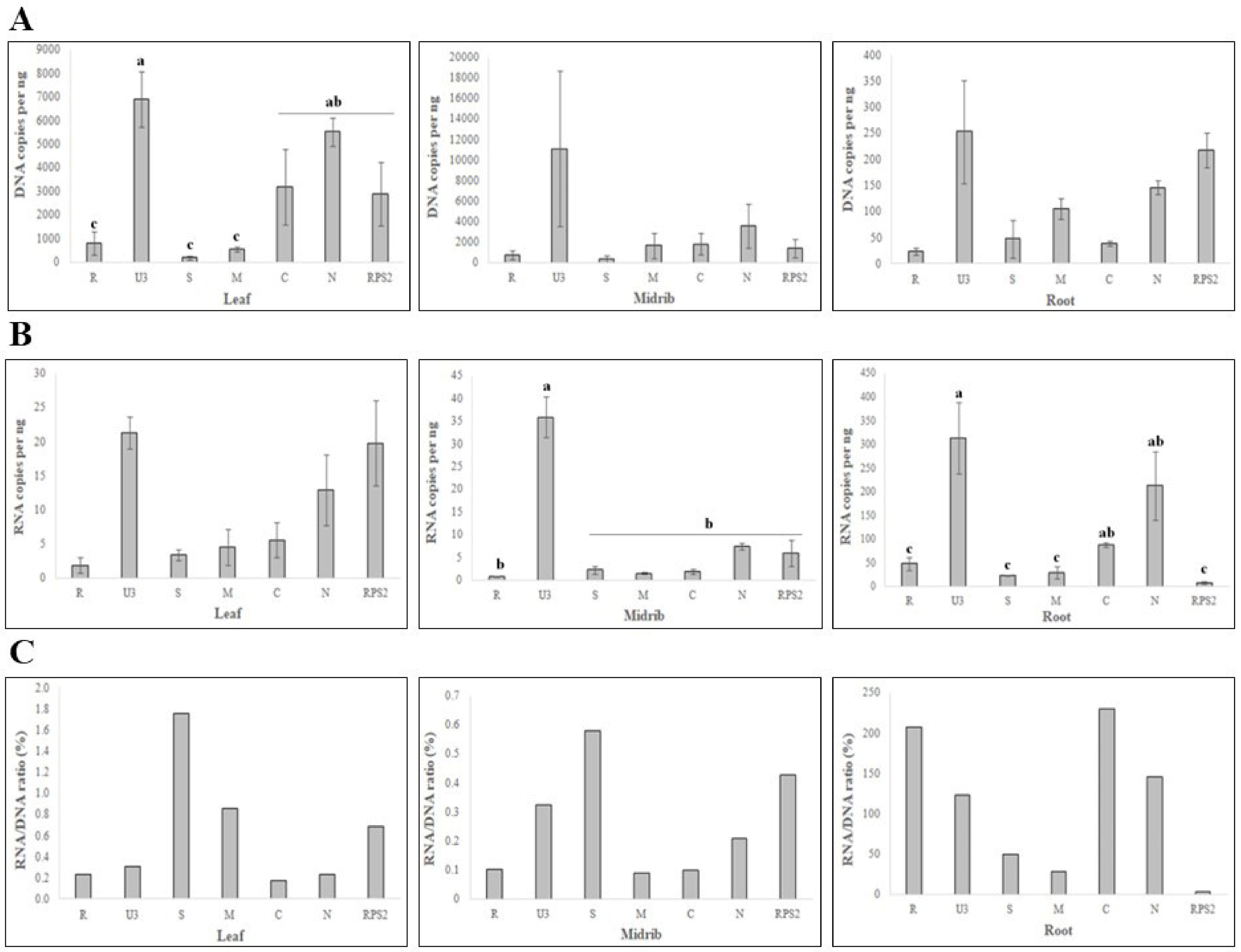
3.1. Quantitative Analysis by qPCR Showed Differential Levels of BBTV Genomic Components
3.2. Quantitative Analysis by RT-qPCR Confirmed Differential Levels of BBTV Transcripts
3.3. Independent Modulation of Transcript Levels in Each BBTV Genomic Component
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, A.; Vo, T.T.B.; Sanjaya, I.G.N.P.W.; Ho, P.T.; Kim, J.K.; Kil, E.J.; Lee, S. Nanovirus Disease Complexes: An Emerging Threat in the Modern Era. Front. Plant Sci. 2020, 11, 558403. [Google Scholar] [CrossRef] [PubMed]
- Bashir, S.; Naqvi, S.M.S.; Muhammad, A.; Hussain, I.; Ali, K.; Khan, M.R.; Farrakh, S.; Yasmin, T.; Hyder, M.Z. Banana Bunchy Top Virus Genetic Diversity in Pakistan and Association of Diversity with Recombination in Its Genomes. PLoS ONE 2022, 17, e0263875. [Google Scholar] [CrossRef] [PubMed]
- Fauquet, C.M.; Mayo, M.A.; Maniloff, J.; Desselberger, U.; Ball, L.A. Virus Taxonomy: VIIIth Report of the International Committee on Taxonomy of Viruses; Academic Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Dale, J.L. Banana Bunchy Top: An Economically Important Tropical Plant Virus Disease. Adv. Virus Res. 1987, 33, 301–325. [Google Scholar] [PubMed]
- Thomas, J.E. Banana Bunchy Top Virus (Nanoviridae). Encycl. Virol. 2021, 3, 169–175. [Google Scholar] [CrossRef]
- Karan, M.; Harding, R.M.; Dale, J.L. Association of Banana Bunchy Top Virus DNA Components 2 to 6 with Bunchy Top Disease. Mol. Plant Pathol. 1997, 7. Available online: http://www.bspp.org.uk/mppol/1997/0624karan (accessed on 23 February 2016).
- Hafner, G.J.; Stafford, M.R.; Wolter, L.C.; Harding, R.M.; Dale, J.L. Nicking and Joining Activity of Banana Bunchy Top Virus Replication Protein in Vitro. J. Gen. Virol. 1997, 78, 1795–1802. [Google Scholar] [CrossRef]
- Wanitchakorn, R.; Hafner, G.J.; Harding, R.M.; Dale, J.L. Functional Analysis of Proteins Encoded by Banana Bunchy Top Virus DNA-4 to -6. J. Gen. Virol. 2000, 81, 299–306. [Google Scholar] [CrossRef]
- Guyot, V.; Rajeswaran, R.; Chu, H.C.; Karthikeyan, C.; Laboureau, N.; Galzi, S.; Mukwa, L.F.T.; Krupovic, M.; Kumar, P.L.; Iskra-Caruana, M.L.; et al. A Newly Emerging Alphasatellite Affects Banana Bunchy Top Virus Replication, Transcription, SiRNA Production and Transmission by Aphids. PLoS Pathog. 2022, 18, e1010448. [Google Scholar] [CrossRef]
- Perales, C.; Henry, M.; Domingo, E.; Wain-Hobson, S.; Vartanian, J.-P. Lethal Mutagenesis of Foot-and-Mouth Disease Virus Involves Shifts in Sequence Space. J. Virol. 2011, 85, 12227–12240. [Google Scholar] [CrossRef] [Green Version]
- Escriu, F.; Fraile, A.; García-Arenal, F. Constraints to Genetic Exchange Support Gene Coadaptation in a Tripartite RNA Virus. PLoS Pathog. 2007, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.T.; Xie, H.M.; Zhang, Y.L.; Wang, J.H.; Xiong, Z.; Liu, Z.X. Independent Modulation of Individual Genomic Component Transcription and a Cis-Acting Element Related to High Transcriptional Activity in a Multipartite DNA Virus. BMC Genom. 2019, 20, 1–13. [Google Scholar] [CrossRef]
- Sicard, A.; Yvon, M.; Timchenko, T.; Gronenborn, B.; Michalakis, Y.; Gutierrez, S.; Blanc, S. Gene Copy Number Is Differentially Regulated in a Multipartite Virus. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motghare, M.; Kumar Dhar, A.; Kokane, A.; Warghane, A.; Kokane, S.; Kumar Sharma, A.; Krishna Reddy, M.; Kumar Ghosh, D. Quantitative Distribution of Citrus Yellow Mosaic Badnavirus in Sweet Orange (Citrus Sinensis) and Its Implication in Developing Disease Diagnostics. J. Virol. Methods 2018, 259, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Leisner, S.M.; Turgeon, R.; Howell, S.H. Long Distance Movement of Cauliflower Mosaic Virus in Infected Turnip Plants. Mol. Plant-Microbe Interact. 1992, 5, 41–47. [Google Scholar] [CrossRef]
- Leisner, S.M.; Turgeon, R. Movement of Virus and Photoassimilate in the Phloem: A Comparative Analysis. Bioessays 1993, 15, 741–748. [Google Scholar] [CrossRef]
- Hyder, M.Z.; Raza, S.Q.; Naqvi, S.M.S.; Hameed, S.; Khalid, S. Phylogenetic Relationship of TJ1 Isolate of Banana Bunchy Top Virus from Pakistan by DNA-R Sequence Analysis. Can. J. Plant Pathol. 2007, 29, 63–68. [Google Scholar] [CrossRef]
- Hyder, M.Z. Sequencing and Genetic Characterization of Major DNA Components of Banana Bunchy Top Virus. Ph.D. Thesis, PMAS-Arid Agriculture University, Rawalpindi, Pakistan, 2009. [Google Scholar]
- Dugdale, B.; Beetham, P.R.; Becker, D.K.; Harding, R.M.; Dale, J.L. Promoter Activity Associated with the Intergenic Regions of Banana Bunchy Top Virus DNA-1 to -6 in Transgenic Tobacco and Banana Cells. J. Gen. Virol. 1998, 79, 2301–2311. [Google Scholar] [CrossRef]
- Anhalt, M.D.; Almeida, R.P.P. Effect of Temperature, Vector Life Stage, and Plant Access Period on Transmission of Banana Bunchy Top Virus to Banana. Phytopathology 2008, 98, 743–748. [Google Scholar] [CrossRef] [Green Version]
- Jebakumar, R.M.; Balasubramanian, V.; Selvarajan, R. Virus Titre Determines the Efficiency of Pentalonia nigronervosa (Aphididae: Hemiptera) to Transmit Banana Bunchy Top Virus. VirusDisease 2018, 29, 499–505. [Google Scholar] [CrossRef]
- Ji, X.L.; Yu, N.T.; Qu, L.; Li, B.B.; Liu, Z.X. Banana Bunchy Top Virus (BBTV) Nuclear Shuttle Protein Interacts and Re-Distributes BBTV Coat Protein in Nicotiana Benthamiana. 3 Biotech 2019, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Krapp, S.; Greiner, E.; Amin, B.; Sonnewald, U.; Krenz, B. The Stress Granule Component G3BP Is a Novel Interaction Partner for the Nuclear Shuttle Proteins of the Nanovirus Pea Necrotic Yellow Dwarf Virus and Geminivirus Abutilon Mosaic Virus. Virus Res. 2017, 227, 6–14. [Google Scholar] [CrossRef]
- Roberts, C.A.; Dietzgen, R.G.; Heelan, L.A.; Maclean, D.J. Real-Time RT-PCR Fluorescent Detection of Tomato Spotted Wilt Virus. J. Virol. Methods 2000, 88, 1–8. [Google Scholar] [CrossRef]
- Mware, B.O. Development of Banana Bunchy Top Virus Resistance in Banana; RNAi Approach. Ph.D. Thesis, Queensland University of Technology, Brisbane, Australia, 2016. [Google Scholar]
- Lageix, S.; Catrice, O.; Deragon, J.M.; Gronenborn, B.; Pélissier, T.; Ramírez, B.C. The nanovirus-encoded Clink protein affects plant cell cycle regulation through interaction with the retinoblastoma-related protein. J. Virol. 2007, 81, 4177–4185. [Google Scholar] [CrossRef] [Green Version]
- Stranger, B.E.; Forrest, M.S.; Dunning, M.; Ingle, C.E.; Beazlsy, C.; Thorne, N.; Redon, R.; Bird, C.P.; De Grassi, A.; Lee, C.; et al. Relative Impact of Nucleotide and Copy Number Variation on Gene Phenotypes. Science 2007, 315, 848–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grafi, G.; Burnett, R.J.; Helentjaris, T.; Larkins, B.A.; Decaprio, J.A.; Sellers, W.R.; Kaelin, W.G. A Maize CDNA Encoding a Member of the Retinoblastoma Protein Family: Involvement in Endoreduplication. Proc. Natl. Acad. Sci. USA 1996, 93, 8962–8967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordman, J.; Orr-Weaver, T.L. Regulation of DNA Replication during Development. Development 2012, 139, 455–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hastings, P.J.; Lupski, J.R.; Rosenberg, S.M.; Ira, G. Mechanisms of Change in Gene Copy Number. Nat. Rev. Genet. 2009, 10, 551–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugdale, B.; Becker, D.K.; Beetham, P.R.; Harding, R.M.; Dale, J.L. Promoters Derived from Banana Bunchy Top Vires DNA-1 to-5 Direct Vascular-Associated Expression in Transgenic Banana (Musa Spp.). Plant Cell Rep. 2000, 19, 810–814. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| BBTV Genomic Component | Name of Primer | Sequence 5′-3′ Direction | Product Size (bp) |
|---|---|---|---|
| Primers for qPCR and RT-qPCR | |||
| DNA-R | DNA-R F DNA-R R | GGAAGATACAAGAATCGAAGG GGAAGGAAGTTAGCCATTAC | 567 |
| DNA-U3 | DNA-U3 F DNA-U3 R | CCGGTCAACATTATTCTGG CAGAAGAGAGAGAGCAATTATC | 140 |
| DNA-S | DNA-S F DNA-S R | GACAACAAATGGCTAGGTATC CCGTTACATTCTTCCTCAAC | 471 |
| DNA-M | DNA-M F DNA-M R | ATGGCATTAACAACAGAGC TTCCTTGCTCATCCCTTC | 325 |
| DNA-C | DNA-C F DNA-C R | CGATGTGAAGAGAGAGATTAAG GACAATCACTCCAAGAGAAC | 413 |
| DNA-N | DNA-N F DNA-N R | GGATGCGATTGGAAGAAG CATTACAGTTGTTGCTGTTAC | 373 |
| RPS2 | RPS2-F RPS2- R | AATTGGTCTACTACGGAAAC CATCTCCACGGTACATAAG | 299 |
| Primers for confirmation and sequencing | |||
| DNA-R | DNA-R F DNA-R R | ATGGCGCGATATGTGGTATG TCTGTCGTCGATGATGATCTTG | 1111 |
| DNA-U3 | DNA-U3 F DNA-U3 R | TTGTGCTGAGGCGGAAGAT CCACCTTCACAGAAGAGAG | 1062 |
| DNA-S | DNA-S F DNA-S R | GTATCCGAAGAAATCCATC CTAGCCATTTGTTGTCTG | 1075 |
| DNA-M | DNA-M F DNA-M R | ATGGCATTAACAACAGAGCG TTAGCAGGGTCCTATTTATAGG | 1046 |
| DNA-C | DNA-C F DNA-C R | GAATCGTCTGCTATGCCTG CCAGAACTCCATTTCTCTTC | 1018 |
| DNA-N | DNA-N F DNA-N R | GATGGATTGGGCGGAATCA GCTTCTGCTTTGCTTTCGC | 1088 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashir, S.; Farrakh, S.; Yasmin, T.; Muhammad, A.; Bashir, T.; Manghwar, H.; Mora-Poblete, F.; Iqbal, S.; Baazeem, A.; Hyder, M.Z. Quantitation of Multipartite Banana Bunchy Top Virus Genomic Components and Their Transcripts in Infected Tissues of Banana (Musa acuminata). Agronomy 2022, 12, 2990. https://doi.org/10.3390/agronomy12122990
Bashir S, Farrakh S, Yasmin T, Muhammad A, Bashir T, Manghwar H, Mora-Poblete F, Iqbal S, Baazeem A, Hyder MZ. Quantitation of Multipartite Banana Bunchy Top Virus Genomic Components and Their Transcripts in Infected Tissues of Banana (Musa acuminata). Agronomy. 2022; 12(12):2990. https://doi.org/10.3390/agronomy12122990
Chicago/Turabian StyleBashir, Sana, Sumaira Farrakh, Tayyaba Yasmin, Aish Muhammad, Tasmia Bashir, Hakim Manghwar, Freddy Mora-Poblete, Shehzad Iqbal, Alaa Baazeem, and Muhammad Zeeshan Hyder. 2022. "Quantitation of Multipartite Banana Bunchy Top Virus Genomic Components and Their Transcripts in Infected Tissues of Banana (Musa acuminata)" Agronomy 12, no. 12: 2990. https://doi.org/10.3390/agronomy12122990