
The Effects of Nitrogen Fertilisation on Plant Species Richness in European Permanent Grasslands: A Systematic Review and Meta-Analysis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Literature Review and Data Extraction
2.2. Meta-Analysis
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A
- For grassland: (dryland? or grass* or grazed or grazing or hayfield? or hay field? or haymeadow? or herbaceous or herbage? or meadow? or pasture? or rangeland? or range land? or ryegrass* or silvopast* or steppe or steppes or sward? or woodpast*)
- For Europe: exp europe/or (albania or andorra or austria or belarus or belgium or bosnia or british isles or bulgaria or croatia or cyprus or czech republic or czechia or czechoslovakia or denmark or england or estonia or estonian soviet socialist republic or estonian ssr or europe* or finland or flanders or france or german democratic republic or german federal republic or germany or great britain or greece or hercegovina or herzegovina or hungary or iceland or ireland or irish republic or italy or kosovo or latvia or latvian ssr or liechtenstein or lithuania or lithuanian soviet socialist republic or lithuanian ssr or luxemb?urg or macedonia or moldova or monaco or montenegro or netherlands or norway or poland or portugal or romania or san marino or scotland or serbia or slovakia or slovenia or spain or sweden or switzerland or uk or ukraine or united kingdom or wales or wallonia or walloon or yugoslavia) not (new england or new south wales)
- For plant diversity: ((plant? adj3 composition?) or (plant? adj3 diversit*) or (plant? adj3 richness) or (botanical adj3 composition? or (botanical adj3 diversit*) or botanical richness or (floristic adj3 composition?) or (floristic adj3 diversit*) or (floristic adj3 richness) or (species adj4 composition?) or (species adj3 diversit*) or (species adj3 richness) or (vegetation? adj3 composition?) or (vegetation? adj 3 richness) or biodiversity or agrobiodiversity) not (anti oxidant? or anti-oxidant? or biochemical composition or biofuel or chemical composition or extract* or medical or medicinal or pharmaceut*)
- Outside these European Natura 2000 biogeographic zones of interest: Alpine, Atlantic, Boreal, Continental, Mediterranean or Pannonian. Biogeographical boundaries are a combination of official delineations used in the Habitats Directive (92/43/EEC) and for the EMERALD Network under the Convention on the Conservation of European Wildlife and Natural Habitats (Bern Convention). They are independent of political boundaries of Emerald Network countries or EU Member States (https://www.eea.europa.eu/data-and-maps/data/biogeographical-regions-europe-3. (accessed on 1 February 2021)).
- Outside these countries in Europe: Member states of the EU-28 or Albania, Belarus, Bosnia Herzegovina, Kosovo, Macedonia, Moldova, Montenegro, Norway, Serbia, Switzerland or Ukraine.
- Unit of study was not grassland.
- Not in the English language.
- The outcome was not one of the indicators of interest.
- Papers on urban amenity grasses.
- Reviews.
- Modelling studies.
- Experiments under controlled conditions: laboratories, greenhouses or pots
- Study took place within Europe
- Study contained permanent grassland, defined as “land used to grow grasses or other herbaceous forage that has not been included in the crop rotation of the holding for a duration of five years or longer” (EU, 2004).
- Study reported a change in plant diversity as a response of nitrogen fertilisation
- Full-text, or required data could be readily obtained
- Study results were not duplicated in other records
- Article was not a review or modelling study
- Written in the English language
- Articles reported a measure of plant richness, whereby data, trends or qualitative results on these measures could be obtained from text, tables, figures or supplementary information.
- Articles included a measure of nitrogen fertilisation, whether from artificial or organic (e.g., manure) sources, whereby the total amount of nitrogen per unit area per unit time was given or could be calculated. We excluded studies of nitrogen enrichment from atmospheric deposition or from nearby sources (i.e., not applied directly to the plots being measured).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Contrast(s) | Articles (n) |
|---|---|
| Nitrogen fertilisation | 36 |
| Nitrogen fertilisation and cutting | 11 |
| Nitrogen fertilisation and grazing | 3 |
| Nitrogen fertilisation, cutting and grazing | 2 |
| Category | Rating | Rationale |
|---|---|---|
| Risk of bias (RoB) | High | We rated the overall RoB as high, mostly driven by a lack of specificity of measurements making changes in plant species richness difficult to directly attribute to the addition of nitrogen fertilisers. High heterogeneity between studies (inconsistency) make generalisations difficult. Limitations in study design and execution compared to, for example, randomised control trials, also increase the overall RoB. |
| Indirectness | High | Owing to study designs, and our aim to include studies most relevant to ‘real-world’ agricultural practice, indirectness of measurements is considered to be high. |
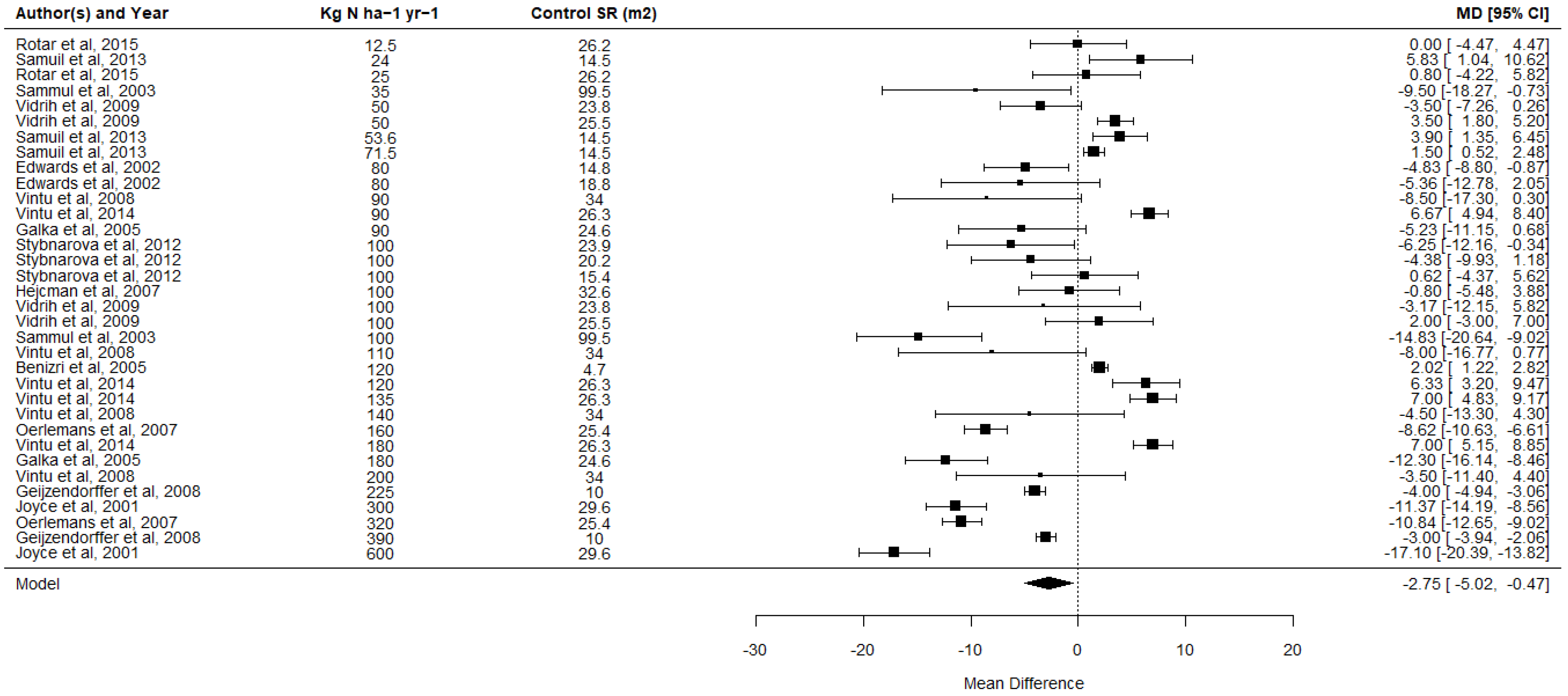
| Inconsistency | High | Figure 1 shows high heterogeneity and inconsistency between studies, making generalisations difficult and highlighting the importance of contextual factors. |
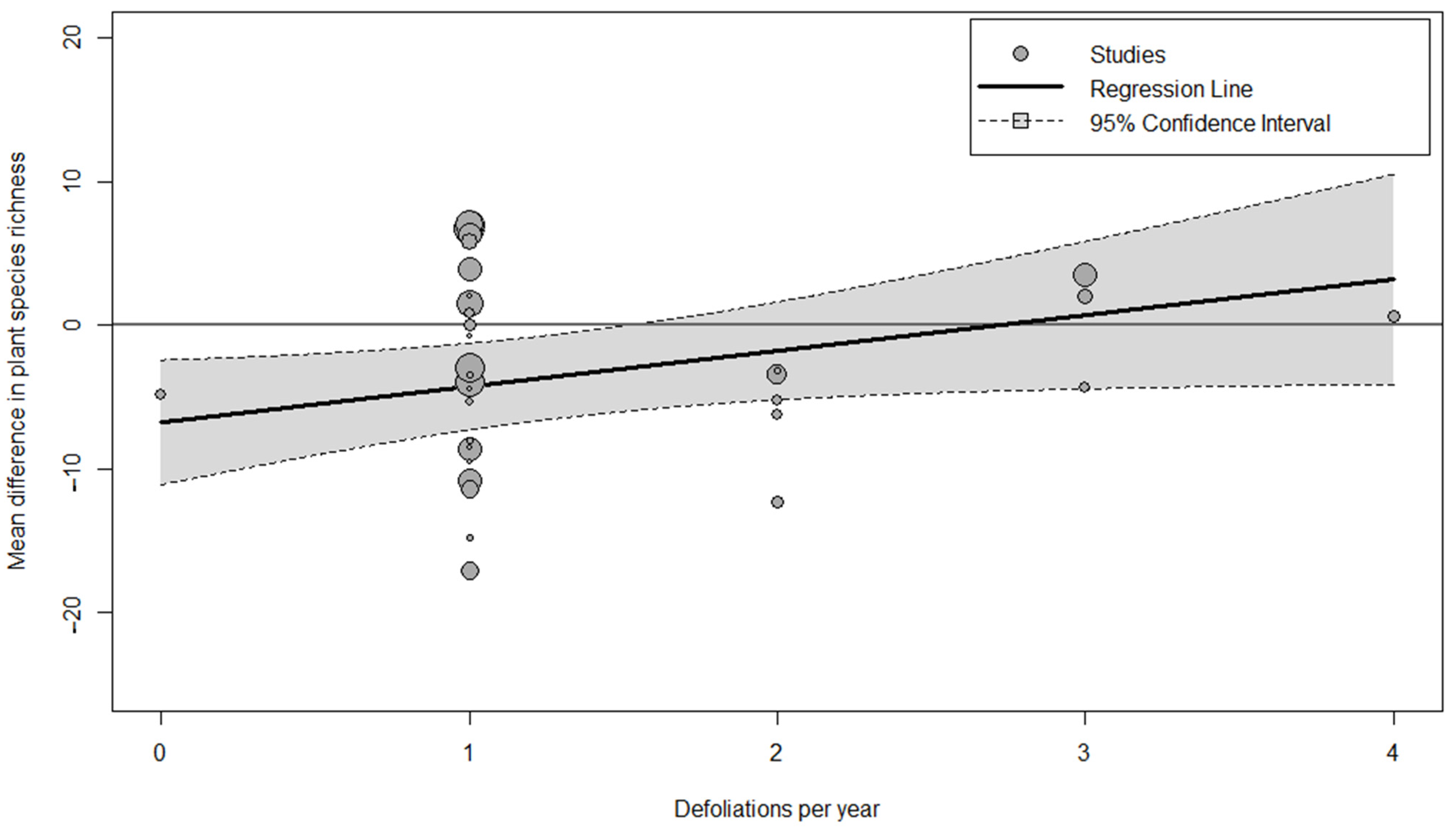
| Imprecision | High | Our findings suggest that local context, (some of which we included, i.e., defoliation, baseline richness, etc., and other factors which we did not include, i.e., soil type, landscape history) may be more important than the strength or shape of the effect of nitrogen fertilisation, which overall explained a low amount of data variation. |
| Publication bias | Low | Funnel plot examination showed no great evidence of publication bias, however it is reasonable to expect that some studies finding no significant effect (in either direction) were not published and therefore not included here. Furthermore, our review is biased towards studies published in the English language. |
| Magnitude of effect | Low | The majority of studies found a low magnitude effect, and over 40% of included effect sizes had confidence intervals that crossed 0 (i.e., no effect) |
| Overall Strength of Evidence | Low-moderate | An overall negative effect was observed, and we have reasonable confidence in this. This confidence is constrained by the number, size and quality of individual studies, high inconsistency in effects between studies, and lack of information at some scales of fertilisation and species richness. More information has a high potential to change this conclusion. |
References
- EU Commission Regulation (EC) No 796/2004 of 21 April 2004 Laying Down Detailed Rules for the Implementation of Cross-Compliance, Modulation and the Integrated Administration and Control System Provided for in of Council Regulation (EC) No 1782/2003 Establis. 2004. Available online: https://eur-lex.europa.eu/legal-content/en/ALL/?uri=CELEX%3A02004R0796-20100101 (accessed on 1 October 2022).
- Green, B.H. Agricultural intensification and the loss of habitat, species and amenity in British grasslands: A review of historical change and assessment of future prospects. Grass Forage Sci. 1990, 45, 365–372. [Google Scholar] [CrossRef]
- Lemaire, G.; Hodgson, J.; Abad, C. Grassland Productivity and Ecosystem Services; CABI: Cambridge, UK, 2011. [Google Scholar]
- Hejcman, M.; Hejcmanová, P.; Pavlů, V.; Beneš, J. Origin and history of grasslands in Central Europe—A review. Grass Forage Sci. 2013, 68, 345–363. [Google Scholar] [CrossRef]
- Van Den Pol-Van Dasselaar, A.; Bastiaansen-Aantjes, L.; Bogue, F.; O’Donovan, M.; Huyghe, C. Grassland Use in Europe: A Syllabus for Young Farmers; Inno4Grass: Versailles, France, 2019; ISBN 978-2-7592–3146-1. [Google Scholar]
- O’Mara, F.P. The role of grasslands in food security and climate change. Ann. Bot. 2012, 110, 1263–1270. [Google Scholar] [CrossRef] [Green Version]
- Habel, J.C.; Dengler, J.; Janišová, M.; Török, P.; Wellstein, C.; Wiezik, M. European grassland ecosystems: Threatened hotspots of biodiversity. Biodivers. Conserv. 2013, 22, 2131–2138. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, J.; Bullock, J.M.; O’Farrell, P.J.; O’Connor, T.; Egoh, B.; Smith, H.G.; Everson, T.; Lindborg, R.; Everson, C. Grasslands-more important for ecosystem services than you might think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Schils, R.L.M.; Bufe, C.; Rhymer, C.M.; Francksen, R.M.; Klaus, V.H.; Abdalla, M.; Milazzo, F.; Lellei-Kovács, E.; ten Berge, H.; Bertora, C.; et al. Permanent grasslands in Europe: Land use change and intensification decrease their multifunctionality. Agric. Ecosyst. Environ. 2022, 330, 107891. [Google Scholar] [CrossRef]
- Nitsch, H.; Osterburg, B.; Roggendorf, W.; Laggner, B. Cross compliance and the protection of grassland—Illustrative analyses of land use transitions between permanent grassland and arable land in German regions. Land Use Policy 2012, 29, 440–448. [Google Scholar] [CrossRef]
- Stoate, C.; Báldi, A.; Beja, P.; Boatman, N.D.; Herzon, I.; van Doorn, A.; de Snoo, G.R.; Rakosy, L.; Ramwell, C. Ecological impacts of early 21st century agricultural change in Europe—A review. J. Environ. Manag. 2009, 91, 22–46. [Google Scholar] [CrossRef] [PubMed]
- Eze, S.; Palmer, S.M.; Chapman, P.J. Soil organic carbon stock in grasslands: Effects of inorganic fertilizers, liming and grazing in different climate settings. J. Environ. Manag. 2018, 223, 74–84. [Google Scholar] [CrossRef]
- Fuller, R.M. The changing extent and conservation interest of lowland grasslands in England and Wales: A review of grassland surveys 1930–1984. Biol. Conserv. 1987, 40, 281–300. [Google Scholar] [CrossRef]
- Boch, S.; Biurrun, I.; Rodwell, J. Grasslands of Western Europe. In Encyclopedia of the world’s biomes; Goldstein, M.I., DellaSala, D.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 678–688. [Google Scholar]
- Bardgett, R.D.; Bullock, J.M.; Lavorel, S.; Manning, P.; Schaffner, U.; Ostle, N.; Chomel, M.; Durigan, G.; Fry, E.L.; Johnson, D.; et al. Combatting global grassland degradation. Nat. Rev. Earth Environ. 2021, 2, 720–735. [Google Scholar] [CrossRef]
- Perkins, G. Mind the meadows. New Sci. 2021, 252, 27. [Google Scholar] [CrossRef]
- Korevaar, H.; Sacco, D.; Ravetto Enri, S.; Lombardi, G.; Ten Berge, H.; Bufe, C.; Whittingham, M.; Smith, P.; Vanwalleghem, T.; Lellei-Kovács, E.; et al. Characterising permanent grassland-based farming systems in Europe. In Improving Sown Grasslands through Breeding and Management; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; Volume 24, pp. 164–166. [Google Scholar]
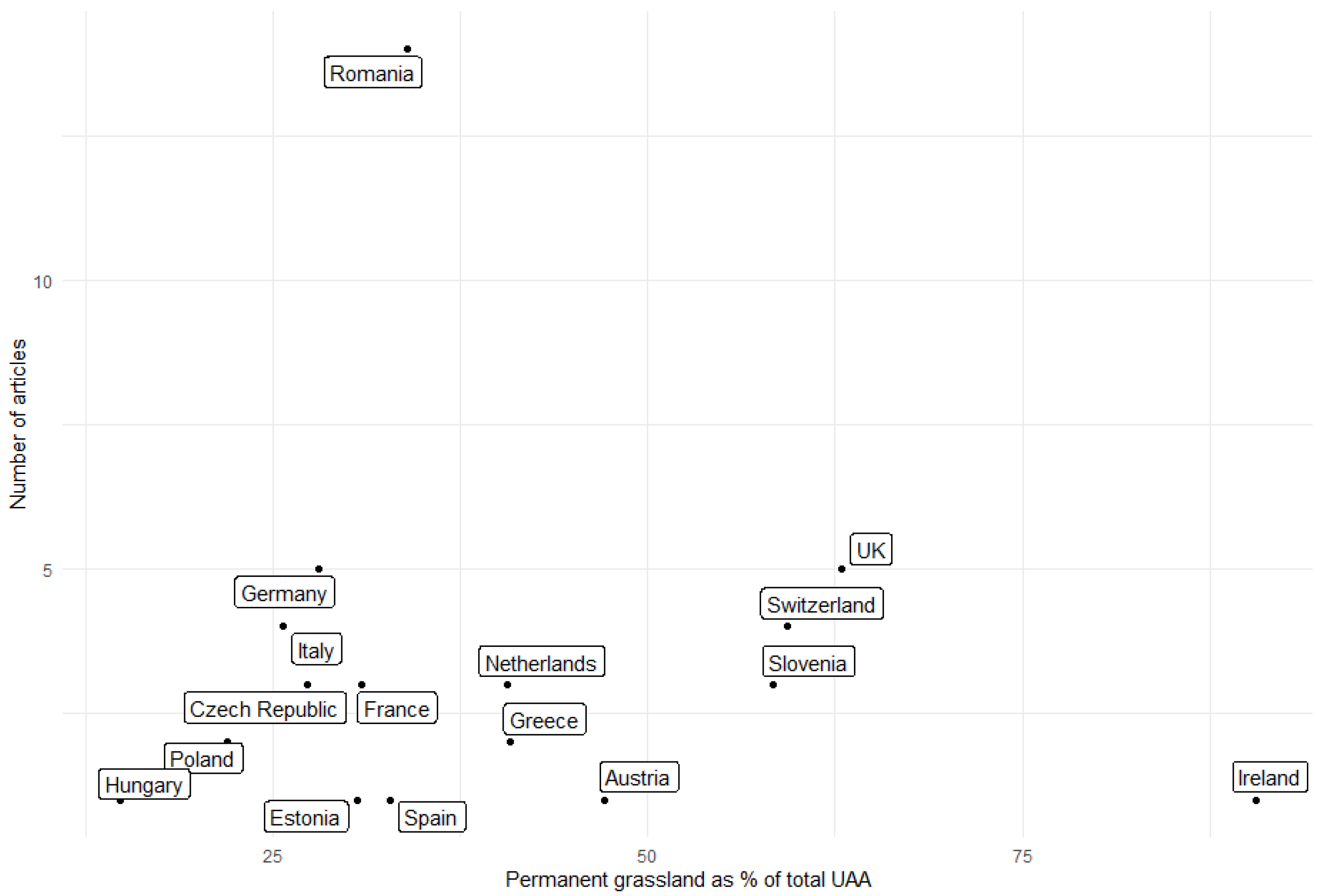
- Eurostat Share of Main Land Types in Utilised Agricultural Area (UAA) by NUTS 2 Regions. Available online: https://ec.europa.eu/eurostat/cache/metadata/en/tai05_esmsip2.htm (accessed on 1 October 2022).
- Huyghe, C.; De Vliegher, A.; Van Gils, B.; Peeters, A. Grasslands and Herbivore Production in Europe and Effects of Common Policies; Éditions Quae: Versailles, France, 2014; ISBN 978-2-7592-2157-8. [Google Scholar]
- Harpole, W.S.; Ngai, J.T.; Cleland, E.E.; Seabloom, E.W.; Borer, E.T.; Bracken, M.E.S.; Elser, J.J.; Gruner, D.S.; Hillebrand, H.; Shurin, J.B.; et al. Nutrient co-limitation of primary producer communities. Ecol. Lett. 2011, 14, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Fay, P.A.; Prober, S.M.; Harpole, W.S.; Knops, J.M.H.; Bakker, J.D.; Borer, E.T.; Lind, E.M.; MacDougall, A.S.; Seabloom, E.W.; Wragg, P.D.; et al. Grassland productivity limited by multiple nutrients. Nat. Plants 2015, 1, 15080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawley, M.J.; Johnston, A.E.; Silvertown, J.; Dodd, M.; de Mazancourt, C.; Heard, M.S.; Henman, D.F.; Edwards, G.R. Determinants of Species Richness in the Park Grass Experiment. Am. Nat. 2005, 165, 179–192. [Google Scholar] [CrossRef]
- Harpole, W.S.; Sullivan, L.L.; Lind, E.M.; Firn, J.; Adler, P.B.; Borer, E.T.; Chase, J.; Fay, P.A.; Hautier, Y.; Hillebrand, H.; et al. Addition of multiple limiting resources reduces grassland diversity. Nature 2016, 537, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Ladouceur, E.; Blowes, S.A.; Chase, J.M.; Clark, A.T.; Garbowski, M.; Alberti, J.; Arnillas, C.A.; Bakker, J.D.; Barrio, I.C.; Bharath, S.; et al. Linking changes in species composition and biomass in a globally distributed grassland experiment. Ecol. Lett. 2022. [Google Scholar] [CrossRef]
- Hautier, Y.; Niklaus, P.A.; Hector, A. Competition for Light Causes Plant Biodiversity Loss After Eutrophication. Science 2009, 324, 636–638. [Google Scholar] [CrossRef] [Green Version]
- Suding, K.N.; Collins, S.L.; Gough, L.; Clark, C.; Cleland, E.E.; Gross, K.L.; Milchunas, D.G.; Pennings, S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. USA 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [PubMed]
- Clark, C.M.; Cleland, E.E.; Collins, S.L.; Fargione, J.E.; Gough, L.; Gross, K.L.; Pennings, S.C.; Suding, K.N.; Grace, J.B. Environmental and plant community determinants of species loss following nitrogen enrichment. Ecol. Lett. 2007, 10, 596–607. [Google Scholar] [CrossRef]
- De Schrijver, A.; De Frenne, P.; Ampoorter, E.; Van Nevel, L.; Demey, A.; Wuyts, K.; Verheyen, K. Cumulative nitrogen input drives species loss in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2011, 20, 803–816. [Google Scholar] [CrossRef]
- Soons, M.B.; Hefting, M.M.; Dorland, E.; Lamers, L.P.M.; Versteeg, C.; Bobbink, R. Nitrogen effects on plant species richness in herbaceous communities are more widespread and stronger than those of phosphorus. Biol. Conserv. 2017, 212, 390–397. [Google Scholar] [CrossRef]
- Allan, E.; Manning, P.; Alt, F.; Binkenstein, J.; Blaser, S.; Blüthgen, N.; Böhm, S.; Grassein, F.; Hölzel, N.; Klaus, V.H.; et al. Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol. Lett. 2015, 18, 834–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef] [PubMed]
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Brondizio, E.S., Settele, J., Díaz, S., Ngo, H.T., Eds.; IPBES Secretariat: Bonn, Germany, 2019. [Google Scholar]
- Neyret, M.; Fischer, M.; Allan, E.; Hölzel, N.; Klaus, V.H.; Kleinebecker, T.; Krauss, J.; Le Provost, G.; Peter, S.; Schenk, N.; et al. Assessing the impact of grassland management on landscape multifunctionality. Ecosyst. Serv. 2021, 52, 101366. [Google Scholar] [CrossRef]
- Erisman, J.W.; Galloway, J.; Seitzinger, S.; Bleeker, A.; Butterbach-Bahl, K. Reactive nitrogen in the environment and its effect on climate change. Curr. Opin. Environ. Sustain. 2011, 3, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Tzilivakis, J.; Green, A.; Warner, D.; Lewis, K.A. Identification of Approaches and Measures in Action Programmes under Directive 91/676/EEC. Final Report: Report Prepared for Directorate-General Environment, European Commission, for Project ENV.D.1/SER/2018/0017 by the Agriculture and Environment Researc. 2020. Available online: https://ec.europa.eu/environment/water/water-nitrates/pdf/NAPINFO%20report%20-%20Annex%20B.pdf (accessed on 1 October 2022).
- Nitrate Pollution Prevention Regulations. No. 668. 2015. Available online: https://www.legislation.gov.uk/uksi/2015/668/contents (accessed on 1 October 2022).
- Vogt, J.; Klaus, V.; Both, S.; Fürstenau, C.; Gockel, S.; Gossner, M.; Heinze, J.; Hemp, A.; Hölzel, N.; Jung, K.; et al. Eleven years’ data of grassland management in Germany. Biodivers. Data J. 2019, 7, e36387. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.H.; Caldarone, G.; Duarte, T.K.; Ennaanay, D.; Hannahs, N.; Mendoza, G.; Polasky, S.; Wolny, S.; Daily, G.C. Integrating ecosystem-service tradeoffs into land-use decisions. Proc. Natl. Acad. Sci. USA 2012, 109, 7565–7570. [Google Scholar] [CrossRef]
- Craven, D.; Isbell, F.; Manning, P.; Connolly, J.; Bruelheide, H.; Ebeling, A.; Roscher, C.; van Ruijven, J.; Weigelt, A.; Wilsey, B.; et al. Plant diversity effects on grassland productivity are robust to both nutrient enrichment and drought. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150277. [Google Scholar] [CrossRef] [Green Version]
- Delevatti, L.M.; Cardoso, A.S.; Barbero, R.P.; Leite, R.G.; Romanzini, E.P.; Ruggieri, A.C.; Reis, R.A. Effect of nitrogen application rate on yield, forage quality, and animal performance in a tropical pasture. Sci. Rep. 2019, 9, 7596. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, A. WebPlotDigitiser. Available online: https://automeris.io/WebPlotDigitizer (accessed on 1 December 2021).
- AHDB. Nutrient Management Guide (RB209). Available online: https://ahdb.org.uk/nutrient-management-guide-rb209. (accessed on 1 December 2021).
- Dengler, J.; Matthews, T.J.; Steinbauer, M.J.; Wolfrum, S.; Boch, S.; Chiarucci, A.; Conradi, T.; Dembicz, I.; Marcenò, C.; García-Mijangos, I.; et al. Species–area relationships in continuous vegetation: Evidence from Palaearctic grasslands. J. Biogeogr. 2020, 47, 72–86. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J.; Gurevitch, J. Uses and misuses of meta-analysis in plant ecology. J. Ecol. 2014, 102, 828–844. [Google Scholar] [CrossRef]
- Assink, M.; Wibbelink, C.J.M. Fitting three-level meta-analytic models in R: A step-by-step tutorial. Quant. Methods Psychol. 2016, 12, 154–174. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 1 December 2021).
- Song, C.; Peacor, S.D.; Osenberg, C.W.; Bence, J.R. An assessment of statistical methods for nonindependent data in ecological meta-analyses. Ecology 2020, 101, e03184. [Google Scholar] [CrossRef] [PubMed]
- Viechtbauer, W. Model Selection Using the Glmulti Package. Available online: https://www.metafor-project.org/doku.php/tips:model_selection_with_glmulti (accessed on 1 January 2022).
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- O’Dea, R.E.; Lagisz, M.; Jennions, M.D.; Koricheva, J.; Noble, D.W.A.; Parker, T.H.; Gurevitch, J.; Page, M.J.; Stewart, G.; Moher, D.; et al. Preferred reporting items for systematic reviews and meta-analyses in ecology and evolutionary biology: A PRISMA extension. Biol. Rev. 2021, 96, 1695–1722. [Google Scholar] [CrossRef] [PubMed]
- Benizri, E.; Amiaud, B. Relationship between plants and soil microbial communities in fertilized grasslands. Soil Biol. Biochem. 2005, 37, 2055–2064. [Google Scholar] [CrossRef]
- Edwards, G.R.; Mitchley, J.; Tarleton, S.; Burch, F.M.; Buckley, G.P. Grassland botanical composition after 13 years of fertilizer and cutting treatments. In Multi-function grasslands: Quality forages, animal products and landscapes, Proceedings of the 19th General Meeting of the European Grassland Federation, La Rochelle, France, 27–30 May 2002; Organizing Committee of the European Grassland Federation: Versailles cedex, France, 2002; pp. 782–783. [Google Scholar]
- Štýbnarová, M.; Pozdíšek, J.; Vencálek, O.; Miˇcová, P. Effect of Fertilization and Pasture Management on Species Diversity and Forage Quality. Výzkum Chovu Skotu 2012, 54, 34–50. [Google Scholar]
- Vidrih, M.; Čop, J.; Trdan, S.; Eler, K. Changes in Floristic Composition over Three Years of Ljubljana Marsh Grassland in Relation to Cutting and Fertilising Management. Acta Agric. Slov. 2009, 93, 193. [Google Scholar]
- Vintu, V.; Samuil, C.; Sarbu, C.; Saghin, G.; Iacob, T. The Influence of Grassland Management on Biodiversity in the Mountainous Region of NE Romania. In Biodiversity and Animal Feed: Future Challenges for Grassland Production, Proceedings of the 22nd General Meeting of the European Grassland Federation, Uppsala, Sweden, 9–12 June 2008; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2008; pp. 183–185. [Google Scholar]
- Vintu, V.; Samuil, C.; Popovici, C.I.; Boureanu, C.; Stavarache, M. Management of Nardus Stricta L. and Festuca Rubra L. Grasslands in the Dorna Basin. Lucr. Ştiinţifice Ser. Agron. 2014, 57, 73–78. [Google Scholar]
- Galka, A.; Zarzynski, J.; Kopec, M. Effect of Different Fertilization Regimes on Species Composition and Habitat in Long-Term Grassland Experiment. Grassl. Sci. Eur. 2005, 10, 132–135. [Google Scholar]
- Geijzendorffer, I.R.; Schulte, R.P.O.; Finn, J.F.A.; Purvis, G. The Effect of Reduced Nitrogen Application Rates on the Botanical Diversity of Agricultural Grasslands. Tearmann Ir. J. Agri-Environ. Res. 2008, 6, 103–112. [Google Scholar]
- Hejcman, M.; Klaudisová, M.; Schellberg, J.; Honsová, D. The Rengen Grassland Experiment: Plant Species Composition after 64 Years of Fertilizer Application. Agric. Ecosyst. Environ. 2007, 122, 259–266. [Google Scholar] [CrossRef]
- Joyce, C. The Sensitivity of a Species-Rich Flood-Meadow Plant Community to Fertilizer Nitrogen: The Lužnice River Floodplain, Czech Republic. Plant Ecol. 2001, 155, 47–60. [Google Scholar] [CrossRef]
- Oerlemans, J.; von Boberfeld, W.O.; Wolf, D. Impact of Long-Term Nutrient Supply on Plant Species Diversity in Grassland: An Experimental Approach on Conventionally Used Pastures. J. Appl. Bot. Food Qual. 2012, 2, 151–157. [Google Scholar]
- Rotar, I.; Păcurar, F.; Pleșa, A.; Balázsi, Á. Low Mineral Fertilization on Grassland after 6 Years. Lucr. Ştiinţifice Ser. Agron. 2015, 2, 51–54. [Google Scholar]
- Sammul, M.; Kull, K.; Tamm, A. Clonal Growth in a Species-Rich Grassland: Results of a 20-Year Fertilization Experiment. Folia Geobot. 2003, 38, 1–20. [Google Scholar] [CrossRef]
- Samuil, C.; Vintu, V.; Sirbu, C.; Stavarache, M. Influence of Fertilizers on the Biodiversity of Semi-Natural Grassland in the Eastern Carpathians. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 195. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R.; Huyvaert, K.P. AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons. Behav. Ecol. Sociobiol. 2011, 65, 23–35. [Google Scholar] [CrossRef]
- Klaus, V.H.; Hölzel, N.; Boch, S.; Müller, J.; Socher, S.A.; Prati, D.; Fischer, M.; Kleinebecker, T. Direct and indirect associations between plant species richness and productivity in grasslands: Regional differences preclude simple generalization of productivity-biodiversity relationships. Preslia 2013, 85, 97–112. [Google Scholar]
- Boch, S.; Kurtogullari, Y.; Allan, E.; Lessard-Therrien, M.; Rieder, N.S.; Fischer, M.; Martínez De León, G.; Arlettaz, R.; Humbert, J.-Y. Effects of fertilization and irrigation on vascular plant species richness, functional composition and yield in mountain grasslands. J. Environ. Manag. 2021, 279, 111629. [Google Scholar] [CrossRef] [PubMed]
- Dullau, S.; Rydgren, K.; Kirmer, A.; Jäger, U.G.; Meyer, M.H.; Tischew, S. The Dessau Grassland Experiment—Impact of Fertilization on Forage Quality and Species Assembly in a Species-Rich Alluvial Meadow. Agriculture 2021, 11, 339. [Google Scholar] [CrossRef]
- Busch, V.; Klaus, V.H.; Schäfer, D.; Prati, D.; Boch, S.; Müller, J.; Chisté, M.; Mody, K.; Blüthgen, N.; Fischer, M.; et al. Will I stay or will I go? Plant species-specific response and tolerance to high land-use intensity in temperate grassland ecosystems. J. Veg. Sci. 2019, 30, 674–686. [Google Scholar] [CrossRef] [Green Version]
- Humbert, J.-Y.; Dwyer, J.M.; Andrey, A.; Arlettaz, R. Impacts of nitrogen addition on plant biodiversity in mountain grasslands depend on dose, application duration and climate: A systematic review. Glob. Chang. Biol. 2016, 22, 110–120. [Google Scholar] [CrossRef]
- Bonanomi, G.; Caporaso, S.; Allegrezza, M. Short-term effects of nitrogen enrichment, litter removal and cutting on a Mediterranean grassland. Acta Oecologica 2006, 30, 419–425. [Google Scholar] [CrossRef]
- Czarniecka-Wiera, M.; Kącki, Z.; Chytrý, M.; Palpurina, S. Diversity loss in grasslands due to the increasing dominance of alien and native competitive herbs. Biodivers. Conserv. 2019, 28, 2781–2796. [Google Scholar] [CrossRef] [Green Version]
- Gough, L.; Osenberg, C.W.; Gross, K.L.; Collins, S.L. Fertilization effects on species density and primary productivity in herbaceous plant communities. Oikos 2000, 89, 428–439. [Google Scholar] [CrossRef]
- Pywell, R.F.; Bullock, J.M.; Hopkins, A.; Walker, K.J.; Sparks, T.H.; Burke, M.J.W.; Peel, S. Restoration of species-rich grassland on arable land: Assessing the limiting processes using a multi-site experiment. J. Appl. Ecol. 2002, 39, 294–309. [Google Scholar] [CrossRef]
- Oelmann, Y.; Lange, M.; Leimer, S.; Roscher, C.; Aburto, F.; Alt, F.; Bange, N.; Berner, D.; Boch, S.; Boeddinghaus, R.S.; et al. Above- and belowground biodiversity jointly tighten the P cycle in agricultural grasslands. Nat. Commun. 2021, 12, 4431. [Google Scholar] [CrossRef]
- Luoto, M.; Rekolainen, S.; Aakkula, J.; Pykälä, J. Loss of Plant Species Richness and Habitat Connectivity in Grasslands Associated with Agricultural Change in Finland. AMBIO J. Hum. Environ. 2003, 32, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Humbert, J.-Y.; Pellet, J.; Buri, P.; Arlettaz, R. Does delaying the first mowing date benefit biodiversity in meadowland? Environ. Evid. 2012, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Pierik, M.; van Ruijven, J.; Bezemer, T.M.; Geerts, R.H.E.M.; Berendse, F. Recovery of plant species richness during long-term fertilization of a species-rich grassland. Ecology 2011, 92, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Midolo, G.; Alkemade, R.; Schipper, A.M.; Benítez-López, A.; Perring, M.P.; De Vries, W. Impacts of nitrogen addition on plant species richness and abundance: A global meta-analysis. Glob. Ecol. Biogeogr. 2019, 28, 398–413. [Google Scholar] [CrossRef] [Green Version]
- Stevens, C.J.; Gowing, D.J.G.; Wotherspoon, K.A.; Alard, D.; Aarrestad, P.A.; Bleeker, A.; Bobbink, R.; Diekmann, M.; Dise, N.B.; Duprè, C.; et al. Addressing the Impact of Atmospheric Nitrogen Deposition on Western European Grasslands. Environ. Manag. 2011, 48, 885–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClean, C.J.; Berg, L.J.L.; Ashmore, M.R.; Preston, C.D. Atmospheric nitrogen deposition explains patterns of plant species loss. Glob. Chang. Biol. 2011, 17, 2882–2892. [Google Scholar] [CrossRef]
- Gilliam, F.S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef] [Green Version]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef]
- Smits, N.A.C.; Bobbink, R.; Laanbroek, H.J.; Paalman, A.J.; Hefting, M.M. Repression of potential nitrification activities by matgrass sward species. Plant Soil 2010, 337, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; De Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francksen, R.M.; Turnbull, S.; Rhymer, C.M.; Hiron, M.; Bufe, C.; Klaus, V.H.; Newell-Price, P.; Stewart, G.; Whittingham, M.J. The Effects of Nitrogen Fertilisation on Plant Species Richness in European Permanent Grasslands: A Systematic Review and Meta-Analysis. Agronomy 2022, 12, 2928. https://doi.org/10.3390/agronomy12122928
Francksen RM, Turnbull S, Rhymer CM, Hiron M, Bufe C, Klaus VH, Newell-Price P, Stewart G, Whittingham MJ. The Effects of Nitrogen Fertilisation on Plant Species Richness in European Permanent Grasslands: A Systematic Review and Meta-Analysis. Agronomy. 2022; 12(12):2928. https://doi.org/10.3390/agronomy12122928
Chicago/Turabian StyleFrancksen, Richard M., Samantha Turnbull, Caroline M. Rhymer, Matthew Hiron, Conny Bufe, Valentin H. Klaus, Paul Newell-Price, Gavin Stewart, and Mark J. Whittingham. 2022. "The Effects of Nitrogen Fertilisation on Plant Species Richness in European Permanent Grasslands: A Systematic Review and Meta-Analysis" Agronomy 12, no. 12: 2928. https://doi.org/10.3390/agronomy12122928