


Identification of Genomic Loci Controlling Grain Macro and Micronutrient Variation in a Wild Barley (Hordeum vulgare spp. spontaneum) Diversity Panel
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Micronutrient Analysis
2.3. Statistical Analysis
2.4. Genotyping Association Analysis
2.5. Candidate Gene Identification
3. Results and Discussion
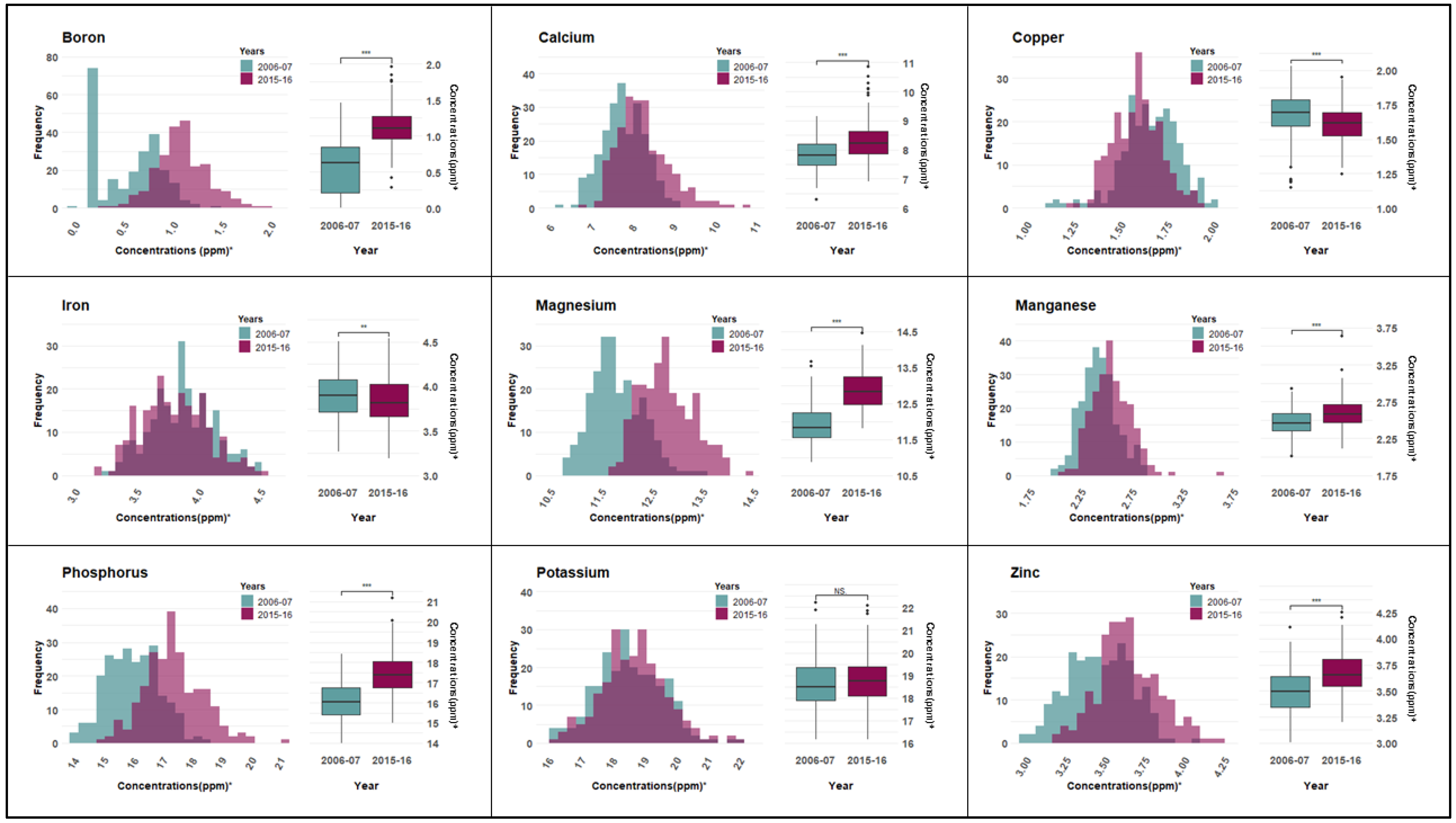
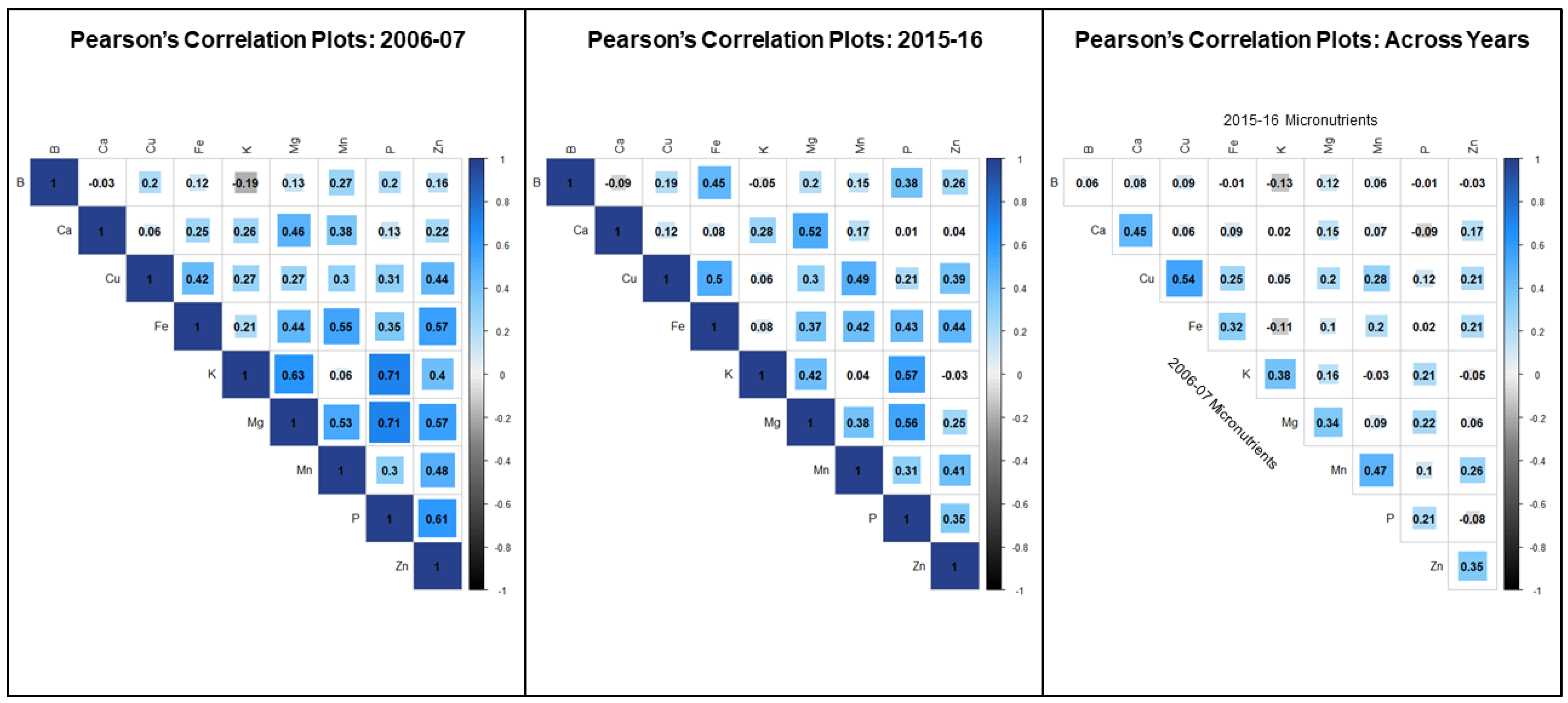
3.1. Macro and Micronutrient Content and Correlation Analysis
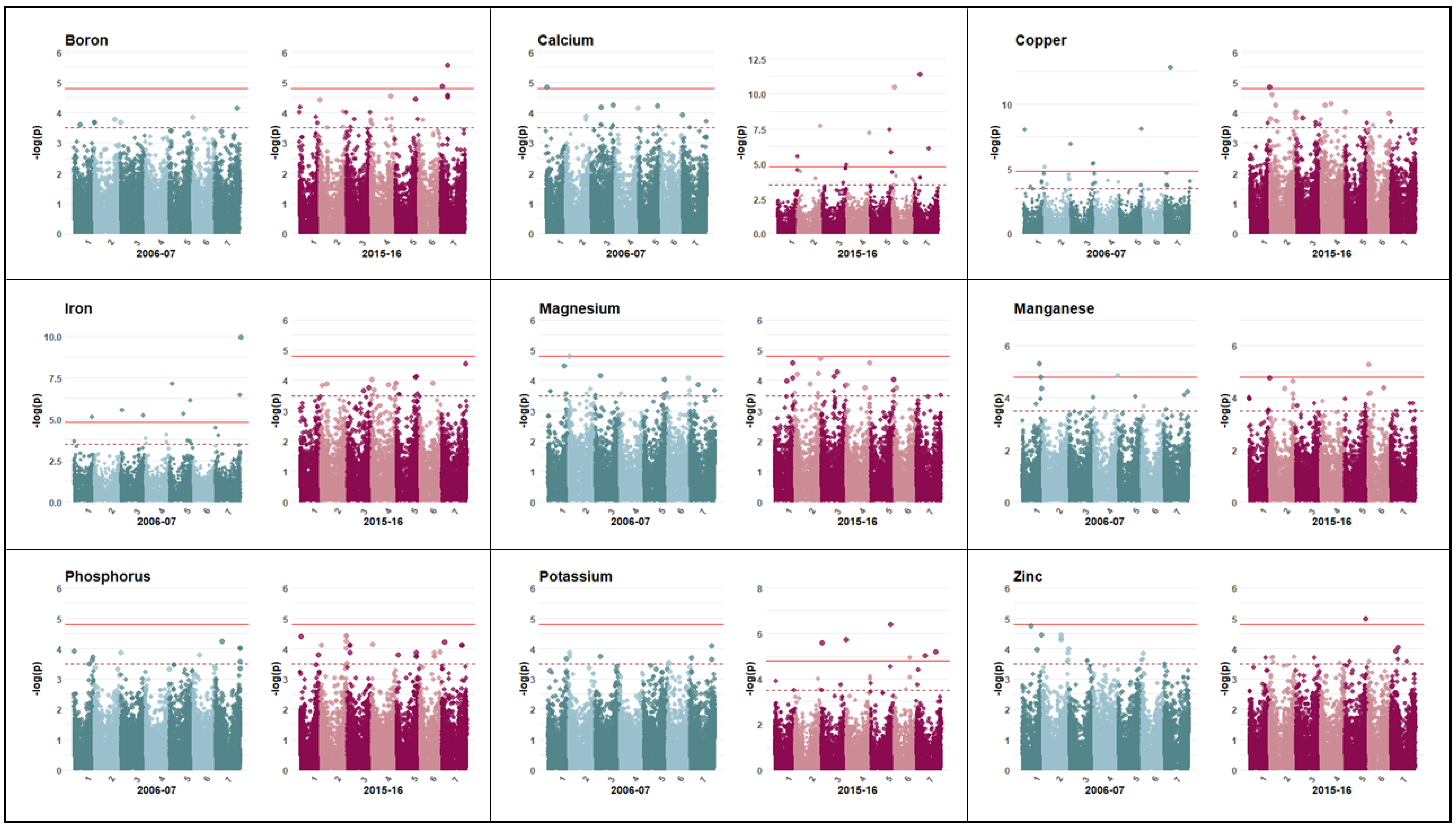
3.2. Marker Trait Association
3.3. Primary Macronutrients: Phosphorus and Potassium
3.4. Secondary Macronutrients: Calcium and Magnesium
3.5. Micronutrients: Boron, Manganese, Copper, Iron, and Zinc
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Badr, A.; Müller, K.; Schäfer-Pregl, R.; El Rabey, H.; Effgen, S.; Ibrahim, H.H.; Pozzi, C.; Rohde, W.; Salamini, F. On the Origin and Domestication History of Barley (Hordeum vulgare). Mol. Biol. Evol. 2000, 17, 499–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 2 August 2022).
- Grando, S.; Macpherson, H.G. Food Barley: Importance, Uses and Local Knowledge. In Proceedings of the International Workshop on Food Barley Improvement, Hammamet, Tunisia, 14–17 January 2002; ICARDA: Aleppo, Syria, 2005. [Google Scholar]
- Åman, P.; Newman, C.W. Chemical Composition of Some Different Types of Barley Grown in Montana, U.S.A. J. Cereal Sci. 1986, 4, 133–141. [Google Scholar] [CrossRef]
- The World Is at a Critical Juncture. Available online: https://www.fao.org/state-of-food-security-nutrition/2021/en/ (accessed on 26 August 2022).
- The Concept of the Global Hunger Index. Available online: https://www.globalhungerindex.org/about.html (accessed on 20 June 2022).
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The Essential Metals for Humans: A Brief Overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Centeno, V.; de Barboza, G.D.; Marchionatti, A.; Rodríguez, V.; Tolosa de Talamoni, N. Molecular Mechanisms Triggered by Low-Calcium Diets. Nutr. Res. Rev. 2009, 22, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayod, M.; Tyerman, S.D.; Leigh, R.A.; Gilliham, M. Calcium Storage in Plants and the Implications for Calcium Biofortification. Protoplasma 2010, 247, 215–231. [Google Scholar] [CrossRef]
- Pravina, P.; Sayaji, D.; Avinash, M. Calcium and Its Role in Human Body. Int. J. Res. 2013, 4, 659–668. [Google Scholar]
- Rude, R.K. Magnesium Deficiency: A Cause of Heterogeneous Disease in Humans. J. Bone Miner. Res. 1998, 13, 749–758. [Google Scholar] [CrossRef]
- Gröber, U.; Schmidt, J.; Kisters, K. Magnesium in Prevention and Therapy. Nutrients 2015, 7, 8199–8226. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I. Magnesium in Crop Production, Food Quality and Human Health. Plant Soil 2013, 368, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hänsch, R.; Mendel, R.R. Physiological Functions of Mineral Micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for Micronutrients in Staple Food Crops from a Human Nutrition Perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef]
- Vos, T.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 328 Diseases and Injuries for 195 Countries, 1990–2016: A Systematic Analysis. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Nutrition Targets 2025: Policy Brief Series. Available online: https://www.who.int/nutrition/publications/globaltargets2025_policybrief_anaemia/en (accessed on 28 September 2022).
- Hambidge, K.M.; Walravens, P.A. Disorders of Mineral Metabolism. Clin. Gastroenterol. 1982, 11, 87–117. [Google Scholar] [CrossRef]
- Farahat, M.H.; Sharaf, M.A. Hematological and Biochemical Parameter Alteration after Plateletpheresis Donation. Egypt. J. Haematol. 2016, 41, 174. [Google Scholar] [CrossRef]
- Cakmak, I.; Yazici, A.M. Others Magnesium: A Forgotten Element in Crop Production. Better Crops 2010, 94, 23–25. [Google Scholar]
- Baloch, F.S.; Nadeem, M.A. Unlocking the Genomic Regions Associated with Seed Protein Contents in Turkishcommon Bean Germplasm through Genome–Wide Association Study. Turk. J. Agric. For. 2022, 46, 113–128. [Google Scholar] [CrossRef]
- Cayton, M.T.C. Boron Toxicity in Rice; IRRI Research Paper Series; International Rice Research Institute: Manila, Philippines, 1985; Volume 113. Available online: https://pdf.usaid.gov/pdf_docs/PNABC298.pdf (accessed on 27 September 2022).
- Chatterjee, M.; Tabi, Z.; Galli, M.; Malcomber, S.; Buck, A.; Muszynski, M.; Gallavotti, A. The Boron Efflux Transporter ROTTEN EAR Is Required for Maize Inflorescence Development and Fertility. Plant Cell 2014, 26, 2962–2977. [Google Scholar] [CrossRef] [Green Version]
- Sutton, T.; Baumann, U.; Hayes, J.; Collins, N.C.; Shi, B.-J.; Schnurbusch, T.; Hay, A.; Mayo, G.; Pallotta, M.; Tester, M.; et al. Boron-Toxicity Tolerance in Barley Arising from Efflux Transporter Amplification. Science 2007, 318, 1446–1449. [Google Scholar] [CrossRef]
- Shao, J.F.; Yamaji, N.; Liu, X.W.; Yokosho, K.; Shen, R.F.; Ma, J.F. Preferential Distribution of Boron to Developing Tissues Is Mediated by the Intrinsic Protein OsNIP3. Plant Physiol. 2018, 176, 1739–1750. [Google Scholar] [CrossRef] [Green Version]
- Diehn, T.A.; Bienert, M.D.; Pommerrenig, B.; Liu, Z.; Spitzer, C.; Bernhardt, N.; Fuge, J.; Bieber, A.; Richet, N.; Chaumont, F.; et al. Boron Demanding Tissues of Brassica Napus Express Specific Sets of Functional Nodulin26-like Intrinsic Proteins and BOR1 Transporters. Plant J. 2019, 100, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Bienert, M.D.; von Wirén, N.; Bienert, G.P. Genome-Wide Association Mapping Identifies HvNIP2;2/HvLsi6 Accounting for Efficient Boron Transport in Barley. Physiol. Plant. 2021, 171, 809–822. [Google Scholar] [CrossRef]
- Bienert, G.P.; Bienert, M.D.; Jahn, T.P.; Boutry, M.; Chaumont, F. Solanaceae XIPs Are Plasma Membrane Aquaporins That Facilitate the Transport of Many Uncharged Substrates. Plant J. 2011, 66, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Pedas, P.; Ytting, C.K.; Fuglsang, A.T.; Jahn, T.P.; Schjoerring, J.K.; Husted, S. Manganese Efficiency in Barley: Identification and Characterization of the Metal Ion Transporter HvIRT1. Plant Physiol. 2008, 148, 455–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longnecker, N.E.; Marcar, N.E.; Graham, R.D. Increased Manganese Content of Barley Seeds Can Increase Grain Yield in Manganese-Deficient Conditions. Aust. J. Agric. Res. 1991, 42, 1065–1074. [Google Scholar] [CrossRef]
- Burnell, J.N. The Biochemistry of Manganese in Plants. In Manganese in Soils and Plants: Proceedings of the International Symposium on ‘Manganese in Soils and Plants’ Held at the Waite Agricultural Research Institute, The University of Adelaide, Glen Osmond, South Australia, August 22–26, 1988 as an Australian Bicentennial Event; Graham, R.D., Hannam, R.J., Uren, N.C., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 125–137. ISBN 9789400928176. [Google Scholar]
- Marschner, H. Mineral Nutrition of Higher Plants. 2nd. 1995. Available online: https://www.scirp.org›reference›ReferencesPapers (accessed on 28 September 2022).
- Puig, S.; Andrés-Colás, N.; García-Molina, A.; Peñarrubia, L. Copper and Iron Homeostasis in Arabidopsis: Responses to Metal Deficiencies, Interactions and Biotechnological Applications. Plant Cell Environ. 2007, 30, 271–290. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of Crops with Seven Mineral Elements Often Lacking in Human Diets--Iron, Zinc, Copper, Calcium, Magnesium, Selenium and Iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Shariatipour, N.; Heidari, B. Genetic-Based Biofortification of Staple Food Crops to Meet Zinc and Iron Deficiency-Related Challenges. Plant Micronutr. 2020, 173–223. [Google Scholar] [CrossRef]
- Mikula, K.; Izydorczyk, G.; Skrzypczak, D.; Mironiuk, M.; Moustakas, K.; Witek-Krowiak, A.; Chojnacka, K. Controlled Release Micronutrient Fertilizers for Precision Agriculture–A Review. Sci. Total Environ. 2020, 712, 136365. [Google Scholar] [CrossRef]
- Lowe, N.M. The Global Challenge of Hidden Hunger: Perspectives from the Field. Proc. Nutr. Soc. 2021, 80, 283–289. [Google Scholar] [CrossRef]
- Saltzman, A.; Birol, E.; Bouis, H.E.; Boy, E.; De Moura, F.F.; Islam, Y.; Pfeiffer, W.H. Biofortification: Progress toward a More Nourishing Future. Glob. Food Secur. 2013, 2, 9–17. [Google Scholar] [CrossRef]
- Bouis, H.E.; Hotz, C.; McClafferty, B. Biofortification: A New Tool to Reduce Micronutrient Malnutrition. Food Nutr. 2011, 32 (Suppl. 1), S31–S40. [Google Scholar] [CrossRef]
- Cakmak, I.; Kutman, U.B. Agronomic Biofortification of Cereals with Zinc: A Review. Eur. J. Soil Sci. 2018, 69, 172–180. [Google Scholar] [CrossRef]
- Bashir, K.; Takahashi, R.; Nakanishi, H.; Nishizawa, N.K. The Road to Micronutrient Biofortification of Rice: Progress and Prospects. Front. Plant Sci. 2013, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Verma, V.; Kaur, M.; Shivay, Y.S.; Nisar, S.; Gaber, A.; Brestic, M.; Barek, V.; et al. Biofortification—A Frontier Novel Approach to Enrich Micronutrients in Field Crops to Encounter the Nutritional Security. Molecules 2022, 27, 1340. [Google Scholar] [CrossRef]
- Laurie, S.; Faber, M.; Adebola, P.; Belete, A. Biofortification of Sweet Potato for Food and Nutrition Security in South Africa. Food Res. Int. 2015, 76, 962–970. [Google Scholar] [CrossRef]
- Jha, A.B.; Warkentin, T.D. Biofortification of Pulse Crops: Status and Future Perspectives. Plants 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Thabet, S.G.; Alomari, D.Z.; Brinch-Pedersen, H.; Alqudah, A.M. Genetic Analysis toward More Nutritious Barley Grains for a Food Secure World. Bot. Stud. 2022, 63, 6. [Google Scholar] [CrossRef]
- Detterbeck, A.; Nagel, M.; Rensch, S.; Weber, M.; Börner, A.; Persson, D.P.; Schjoerring, J.K.; Christov, V.; Clemens, S. The Search for Candidate Genes Associated with Natural Variation of Grain Zn Accumulation in Barley. Biochem. J. 2019, 476, 1889–1909. [Google Scholar] [CrossRef]
- Gyawali, S.; Otte, M.L.; Chao, S.; Jilal, A.; Jacob, D.L.; Amezrou, R.; Verma, R.P.S. Genome Wide Association Studies (GWAS) of Element Contents in Grain with a Special Focus on Zinc and Iron in a World Collection of Barley (Hordeum Vulgare L.). J. Cereal Sci. 2017, 77, 266–274. [Google Scholar] [CrossRef]
- Mamo, B.E.; Barber, B.L.; Steffenson, B.J. Genome-Wide Association Mapping of Zinc and Iron Concentration in Barley Landraces from Ethiopia and Eritrea. J. Cereal Sci. 2014, 60, 497–506. [Google Scholar] [CrossRef]
- Sallam, A.H.; Tyagi, P.; Brown-Guedira, G.; Muehlbauer, G.J.; Hulse, A.; Steffenson, B.J. Genome-Wide Association Mapping of Stem Rust Resistance in Hordeum vulgare Subsp. Spontaneum. G3 Genes Genomes Genet. 2017, 7, 3491–3507. [Google Scholar] [CrossRef] [Green Version]
- Roy, J.K.; Smith, K.P.; Muehlbauer, G.J.; Chao, S.; Close, T.J.; Steffenson, B.J. Association Mapping of Spot Blotch Resistance in Wild Barley. Mol. Breed. 2010, 26, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.J.; Gyenis, L.; Hayes, P.M.; Matus, I.; Smith, K.P.; Steffenson, B.J.; Muehlbauer, G.J. Quantitative Trait Loci for Multiple Disease Resistance in Wild Barley. Crop Sci. 2005, 45, 2563–2572. [Google Scholar] [CrossRef]
- Ames, N.; Dreiseitl, A.; Steffenson, B.J.; Muehlbauer, G.J. Mining Wild Barley for Powdery Mildew Resistance. Plant Pathol. 2015, 64, 1396–1406. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, R.; Sallam, A.H.; Steffenson, B.J.; Fiedler, J.D.; Walling, J.G. Genome-Wide Association Analysis of Natural Variation in Seed Tocochromanols of Barley. Plant Genome 2020, 13, e20039. [Google Scholar] [CrossRef]
- Walling, J.G.; Sallam, A.H.; Steffenson, B.J.; Henson, C.; Vinje, M.A.; Mahalingam, R. Quantitative Trait Loci Impacting Grain Β-glucan Content in Wild Barley (Hordeum Vulgare ssp. Spontaneum) Reveals Genes Associated with Cell Wall Modification and Carbohydrate Metabolism. Crop Sci. 2022, 62, 1213–1227. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 8 April 2022).
- Goh, L.; Yap, V.B. Effects of Normalization on Quantitative Traits in Association Test. BMC Bioinform. 2009, 10, 415. [Google Scholar] [CrossRef] [Green Version]
- Taiyun Corrplot; Github. Available online: https://github.com/taiyun/corrplot (accessed on 10 September 2022).
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J.; et al. A Chromosome Conformation Capture Ordered Sequence of the Barley Genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Navrátilová, P.; Toegelová, H.; Tulpová, Z.; Kuo, Y.-T.; Stein, N.; Doležel, J.; Houben, A.; Šimková, H.; Mascher, M. Prospects of Telomere-to-Telomere Assembly in Barley: Analysis of Sequence Gaps in the MorexV3 Reference Genome. Plant Biotechnol. J. 2022, 20, 1373–1386. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.M.; Zaitlen, N.A.; Wade, C.M.; Kirby, A.; Heckerman, D.; Daly, M.J.; Eskin, E. Efficient Control of Population Structure in Model Organism Association Mapping. Genetics 2008, 178, 1709–1723. [Google Scholar] [CrossRef] [Green Version]
- Rosyara, U.R.; De Jong, W.S.; Douches, D.S.; Endelman, J.B. Software for Genome-Wide Association Studies in Autopolyploids and Its Application to Potato. Plant Genome 2016, 9, plantgenome2015.08.0073. [Google Scholar] [CrossRef]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z. GAPIT Version 3: Boosting Power and Accuracy for Genomic Association and Prediction. Genom. Proteom. Bioinform. 2021, 19, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.K.F.; Rowe, H.C.; Kliebenstein, D.J. Understanding the Evolution of Defense Metabolites in Arabidopsis Thaliana Using Genome-Wide Association Mapping. Genetics 2010, 185, 991–1007. [Google Scholar] [CrossRef] [Green Version]
- Pasam, R.K.; Sharma, R.; Malosetti, M.; van Eeuwijk, F.A.; Haseneyer, G.; Kilian, B.; Graner, A. Genome-Wide Association Studies for Agronomical Traits in a World Wide Spring Barley Collection. BMC Plant Biol. 2012, 12, 16. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zaitlen, N.A.; Goddard, M.E.; Visscher, P.M.; Price, A.L. Advantages and Pitfalls in the Application of Mixed-Model Association Methods. Nat. Genet. 2014, 46, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Kinsella, R.J.; Kähäri, A.; Haider, S.; Zamora, J.; Proctor, G.; Spudich, G.; Almeida-King, J.; Staines, D.; Derwent, P.; Kerhornou, A.; et al. Ensembl BioMarts: A Hub for Data Retrieval across Taxonomic Space. Database 2011, 2011, bar030. [Google Scholar] [CrossRef]
- Perera, L.; Freudenthal, B.D.; Beard, W.A.; Shock, D.D.; Pedersen, L.G.; Wilson, S.H. Requirement for Transient Metal Ions Revealed through Computational Analysis for DNA Polymerase Going in Reverse. Proc. Natl. Acad. Sci. USA 2015, 112, E5228–E5236. [Google Scholar] [CrossRef] [Green Version]
- Droppa, M.; Horváth, G. The Role of Copper in Photosynthesis. CRC Crit. Rev. Plant Sci. 1990, 9, 111–123. [Google Scholar] [CrossRef]
- Smith, D.B.; Cannon, W.F.; Woodruff, L.G.; Solano, F.; Kilburn, J.E.; Fey, D.L. USGS Data Series 801: Geochemical and Mineralogical Data for Soils of the Conterminous United States. Available online: https://pubs.usgs.gov/ds/801/ (accessed on 21 October 2022).
- U.S. Department of Agriculture, Natural Resources Conservation Service. National Soil Survey Handbook, Title 430-VI. Available online: http://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/ref/?cid=nrcs142p2_054242 (accessed on 20 October 2022).
- Baloch, F.S.; Karaköy, T.; Demïrbaş, A.; Toklu, F.; Özkan, H.; Hatiïpoğlu, R. Variation of Some Seed Mineral Contents in Open Pollinated Faba Bean (Vicia Faba L.) Landraces from Turkey. Turk. J. Agric. For. 2014, 38, 591–602. [Google Scholar] [CrossRef]
- Rasool, S.; Mahajan, R.; Nazir, M.; Bhat, K.A.; Shikari, A.B.; Ali, G.; Bhat, B.; Bhat, B.A.; Shah, B.; Murtaza, I.; et al. SSR and GBS Based GWAS Study for Identification of QTLs Associated with Nutritional Elemental in Common Bean (Phaseolus Vulgaris L.). Sci. Hortic. 2022, 306, 111470. [Google Scholar] [CrossRef]
- Narwal, R.P.; Kumar, V.; Singh, J.P. Potassium and Magnesium Relationship in Cowpea (Vigna Unguiculata (L.) Walp.). Plant Soil 1985, 86, 129–134. [Google Scholar] [CrossRef]
- Leonard, R.T.; Hotchkiss, C.W. Cation-Stimulated Adenosine Triphosphatase Activity and Cation Transport in Corn Roots. Plant Physiol. 1976, 58, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of Potassium and Magnesium in Photosynthesis, Photosynthate Translocation and Photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [Green Version]
- Koch, M.; Busse, M.; Naumann, M.; Jákli, B. Differential Effects of Varied Potassium and Magnesium Nutrition on Production and Partitioning of Photoassimilates in Potato Plants. Physiologia 2019, 166, 921–935. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Luo, W.; Xu, G. Characterisation of Magnesium Nutrition and Interaction of Magnesium and Potassium in Rice. Ann. Appl. Biol. 2006, 149, 111–123. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, M.L.; Schaefer, R.; Dang, P.; Jiang, T.; Chen, C. GWAS and Coexpression Network Reveal Ionomic Variation in Cultivated Peanut. J. Agric. Food Chem. 2019, 67, 12026–12036. [Google Scholar] [CrossRef]
- Gierth, M.; Mäser, P. Potassium Transporters in Plants–Involvement in K+ Acquisition, Redistribution and Homeostasis. FEBS Lett. 2007, 581, 2348–2356. [Google Scholar] [CrossRef] [Green Version]
- Baoxiang, W.; Yan, L.; Yifeng, W.; Jingfang, L.; Zhiguang, S.; Ming, C.; Yungao, X.; Bo, X.; Bo, Y.; Jian, L.; et al. OsbZIP72 Is Involved in Transcriptional Gene-Regulation Pathway of Abscisic Acid Signal Transduction by Activating Rice High-Affinity Potassium Transporter OsHKT1;1. Rice Sci. 2021, 28, 257–267. [Google Scholar] [CrossRef]
- Vreugdenhil, D.; Aarts, M.G.M.; Koornneef, M.; Nelissen, H.; Ernst, W.H.O. Natural Variation and QTL Analysis for Cationic Mineral Content in Seeds of Arabidopsis Thaliana. Plant Cell Environ. 2004, 27, 828–839. [Google Scholar] [CrossRef]
- Garcia-Oliveira, A.L.; Tan, L.; Fu, Y.; Sun, C. Genetic Identification of Quantitative Trait Loci for Contents of Mineral Nutrients in Rice Grain. J. Integr. Plant Biol. 2009, 51, 84–92. [Google Scholar] [CrossRef]
- Peleg, Z.; Cakmak, I.; Ozturk, L.; Yazici, A.; Jun, Y.; Budak, H.; Korol, A.B.; Fahima, T.; Saranga, Y. Quantitative Trait Loci Conferring Grain Mineral Nutrient Concentrations in Durum Wheat x Wild Emmer Wheat RIL Population. Züchter Genet. Breed. Res. 2009, 119, 353–369. [Google Scholar] [CrossRef] [Green Version]
- Gu, R.; Chen, F.; Liu, B.; Wang, X.; Liu, J.; Li, P.; Pan, Q.; Pace, J.; Soomro, A.-A.; Lübberstedt, T.; et al. Comprehensive Phenotypic Analysis and Quantitative Trait Locus Identification for Grain Mineral Concentration, Content, and Yield in Maize (Zea Mays L.). Theor. Appl. Genet. 2015, 128, 1777–1789. [Google Scholar] [CrossRef] [Green Version]
- Suh, S.J.; Wang, Y.-F.; Frelet, A.; Leonhardt, N.; Klein, M.; Forestier, C.; Mueller-Roeber, B.; Cho, M.H.; Martinoia, E.; Schroeder, J.I. The ATP Binding Cassette Transporter AtMRP5 Modulates Anion and Calcium Channel Activities in Arabidopsis Guard Cells. J. Biol. Chem. 2007, 282, 1916–1924. [Google Scholar] [CrossRef] [Green Version]
- Xing, H.; Fu, X.; Yang, C.; Tang, X.; Guo, L.; Li, C.; Xu, C.; Luo, K. Genome-Wide Investigation of Pentatricopeptide Repeat Gene Family in Poplar and Their Expression Analysis in Response to Biotic and Abiotic Stresses. Sci. Rep. 2018, 8, 2817. [Google Scholar] [CrossRef] [Green Version]
- Saha, D.; Prasad, A.M.; Srinivasan, R. Pentatricopeptide Repeat Proteins and Their Emerging Roles in Plants. Plant Physiol. Biochem. 2007, 45, 521–534. [Google Scholar] [CrossRef]
- Kasajima, I.; Fujiwara, T. Identification of Novel Arabidopsis Thaliana Genes Which Are Induced by High Levels of Boron. Plant Biotechnol. 2007, 24, 355–360. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, K.; Shimizu, A.; Okumoto, Y.; Fujiwara, T.; Matoh, T. Suppression of a NAC-like Transcription Factor Gene Improves Boron-Toxicity Tolerance in Rice. Plant Physiol. 2011, 156, 1457–1463. [Google Scholar] [CrossRef] [Green Version]
- Nyiraguhirwa, S.; Grana, Z.; Ouabbou, H.; Iraqi, D.; Ibriz, M.; Mamidi, S.; Udupa, S.M. A Genome-Wide Association Study Identifying Single-Nucleotide Polymorphisms for Iron and Zinc Biofortification in a Worldwide Barley Collection. Plants 2022, 11, 1349. [Google Scholar] [CrossRef]
- Chen, J.; Vijayakumar, S.; Li, X.; Al-Awqati, Q. Kanadaptin Is a Protein That Interacts with the Kidney but Not the Erythroid Form of Band 3. J. Biol. Chem. 1998, 273, 1038–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, W.A.; Shabala, S.; Cuin, T.A.; Zhou, M.; Penrose, B. Avenues for Biofortification of Zinc in Barley for Human and Animal Health: A Meta-Analysis. Plant Soil 2021, 466, 101–119. [Google Scholar] [CrossRef]
- Hernandez, J.; Meints, B.; Hayes, P. Introgression Breeding in Barley: Perspectives and Case Studies. Front. Plant Sci. 2020, 11, 761. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.; Steffenson, B.J.; Filichkin, T.; Fisk, S.P.; Helgerson, L.; Meints, B.; Vining, K.J.; Marshall, D.; Del Blanco, A.; Chen, X.; et al. Introgression of Rpg4/Rpg5 Into Barley Germplasm Provides Insights Into the Genetics of Resistance to Puccinia Graminis f. Sp. Tritici Race TTKSK and Resources for Developing Resistant Cultivars. Phytopathology 2019, 109, 1018–1028. [Google Scholar] [CrossRef]
- Pickering, R.; Johnston, P.A. Recent Progress in Barley Improvement Using Wild Species of Hordeum. Cytogenet. Genome Res. 2005, 109, 344–349. [Google Scholar] [CrossRef]
- Xu, Q.; Zheng, T.-Q.; Hu, X.; Cheng, L.-R.; Xu, J.-L.; Shi, Y.-M.; Li, Z.-K. Examining Two Sets of Introgression Lines in Rice (Oryza Sativa L.) Reveals Favorable Alleles That Improve Grain Zn and Fe Concentrations. PLoS ONE 2015, 10, e0131846. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.S.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The Genome Sequences of Arachis Duranensis and Arachis Ipaensis, the Diploid Ancestors of Cultivated Peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Jaradat, A.A. Phenotypic and Ionome Profiling of Triticum Aestivum × Aegilops Tauschii Introgression Lines. Crop Sci. 2017, 57, 1916–1934. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Variance Components | Heritability (H2) | |
|---|---|---|---|
| Genotype (Vg) | Residuals (Ve) | ||
| Boron (B) | 0.006 | 0.043 | 0.115 |
| Calcium (Ca) | 0.149 | 0.095 | 0.612 |
| Copper (Cu) | 0.011 | 0.005 | 0.696 |
| Iron (Fe) | 0.022 | 0.023 | 0.481 |
| Potassium (K) | 0.423 | 0.345 | 0.551 |
| Magnesium (Mg) | 0.090 | 0.085 | 0.512 |
| Manganese (Mn) | 0.016 | 0.009 | 0.636 |
| Phosphorus (P) | 0.187 | 0.348 | 0.350 |
| Zinc (Zn) | 0.014 | 0.013 | 0.515 |
| Trait | Year | SNP | Chr | Position | Allele | p-Value | R2 | maf | Stable Gene ID | IPK Descriptions |
|---|---|---|---|---|---|---|---|---|---|---|
| B | 15–16 | S7H_177304153 | 7H | 177304153 | A/G | 2.77 × 10−6 | 0.014 | 0.045 | HORVU.MOREX.r3.7HG0680110 | Potassium transporter |
| HORVU.MOREX.r3.7HG0680120 | 3-oxoacyl-reductase | |||||||||
| HORVU.MOREX.r3.7HG0680130 | NAC (No Apical Meristem) domain transcriptional regulator superfamily protein | |||||||||
| Ca | 15–16 | S1H_514166709 | 1H | 514166709 | C/T | 2.89 × 10−6 | 0.062 | 0.409 | HORVU.MOREX.r3.1HG0094570 | Minichromosome maintenance 8 |
| Ca | 15–16 | S2H_607230798 | 2H | 607230798 | A/C | 1.91 × 10−8 | 0.022 | 0.084 | HORVU.MOREX.r3.2HG0193490 | Growth-regulating factor |
| HORVU.MOREX.r3.2HG0193510 | Gpcr-type g protein 2 | |||||||||
| Ca | 15–16 | S3H_610176767 | 3H | 610176767 | G/T | 1.05 × 10−5 | 0.019 | 0.069 | HORVU.MOREX.r3.3HG0326120 | Glutathione S-transferase T3 |
| HORVU.MOREX.r3.3HG0326130 | FBD-associated F-box protein | |||||||||
| HORVU.MOREX.r3.3HG0326140 | FBD-associated F-box protein | |||||||||
| HORVU.MOREX.r3.3HG0326160 | F-box protein-like protein | |||||||||
| HORVU.MOREX.r3.3HG0326170 | Nuclear transport factor 2 family protein | |||||||||
| HORVU.MOREX.r3.3HG0326190 | ABC transporter ATP-binding protein | |||||||||
| HORVU.MOREX.r3.3HG0326200 | Cofactor assembly | |||||||||
| HORVU.MOREX.r3.3HG0326210 | Mitochondrial-processing peptidase alpha | |||||||||
| HORVU.MOREX.r3.3HG0326220 | Leucine-rich repeat-containing protein 1 | |||||||||
| HORVU.MOREX.r3.3HG0326230 | Vacuolar protein sorting 55 containing protein | |||||||||
| Ca | 15–16 | S4H_591716219 | 4H | 591716219 | C/G | 5.86 × 10−8 | 0.025 | 0.063 | HORVU.MOREX.r3.4HG0411440 | Superoxide dismutase [Cu-Zn] |
| HORVU.MOREX.r3.4HG0411460 | Peptide transporter | |||||||||
| HORVU.MOREX.r3.4HG0411500 | Werner Syndrome-like exonuclease | |||||||||
| Ca | 15–16 | S5H_511093353 | 5H | 511093353 | G/T | 3.59 × 10−8 | 0.003 | 0.233 | HORVU.MOREX.r3.5HG0503550 | Serine/threonine-protein kinase |
| HORVU.MOREX.r3.5HG0503560 | Serine/threonine-protein kinase | |||||||||
| HORVU.MOREX.r3.5HG0503570 | Serine/threonine-protein kinase | |||||||||
| Ca | 15–16 | S5H_541887313 | 5H | 541887313 | C/G | 1.46 × 10−6 | 0.029 | 0.108 | HORVU.MOREX.r3.5HG0516830 | Dehydrin |
| Ca | 15–16 | S6H_42492296 | 6H | 42492296 | C/G | 3.06 × 10−11 | 0.006 | 0.043 | HORVU.MOREX.r3.6HG0555030 | Pentatricopeptide repeat-containing protein |
| HORVU.MOREX.r3.6HG0555040 | Succinate dehydrogenase [ubiquinone] flavoprotein subunit, mitochondrial | |||||||||
| HORVU.MOREX.r3.6HG0555060 | Pentatricopeptide repeat-containing protein | |||||||||
| HORVU.MOREX.r3.6HG0555090 | CASP-like protein | |||||||||
| Ca | 15–16 | S7H_144772413 | 7H | 144772413 | A/G | 3.80 × 10−12 | 0.006 | 0.103 | HORVU.MOREX.r3.7HG0675630 | Pantothenate kinase |
| HORVU.MOREX.r3.7HG0675670 | Chaperone protein dnaJ, putative | |||||||||
| HORVU.MOREX.r3.7HG0675690 | F-box/RNI-like superfamily protein | |||||||||
| Ca | 15–16 | S7H_386796824 | 7H | 386796824 | A/G | 8.56 × 10−7 | 0.097 | 0.119 | HORVU.MOREX.r3.7HG0700320 | Pentatricopeptide repeat-containing protein |
| HORVU.MOREX.r3.7HG0700330 | XH/XS domain protein | |||||||||
| Cu | 06–07 | S1H_21462325 | 1H | 21462325 | A/C | 9.47 × 10−9 | 0.054 | 0.127 | HORVU.MOREX.r3.1HG0009000 | FAR1-related sequence 10 |
| HORVU.MOREX.r3.1HG0009020 | Werner Syndrome-like exonuclease | |||||||||
| HORVU.MOREX.r3.1HG0009050 | MYB transcription factor | |||||||||
| Cu | 06–07 | S2H_10628764 | 2H | 10628764 | C/G | 6.37 × 10−6 | 0.001 | 0.069 | HORVU.MOREX.r3.2HG0099760 | Serpin |
| HORVU.MOREX.r3.2HG0099780 | Maternal effect embryo arrest protein | |||||||||
| HORVU.MOREX.r3.2HG0099790 | Lectin receptor kinase | |||||||||
| HORVU.MOREX.r3.2HG0099800 | Carboxymethylenebutenolidase-like protein | |||||||||
| HORVU.MOREX.r3.2HG0099810 | NAD(P)-binding rossmann-fold protein | |||||||||
| HORVU.MOREX.r3.2HG0099820 | NAD(P)-binding Rossmann-fold protein | |||||||||
| HORVU.MOREX.r3.2HG0099840 | Nicotianamine synthase | |||||||||
| HORVU.MOREX.r3.2HG0099870 | Heat shock protein 90 | |||||||||
| Cu | 06–07 | S3H_11066728 | 3H | 11066728 | C/T | 1.32 × 10−7 | 0.028 | 0.127 | HORVU.MOREX.r3.3HG0224020 | BED Zn finger, hATdimerization domain |
| HORVU.MOREX.r3.3HG0224040 | Pentatricopeptide repeat-containing protein | |||||||||
| Cu | 06–07 | S3H_554863216 | 3H | 554863216 | A/G | 3.96 × 10−6 | 0.035 | 0.297 | HORVU.MOREX.r3.3HG0304030 | Arginine N-methyltransferase (DUF688) |
| HORVU.MOREX.r3.3HG0304070 | Defensin | |||||||||
| HORVU.MOREX.r3.3HG0304080 | Glutaredoxin-like | |||||||||
| Cu | 06–07 | S3H_581535747 | 3H | 581535747 | G/T | 3.38 × 10−6 | 0.008 | 0.162 | HORVU.MOREX.r3.3HG0312640 | AT5g16110 |
| HORVU.MOREX.r3.3HG0312670 | Serine/threonine-protein phosphatase 7 | |||||||||
| HORVU.MOREX.r3.3HG0312690 | Potassium transporter | |||||||||
| Cu | 06–07 | S5H_560889577 | 5H | 560889577 | A/G | 8.41 × 10−9 | 0.013 | 0.069 | HORVU.MOREX.r3.5HG0525290 | Amino acid transporter, putative |
| Cu | 06–07 | S7H_123030780 | 7H | 123030780 | C/T | 1.58 × 10−13 | 0.022 | 0.166 | HORVU.MOREX.r3.7HG0671730 | Kanadaptin |
| HORVU.MOREX.r3.7HG0671750 | RNA-directed DNA polymerase | |||||||||
| Fe | 06–07 | S1H_454616791 | 1H | 454616791 | C/G | 7.16 × 10−6 | 0.009 | 0.101 | HORVU.MOREX.r3.1HG0071240 | Polynucleotidyl transferase, RnaseH family |
| HORVU.MOREX.r3.1HG0071260 | Receptor-like kinase | |||||||||
| Fe | 06–07 | S3H_31618512 | 3H | 31618512 | A/G | 2.97 × 10−6 | 0.026 | 0.153 | HORVU.MOREX.r3.3HG0233410 | Transmembrane protein, putative |
| HORVU.MOREX.r3.3HG0233420 | LINE−1 reverse transcriptase like | |||||||||
| HORVU.MOREX.r3.3HG0233430 | Polynucleotidyl transferase, RNaseH family | |||||||||
| Fe | 06–07 | S3H_557143396 | 3H | 557143396 | C/G | 5.87 × 10−6 | 0.011 | 0.121 | HORVU.MOREX.r3.3HG0304800 | SMC protein |
| HORVU.MOREX.r3.3HG0304810 | Ethylene-responsive transcription factor | |||||||||
| HORVU.MOREX.r3.3HG0304830 | Protein kinase | |||||||||
| Fe | 06–07 | S5H_356777800 | 5H | 356777800 | C/T | 4.75 × 10−6 | 0.031 | 0.252 | None | N/A |
| Fe | 06–07 | S5H_526313021 | 5H | 526313021 | C/G | 7.16 × 10−7 | 0.000 | 0.453 | HORVU.MOREX.r3.5HG0510200 | Protein OBERON 1 |
| HORVU.MOREX.r3.5HG0510210 | Tryptophan RNA-binding attenuator protein | |||||||||
| HORVU.MOREX.r3.5HG0510220 | Late embryogenesis abundant protein | |||||||||
| HORVU.MOREX.r3.5HG0510230 | Dirigent protein | |||||||||
| HORVU.MOREX.r3.5HG0510240 | Dirigent protein | |||||||||
| HORVU.MOREX.r3.5HG0510250 | Dirigent protein | |||||||||
| Fe | 06–07 | S7H_623488700 | 7H | 623488700 | G/T | 3.78 × 10−7 | 0.022 | 0.496 | HORVU.MOREX.r3.7HG0749550 | Disease resistance protein RPM1 |
| HORVU.MOREX.r3.7HG0749560 | Coiled-coil domain-containing protein 12 | |||||||||
| HORVU.MOREX.r3.7HG0749570 | Prolyl oligopeptidase family protein | |||||||||
| Fe | 06–07 | S7H_630244607 | 7H | 630244607 | A/G | 1.10 × 10−10 | 0.033 | 0.332 | HORVU.MOREX.r3.7HG0752330 | 2-oxoglutarate Fe(II)-dependent oxygenase |
| HORVU.MOREX.r3.7HG0752340 | 2-oxoglutarate Fe(II)-dependent oxygenase | |||||||||
| HORVU.MOREX.r3.7HG0752350 | AT hook motif DNA-binding family protein | |||||||||
| K | 15–16 | S3H_19711917 | 3H | 19711917 | A/G | 2.60 × 10−6 | 0.003 | 0.131 | HORVU.MOREX.r3.3HG0228430 | transmembrane protein, putative (DUF247) |
| HORVU.MOREX.r3.3HG0228450 | ArfGap/RecO-like Zn finger domain protein | |||||||||
| K | 15–16 | S3H_609313148 | 3H | 609313148 | C/T | 1.91 × 10−6 | 0.008 | 0.162 | HORVU.MOREX.r3.3HG0325670 | F-box domain containing protein |
| HORVU.MOREX.r3.3HG0325680 | Histidine decarboxylase | |||||||||
| HORVU.MOREX.r3.3HG0325700 | KH domain-containing protein | |||||||||
| HORVU.MOREX.r3.3HG0325710 | Serine/threonine-protein phosphatase 7 | |||||||||
| HORVU.MOREX.r3.3HG0325720 | Actin cytoskeleton-regulatory complex pan1 | |||||||||
| HORVU.MOREX.r3.3HG0325730 | Gamma-tubulin complex component | |||||||||
| K | 15–16 | S5H_496079876 | 5H | 496079876 | C/T | 4.11 × 10−7 | 0.024 | 0.065 | None | N/A |
| K | 15–16 | S6H_403845268 | 6H | 403845268 | A/C | 1.18 × 10−5 | 0.001 | 0.043 | None | N/A |
| K | 15–16 | S7H_221025298 | 7H | 221025298 | A/G | 9.44 × 10−6 | 0.020 | 0.123 | None | N/A |
| K | 15–16 | S7H_488772960 | 7H | 488772960 | C/T | 6.52 × 10−6 | 0.009 | 0.261 | HORVU.MOREX.r3.7HG0713790 | Cellulose synthase |
| HORVU.MOREX.r3.7HG0713820 | Transcription factor | |||||||||
| Mn | 06–07 | S1H_437852428 | 1H | 437852428 | C/G | 4.95 × 10−6 | 0.009 | 0.037 | HORVU.MOREX.r3.1HG0067260 | NB-ARC domain-disease resistance protein |
| HORVU.MOREX.r3.1HG0067270 | H/ACA ribonucleoprotein complex NAF1 | |||||||||
| Mn | 15–16 | S6H_7595410 | 6H | 7595410 | A/G | 5.01 × 10−6 | 0.000 | 0.041 | HORVU.MOREX.r3.6HG0541060 | Ubiquitin family protein |
| HORVU.MOREX.r3.6HG0541070 | Outer envelope protein 61 | |||||||||
| HORVU.MOREX.r3.6HG0541100 | Polygalacturonase non-catalytic protein | |||||||||
| HORVU.MOREX.r3.6HG0541110 | F-box domain containing protein | |||||||||
| Zn | 15–16 | S5H_525224694 | 5H | 525224694 | A/C | 1.00 × 10−5 | 0.003 | 0.209 | HORVU.MOREX.r3.5HG0509880 | Kinase |
| HORVU.MOREX.r3.5HG0509890 | General transcription factor 3C polypeptide 3 | |||||||||
| HORVU.MOREX.r3.5HG0509900 | Stem-specific protein TSJT1 | |||||||||
| Fe | Both | S5H_76074047 | 5H | 76074047 | C/T | >3.1 × 10−4 | 0.008 | 0.50000 | HORVU.MOREX.r3.5HG0439860 | Disease resistance protein RPM1 |
| HORVU.MOREX.r3.5HG0439870 | Dicer-like 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abendroth, J.A.; Sallam, A.H.; Steffenson, B.J.; Vinje, M.A.; Mahalingam, R.; Walling, J.G. Identification of Genomic Loci Controlling Grain Macro and Micronutrient Variation in a Wild Barley (Hordeum vulgare spp. spontaneum) Diversity Panel. Agronomy 2022, 12, 2839. https://doi.org/10.3390/agronomy12112839
Abendroth JA, Sallam AH, Steffenson BJ, Vinje MA, Mahalingam R, Walling JG. Identification of Genomic Loci Controlling Grain Macro and Micronutrient Variation in a Wild Barley (Hordeum vulgare spp. spontaneum) Diversity Panel. Agronomy. 2022; 12(11):2839. https://doi.org/10.3390/agronomy12112839
Chicago/Turabian StyleAbendroth, Jillian A., Ahmad H. Sallam, Brian J. Steffenson, Marcus A. Vinje, Ramamurthy Mahalingam, and Jason G. Walling. 2022. "Identification of Genomic Loci Controlling Grain Macro and Micronutrient Variation in a Wild Barley (Hordeum vulgare spp. spontaneum) Diversity Panel" Agronomy 12, no. 11: 2839. https://doi.org/10.3390/agronomy12112839