
Diversity and Resistance to Thiophanate-Methyl of Colletotrichum spp. in Strawberry Nursery and the Development of Rapid Detection Using LAMP Method


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Colletotrichum Isolates
2.2. Genomic DNA Extraction, PCR Amplification and Sequencing
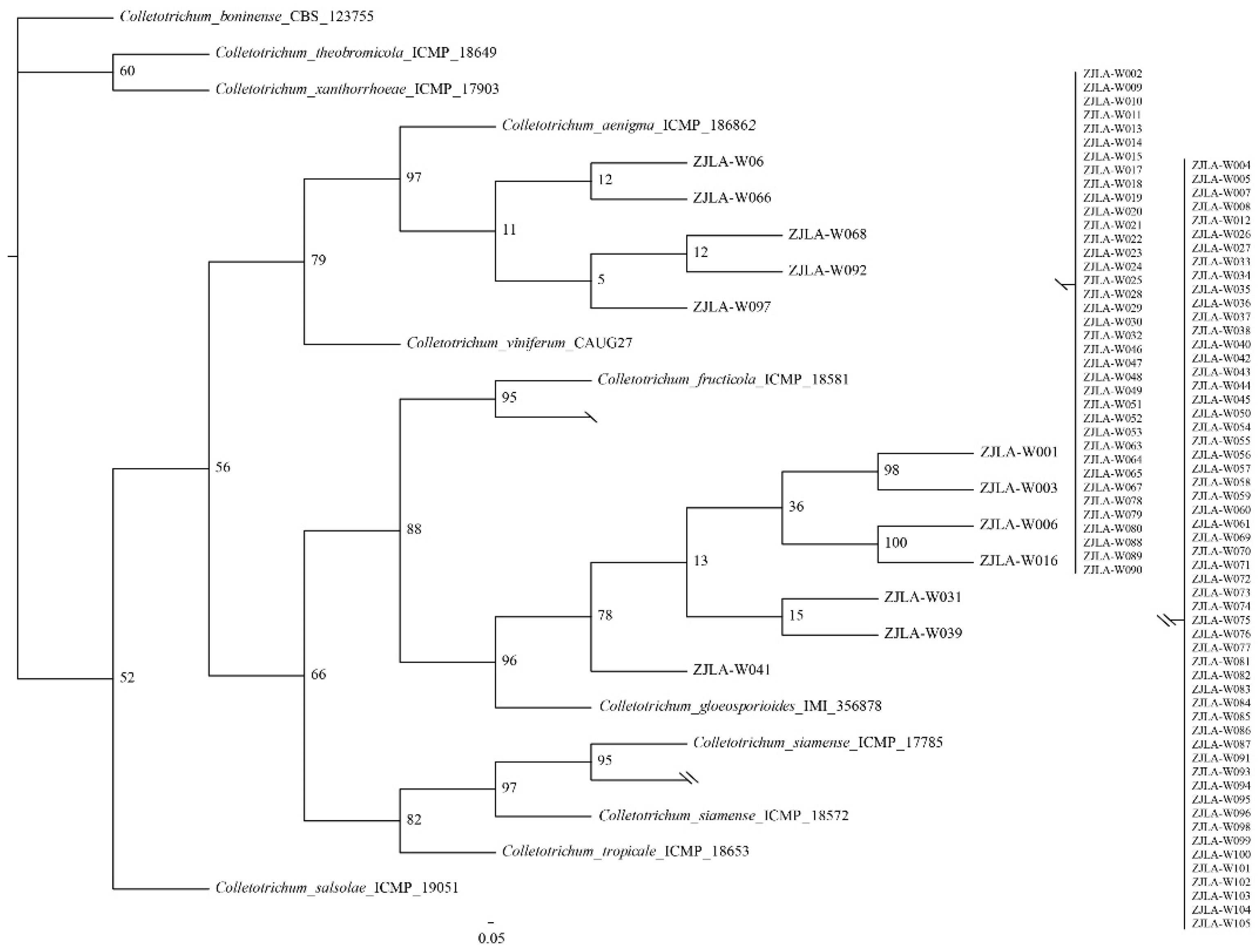
2.3. Multilocus Phylogenetic Analysis
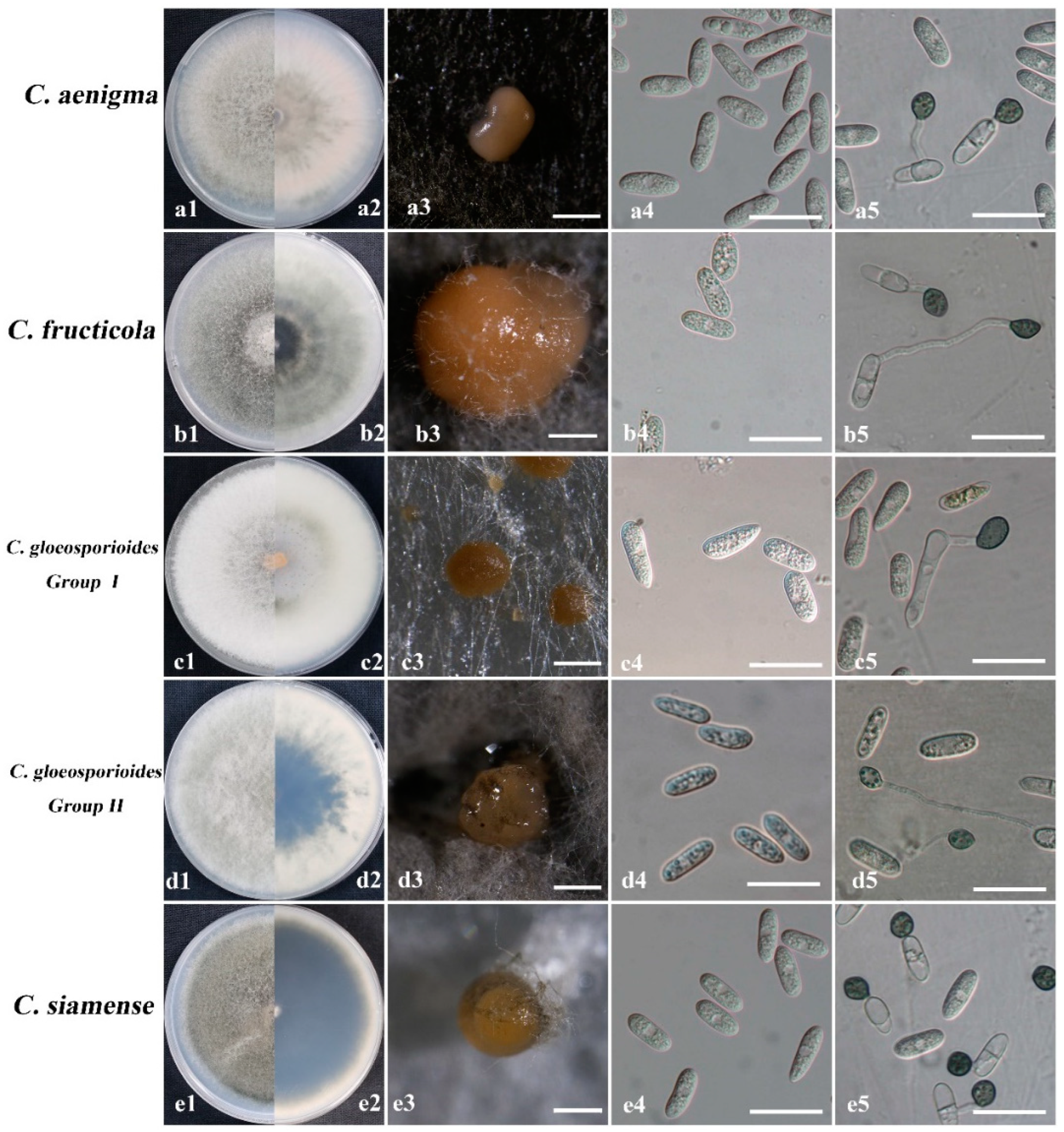
2.4. Morphological Characterization
2.5. Characterization of Resistance of Colletotrichum spp. to Benzimidazole Fungicides
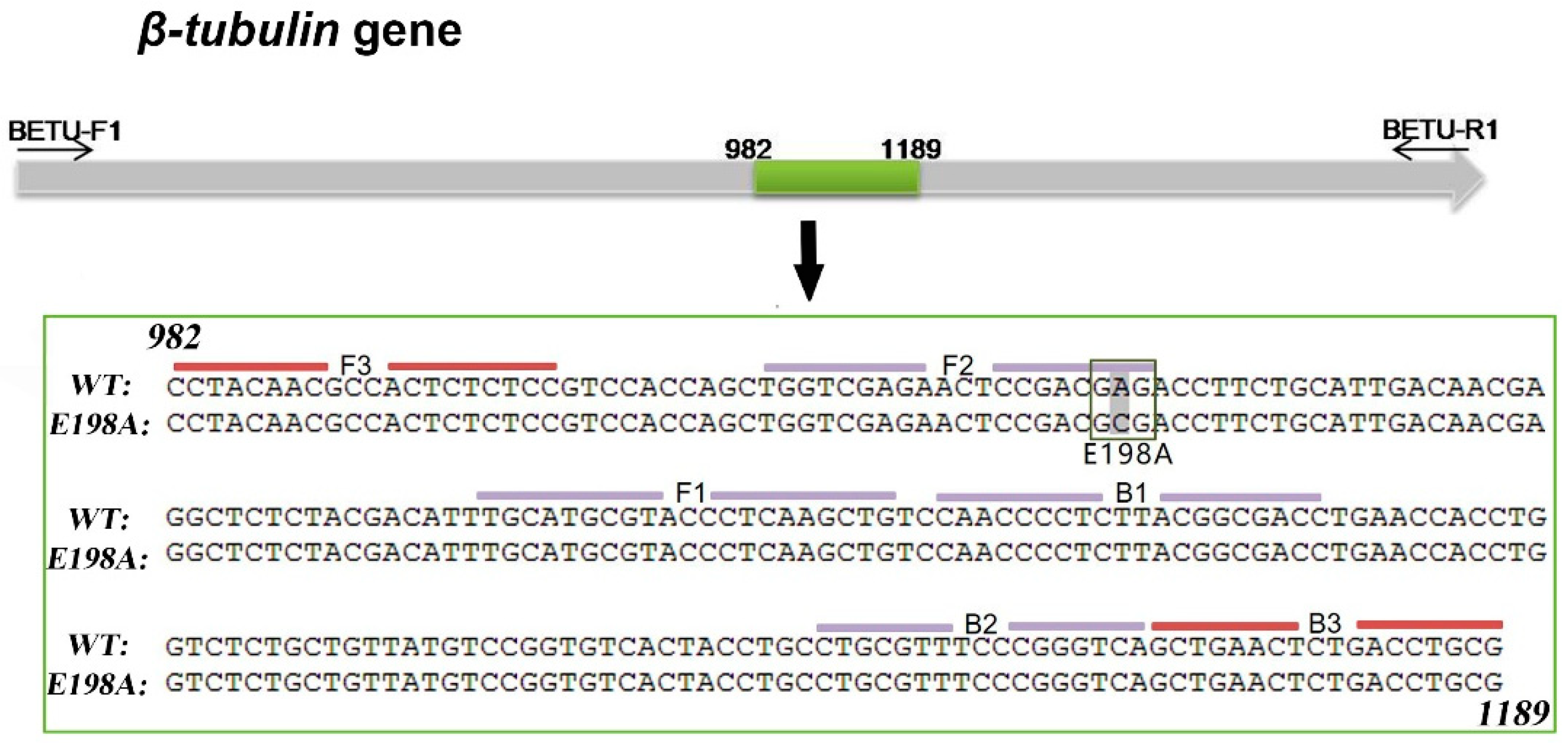
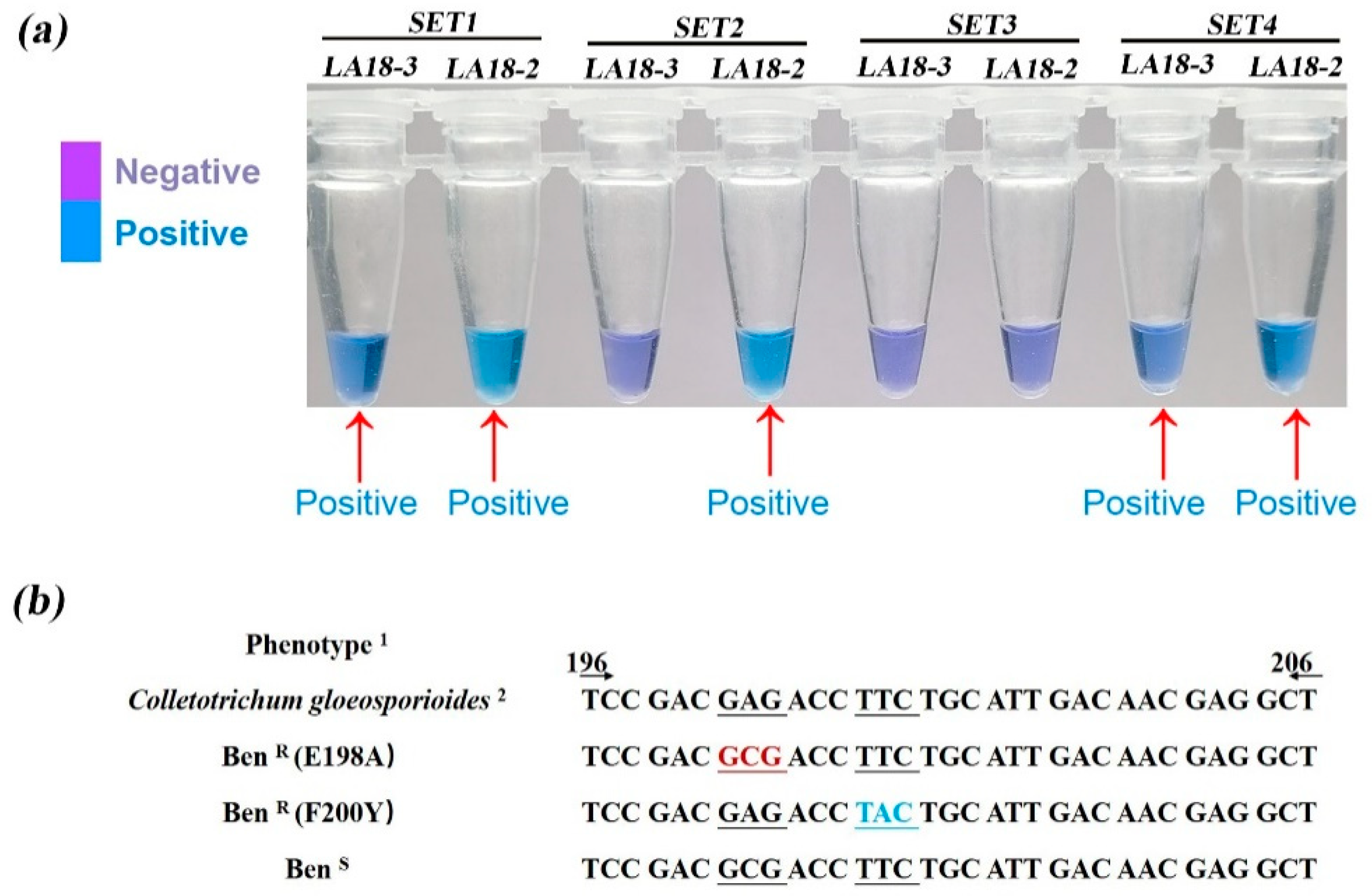
2.6. LAMP Primer Design and Optimization for Detection of the E198A Mutants
2.7. LAMP Assay Used for Detection of E198A-Resistance Genotype
2.8. Analysis of β-Tubulin Gene Sequences of Colletotrichum Isolates
3. Results
3.1. Isolation and Identification of Colletotrichum Species
3.2. Morphology Description
3.3. Thiophanate-Methyl Resistance of Colletotrichum spp. Isolates Are a Severe Problem in Strawberry Nurseries
3.4. Point Mutations in Codon 198 and 200 of the β-Tubulin Gene Are Responsible for High Resistance to MBC Fungicides in Strawberry Anthracnose at the Seedling Stage
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Miller, K.; Feucht, W.; Schmid, M. Bioactive Compounds of Strawberry and Blueberry and Their Potential Health Effects Based on Human Intervention Studies: A Brief Overview. Nutrients 2019, 11, 1510. [Google Scholar] [CrossRef] [Green Version]
- Giampieri, F.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Alvarez-Suarez, J.M.; Afrin, S.; Bompadre, S.; Quiles, J.; Mezzeti, B.; Battino, M. Strawberry as a Health Promoter: An Evidence Based Review. Food Funct. 2015, 6, 1386–1398. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Cao, T.; Huang, Y.; Gao, J.; Hong, W. Investigation On the Occurrence Rules of Strawberry Anthracnose, Grey Mould and Powdery Mildew. Zhejiang Agric. Sci. 2018, 59, 2177–2178. [Google Scholar]
- Howard, C.M.; Maas, J.L.; Chandler, C.L.; Albregts, E.A. Anthracnose of Strawberry Caused by the Colletotrichum complex in Florida. Plant Dis. 1992, 76, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.; Miao, J.; Liu, X.; Zhang, G. Characterization of Colletotrichum spp. Sensitivity to Carbendazim for Isolates Causing Strawberry Anthracnose in China. Plant Dis. 2021, 105, 87–95. [Google Scholar] [CrossRef]
- Jian, Y.; Li, Y.; Tang, G.; Zheng, X.; Khaskheli, M.I.; Gong, G. Identification of Colletotrichum Species Associated with Anthracnose Disease of Strawberry in Sichuan Province, China. Plant Dis. 2021, 105, 3025–3036. [Google Scholar] [CrossRef]
- Han, Y.C.; Zeng, X.G.; Xiang, F.Y.; Ren, L.; Chen, F.Y.; Gu, Y.C. Distribution and Characteristics of Colletotrichum spp. Associated with Anthracnose of Strawberry in Hubei, China. Plant Dis. 2016, 100, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Xu, X.F.; Dai, D.J.; Shi, H.J.; Wang, H.D.; Zhang, C.Q. Differentiation in Development of Benzimidazole Resistance In Colletotrichum gloeosporioides Complex Populations From Strawberry and Grape Hosts. Australas. Plant Pathol. 2016, 45, 241–249. [Google Scholar] [CrossRef]
- Chen, X.Y.; Dai, D.J.; Zhao, S.F.; Shen, Y.; Wang, H.D.; Zhang, C.Q. Genetic Diversity of Colletotrichum spp. Causing Strawberry Anthracnose in Zhejiang, China. Plant Dis. 2020, 104, 1351–1357. [Google Scholar] [CrossRef]
- Weir, B.S.; Johnston, P.R.; Damm, U. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 2012, 73, 115–180. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Ojiambo, P.; Louws, F. Initial Inoculum and Spatial Dispersal of Colletotrichum gloeosporioides, the Causal Agent of Strawberry Anthracnose Crown Rot. Plant Dis. 2015, 99, 80–86. [Google Scholar] [CrossRef]
- Forcelini, B.B.; Seijo, T.E.; Amiri, A.; Peres, N.A. Resistance in Strawberry Isolates of Colletotrichum acutatum from Florida to Quinone-Outside Inhibitor Fungicides. Plant Dis. 2016, 100, 2050–2056. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Zhang, Y.; Yu, H.; Zhou, J.; Hu, M.; Liu, A.; Wu, J.; Wang, H.; Zhang, C. Colletotrichum Spp. Diversity Between Leaf Anthracnose and Crown Rot From the Same Strawberry Plant. Front. Microbiol. 2022, 13, 860694. [Google Scholar] [CrossRef]
- Luo, Q.; Schoeneberg, A.; Hu, M. Resistance to Azoxystrobin and Thiophanate-Methyl Is Widespread in Colletotrichum spp. Isolates From the Mid-Atlantic Strawberry Fields. Plant Dis. 2021, 105, 2202–2208. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.J.; Black, L.L. Morphological, Cultural, and Pathogenic Variation among Colletotrichum species Isolated From strawberry. Plant Dis. 1990, 74, 69–76. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Huang, J.K.; Jin, B.C.; Yan, J.Y.; Li, X.H.; Hyde, K.D.; Bahkali, A.H.; Yin, S.L.; Zhang, G.Z. An Account of Colletotrichum species Associated with Strawberry Anthracnose in China Based on Morphology and Molecular data. Mycosphere 2016, 7, 1147–1163. [Google Scholar] [CrossRef]
- Furuta, K.; Nagashima, S.; Inukai, T.; Masuta, C. Construction of a System for the Strawberry Nursery Production towards Elimination of Latent Infection of Anthracnose Fungi by a Combination of PCR and Microtube Hybridization. Plant Pathol. J. 2017, 33, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Song, L.; Xu, X.; Zou, X.; Duan, K.; Gao, Q. Characterization and Fungicide Sensitivity of Colletotrichum Species Causing Strawberry Anthracnose in Eastern China. Plant Dis. 2020, 104, 1960–1968. [Google Scholar] [CrossRef]
- Torres-Calzada, C.; Tapia-Tussell, R.; Higuera-Ciapara, I.; Martin-Mex, R.; Nexticapan-Garcez, A.; Perez-Brito, D. Sensitivity of Colletotrichum truncatum to Four Fungicides and Characterization of Thiabendazole-Resistant Isolates. Plant Dis. 2015, 99, 1590–1595. [Google Scholar] [CrossRef] [Green Version]
- Vielba-Fernandez, A.; Vicente, A.; Perez-Garcia, A.; Fernandez-Ortuno, D. Monitoring Methyl Benzimidazole Carbamate-Resistant Isolates of the Cucurbit Powdery Mildew Pathogen, Podosphaera xanthii, Using Loop-Mediated Isothermal Amplification. Plant Dis. 2019, 103, 1515–1524. [Google Scholar] [CrossRef]
- Duan, Y.B.; Yang, Y.; Wang, J.X.; Chen, C.J.; Steinberg, G.; Fraaije, B.A.; Zhou, M.G. Simultaneous Detection of Multiple Benzimidazole-Resistant beta-Tubulin Variants of Botrytis cinerea using Loop-Mediated Isothermal Amplification. Plant Dis. 2018, 102, 2016–2024. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.H.; Chung, W.C.; Peng, M.T.; Yang, H.R.; Huang, J.W. Specific Detection of Benzimidazole Resistance in Colletotrichum gloeosporioides From Fruit Crops by PCR-RFLP. New Biotechnol. 2010, 27, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yoshimura, M.A.; Michailides, T.J. Identification and Characterization of Benzimidazole Resistance in Monilinia fructicola From Stone Fruit Orchards in California. Appl. Environ. Microbiol. 2003, 69, 7145–7152. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.B.; Yang, Y.; Wang, J.X.; Liu, C.C.; He, L.L.; Zhou, M.G. Development and Application of Loop-mediated isothermal amplification for Detecting theHighly Benzimidazole-resistant Isolates in Sclerotinia sclerotiorum. Sci. Rep. 2015, 5, 17278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, H.E.; Cools, H.J.; West, J.S.; Shaw, M.W.; Fraaije, B.A. Detection and Molecular Characterisation of Pyrenopeziza brassicae Isolates Resistant to Methyl Benzimidazole Carbamates. Pest Manag. Sci. 2013, 69, 1040–1048. [Google Scholar] [CrossRef]
- Han, Y.C.; Zeng, X.G.; Xiang, F.Y.; Zhang, Q.H.; Guo, C.; Chen, F.Y.; Chen, G.Y. Carbendazim Sensitivity in Populations of Colletotrichum gloeosporioides complex Infecting Strawberry and Yams in Hubei Province of China. J. Integr. Agric. 2018, 17, 1391–1400. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.B.; Yang, Y.; Li, T.; Zhao, D.; Cao, J.; Shi, Y.Y.; Wang, J.X.; Zhou, M.G. Development of A Rapid and High-throughput Molecular Method for Detecting the F200Y Mutant Genotype in Benzimidazole-resistant Isolates of Fusarium asiaticum. Pest Manag. Sci. 2016, 72, 2128–2135. [Google Scholar] [CrossRef]
- Wu, J.Y.; Hu, X.R.; Zhang, C.Q. Molecular Detection of QoI Resistance in Colletotrichum gloeosporioides Causing Strawberry Anthracnose Based on Loop-Mediated Isothermal Amplification Assay. Plant Dis. 2019, 103, 1319–1325. [Google Scholar] [CrossRef]
- Xie, L.; Zhang, J.Z.; Wan, Y.; Hu, D.W. Identification of Colletotrichum spp. Isolated From Strawberry in Zhejiang Province and Shanghai City, China. J. Zhejiang Univ. Sci. B 2010, 11, 61–70. [Google Scholar] [CrossRef]
- Gunnell, P.S.; Gubler, W.D. Taxonomy and Morphology of Colletotrichum Species Pathogenic to Strawberry. Mycologia 1992, 84, 157–165. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Templeton, M.D.; Rikkerink, E.H.; Solon, S.L.; Crowhurst, R.N. Cloning and Molecular Characterization of the Glyceraldehyde-3-phosphate dehydrogenase-encoding gene and cDNA From the Plant Pathogenic Fungus Glomerella cingulata. Gene 1992, 122, 225–230. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation Software for the Fast Assembly of Multi-Gene Datasets with Character Set and Codon Information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Page, R.D. TreeView: An Application to Display Phylogenetic Trees on Personal Computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Fu, M.; Crous, P.W.; Bai, Q.; Zhang, P.F.; Xiang, J.; Guo, Y.S.; Zhao, F.F.; Yang, M.M.; Hong, N.; Xu, W.X.; et al. Colletotrichum species Associated with Anthracnose of Pyrus spp. in China. Persoonia 2019, 42, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Yuan, S.K.; Hu, X.R.; Zhang, C.Q. Shift of Sensitivity in Botrytis cinerea to Benzimidazole Fungicides in Strawberry Greenhouse Ascribing to the Rising-lowering of E198A Subpopulation and its Visual, On-site Monitoring by Loop-mediated Isothermal Amplification. Sci. Rep. 2019, 9, 11644. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van-Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.D.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, J.E.; Foster, G.D. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Jayawardena, R.; Goonasekara, I.; Wang, Y.; Zhang, W.; Liu, M.; Huang, J.B.; Wang, Z.Y.; Shang, J.J.; Peng, Y.L.; et al. Diverse Species of Colletotrichum associated with Grapevine Anthracnose in China. Fungal Divers. 2014, 71, 233–246. [Google Scholar] [CrossRef]
- He, L.; Li, X.; Gao, Y.; Li, B.; Mu, W.; Liu, F. Characterization and Fungicide Sensitivity of Colletotrichum spp. from Different Hosts in Shandong, China. Plant Dis. 2019, 103, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Curry, K.J.; Abril, M.; Avant, J.B.; Smith, B.J. Strawberry Anthracnose: Histopathology of Colletotrichum acutatum and C. fragariae. Phytopathology 2002, 92, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller-Butler, M.A.; Smith, B.J.; Babiker, E.M.; Kreiser, B.R.; Blythe, E.K. Comparison of Whole Plant and Detached Leaf Screening Techniques for Identifying Anthracnose Resistance in Strawberry Plants. Plant Dis. 2018, 102, 2112–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Harrington, T.C.; Batzer, J.C.; Kubota, R.; Peres, N.A.; Gleason, M.L. Detection of Colletotrichum acutatum Sensu Lato on Strawberry by Loop-Mediated Isothermal Amplification. Plant Dis. 2016, 100, 1804–1812. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Valladares, M.; Valderas-Garcia, E.; Gandasegui, J.; Skuce, P.; Morrison, A.; Aguero, V.C.G.; Cambra-Pelleja, M.; Balana-Fouce, R.; Roko-Vazquez, F.A. Teladorsagia circumcincta Beta Tubulin: The Presence of the E198L Polymorphism on Its Own is Associated with Benzimidazole Resistance. Parasites Vectors 2020, 13, 453–455. [Google Scholar] [CrossRef]
- Qin, Y.; Duan, X.; Peng, Y.; Rui, Y. Rapid Detection of a Novel B1-beta-lactamase Gene, blaAFM-1 Using a Loop-mediated Isothermal Amplification (LAMP) assay. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 80–87. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Set | Type | Sequence (5′→3′) |
|---|---|---|---|
| F3 | - | Forward | CCTACAACGCCACTCTCTCC |
| B3 | - | Backward | CGCAGGTCAGAGTTCAGC |
| FIP1 | SET1 | Forward outer | ACAGCTTGAGGGTACGCATGCA-TGGTCGAGAACTCCGACGCG |
| FIP2 | SET2 | Backward outer | ACAGCTTGAGGGTACGCATGCA-TGGTCGAGAACTCCGACCCC |
| FIP3 | SET3 | Forward inner | ACAGCTTGAGGGTACGCATGCA-TGGTCGAGAACTCCGACTCG |
| FIP4 | SET4 | Backward inner | ACAGCTTGAGGGTACGCATGCA-TGGTCGAGAACTCCGACACG |
| BIP | - | Forward outer | CAACCCCTCTTACGGCGACCT-TGACCCGGGAAACGCAG |
| BETU-F1 | Forward | CAGGTTCTCGATGTTGTCCG | |
| BETU-R1 | Backward | CCATCATGTTCTTGGGGTCG |
| Species 1 /Isolate No. | Colony Morphology 2 | Growth Rate (mm/d) 3 | Conidiomata 4 | Conidia 5 | Appressoria 5 | ||
|---|---|---|---|---|---|---|---|
| Length (μm) | Width (μm) | Length (μm) | Width (μm) | ||||
| C. aenigma /ZJLA-W062 | R, C, GG(c) to LG(m) | 10.7 | O, S | 14.77 ± 0.35 (11.24–21.29) | 5.65 ± 0.13 (3.28–7.26) | 6.97 ± 0.51 (6.29–8.15) | 4.90 ± 0.16 (4.60–5.22) |
| C. fructicola /ZJLA-W009 | R, C, O | 10.8 | O, S | 13.65 ± 0.20 (10.63–17.27) | 5.61 ± 0.10 (3.54–7.34) | 7.13 ± 0.37 (4.96–8.50) | 5.28 ± 0.13 (4.78–5.67) |
| C. gloeosporioides /ZJLA-W016 | R, C, W | 13.8 | O, U | 13.91 ± 0.25 (10.38–18.24) | 4.84 ± 0.09 (3.54–6.64) | 6.61 ± 0.72 (5.40–7.79) | 5.67 ± 0.19 (4.51–7.08) |
| C. gloeosporioides /ZJLA-W039 | R, C, GG | 13.2 | O, S | 13.63 ± 0.32 (9.71–16.29) | 4.82 ± 0.12 (3.63–6.55) | 6.57 ± 0.37 (5.60–8.23) | 5.25 ± 0.17 (4.31–6.19) |
| C. siamense /ZJLA-W005 | R, C, A | 10.6 | O, S | 12.68 ± 0.21 (9.83–15.02) | 4.89 ± 0.09 (3.81–6.22) | 6.65 ± 0.12 (5.49–8.32) | 5.62 ± 0.11 (4.51–6.96) |
| Species 1 | N 2 | Resistance Level by MIC 3 | Positive in LAMP 4 | Amino Acid Substitution 5 | ||||
|---|---|---|---|---|---|---|---|---|
| S | LR | MR | HR | E198A | F200Y | |||
| C. aenigma | 5 | 0 | 0 | 1 | 4 | 4 | 4 | 1 |
| C. fructicola | 37 | 2 | 0 | 0 | 35 | 33 | 33 | 2 |
| C. gloeosporioides | 7 | 1 | 0 | 0 | 6 | 5 | 5 | 1 |
| C. siamense | 56 | 1 | 0 | 1 | 54 | 53 | 53 | 2 |
| Total | 105 | 4 | 0 | 2 | 99 | 95 | 95 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Hu, S.; Ye, B.; Hu, X.; Xiao, W.; Yu, H.; Zhang, C. Diversity and Resistance to Thiophanate-Methyl of Colletotrichum spp. in Strawberry Nursery and the Development of Rapid Detection Using LAMP Method. Agronomy 2022, 12, 2815. https://doi.org/10.3390/agronomy12112815
Wu J, Hu S, Ye B, Hu X, Xiao W, Yu H, Zhang C. Diversity and Resistance to Thiophanate-Methyl of Colletotrichum spp. in Strawberry Nursery and the Development of Rapid Detection Using LAMP Method. Agronomy. 2022; 12(11):2815. https://doi.org/10.3390/agronomy12112815
Chicago/Turabian StyleWu, Jianyan, Shuodan Hu, Boyang Ye, Xiaoran Hu, Wenfei Xiao, Hong Yu, and Chuanqing Zhang. 2022. "Diversity and Resistance to Thiophanate-Methyl of Colletotrichum spp. in Strawberry Nursery and the Development of Rapid Detection Using LAMP Method" Agronomy 12, no. 11: 2815. https://doi.org/10.3390/agronomy12112815