

Physiological Responses of Plants to Combined Drought and Heat under Elevated CO2


Abstract
:1. Introduction
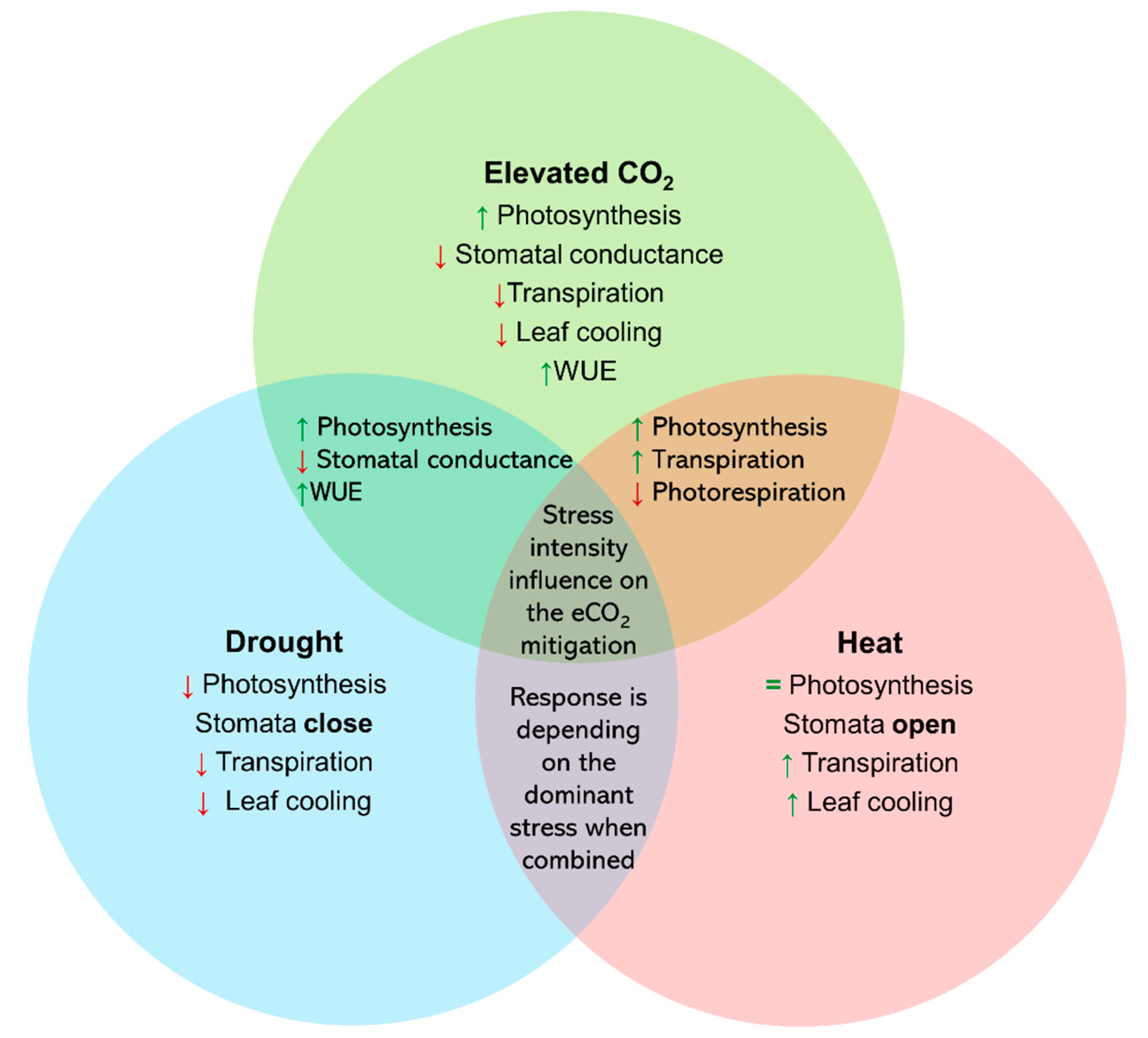
2. Elevated CO2 Improves Plant Photosynthesis and WUE under Moderate Drought Stress
3. Elevated CO2 Reduces Plant Photorespiration and Enhances Photosynthesis under Heat Stress
4. Complex Physiological Responses of Plants to Drought and Heat Stress under eCO2
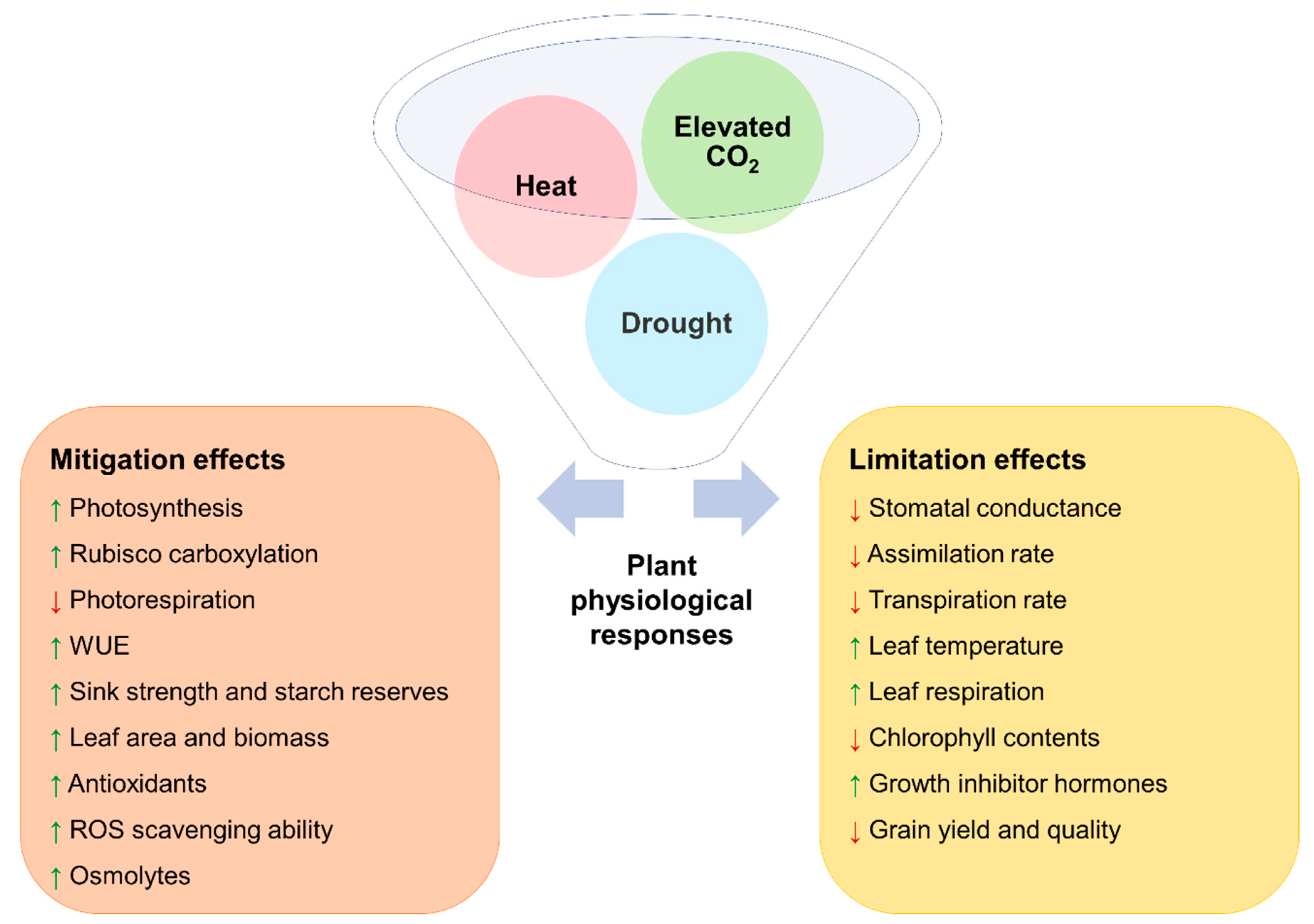
5. The Mitigation of eCO2 on Plants under Combined Drought and Heat Stress
6. The Limitations of eCO2 on Plants under Combined Drought and Heat Stress
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- IPCC. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. 2021. Available online: https://www.ipcc.ch/report/ar6/wg1/ (accessed on 10 July 2021).
- NOAA. Trends in Atmospheric Carbon Dioxide. 2022. Available online: https://gml.noaa.gov/ccgg/trends/gl_trend.html (accessed on 23 August 2022).
- IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. 2007, Volume 59, p. 996. Available online: https://www.ipcc.ch/report/ar4/wg1/ (accessed on 8 June 2021).
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. 2014, Volume 151. Available online: https://www.ipcc.ch/report/ar5/syr/ (accessed on 11 August 2018).
- Hammer, G.L.; McLean, G.; van Oosterom, E.; Chapman, S.; Zheng, B.; Wu, A.; Doherty, A.; Jordan, D. Designing crops for adaptation to the drought and high-temperature risks anticipated in future climates. Crop Sci. 2020, 60, 605–621. [Google Scholar] [CrossRef]
- Wang, B.; Liu, D.L.; Asseng, S.; Macadam, I.; Yu, Q. Modelling wheat yield change under CO2 increase, heat and water stress in relation to plant available water capacity in eastern Australia. Eur. J. Agron. 2017, 90, 152–161. [Google Scholar] [CrossRef]
- Dietzen, C.A.; Larsen, K.S.; Ambus, P.L.; Michelsen, A.; Arndal, M.F.; Beier, C.; Reinsch, S.; Schmidt, I.K. Accumulation of soil carbon under elevated CO2 unaffected by warming and drought. Glob. Chang. Biol. 2019, 25, 2970–2977. [Google Scholar] [CrossRef]
- Wang, J.; Vanga, S.K.; Saxena, R.; Orsat, V.; Raghavan, V. Effect of climate change on the yield of cereal crops: A review. Climate 2018, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Chem. 2018, 6, 26. [Google Scholar] [CrossRef]
- Muluneh, M.G. Impact of climate change on biodiversity and food security: A global perspective—A review article. Agric. Food Secur. 2021, 10, 1–25. [Google Scholar] [CrossRef]
- Cooper, M.; Tang, T.; Gho, C.; Hart, T.; Hammer, G.; Messina, C. Integrating genetic gain and gap analysis to predict improvements in crop productivity. Crop Sci. 2020, 60, 582–604. [Google Scholar] [CrossRef] [Green Version]
- Pareek, A.; Dhankher, O.P.; Foyer, C.H. Mitigating the impact of climate change on plant productivity and ecosystem sustainability. J. Exp. Bot. 2020, 71, 451–456. [Google Scholar] [CrossRef]
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and unique responses of plants to multiple individual stresses and stress combinations: Physiological and molecular mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef] [Green Version]
- Hatfield, J.L.; Dold, C. Agroclimatology and wheat production: Coping with climate change. Front. Plant Sci. 2018, 9, 1–5. [Google Scholar] [CrossRef]
- Gaspar, T.; Franck, T.; Bisbis, B.; Kevers, C.; Jouve, L.; Hausman, J.F.; Dommes, J. Concepts in plant stress physiology. Application to plant tissue cultures. Plant Growth Regul. 2002, 37, 263–285. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Manuka, R.; Penna, S. Plant sugars: Homeostasis and transport under abiotic stress in plants. Physiol. Plant 2021, 171, 739–755. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Devi, P.; Chaudhary, S.; Rani, A.; Jha, U.C.; Kumar, S.; Bindumadhava, H.; Prasad, P.V.V.; Sharma, K.D.; Siddique, K.H.M.; et al. ‘Omics’ approaches in developing combined drought and heat tolerance in food crops. Plant Cell Rep. 2022, 41, 699–739. [Google Scholar] [CrossRef]
- Kadam, N.N.; Xiao, G.; Melgar, R.J.; Bahuguna, R.N.; Quinones, C.; Tamilselvan, A.; Prasad, P.V.V.; Jagadish, K.S.V. Agronomic and physiological responses to high temperature, drought, and elevated CO2 interactions in cereals. Adv. Agron. 2014, 127, 111–156. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yu, X.; Song, X.; Rosenqvist, E.; Wan, H.; Ottosen, C.O. Salinity, waterlogging and elevated [CO2] induced interactive and complicated responses in cultivated and wild tomato. J. Exp. Bot. 2022, 73, 5252–5263. [Google Scholar] [CrossRef] [PubMed]
- van der Kooi, C.J.; Reich, M.; Löw, M.; De Kok, L.J.; Tausz, M. Growth and yield stimulation under elevated CO2 and drought: A meta-analysis on crops. Environ. Exp. Bot. 2016, 122, 150–157. [Google Scholar] [CrossRef]
- Robredo, A.; Pérez-López, U.; de la Maza, H.S.; González-Moro, B.; Lacuesta, M.; Mena-Petite, A.; Muñoz-Rueda, A. Elevated CO2 alleviates the impact of drought on barley improving water status by lowering stomatal conductance and delaying its effects on photosynthesis. Environ. Exp. Bot. 2007, 59, 252–263. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhang, H.; Dong, B.; Shi, C.; Li, Y.; Zhai, H.; Liu, M. Effects of elevated CO2 concentration on growth and water use efficiency of winter wheat under two soil water regimes. Agric. Water Manag. 2010, 97, 1742–1748. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, X.; Wei, Z.; Liu, F. ABA-mediated modulation of elevated CO2 on stomatal response to drought. Curr. Opin. Plant Biol. 2020, 56, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Jiang, Y.; Zhou, G. Response and adaptation of photosynthesis, respiration, and antioxidant systems to elevated CO2 with environmental stress in plants. Front. Plant Sci. 2015, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, K.M.; Kota, V.R.; Reddy, T.P.; Rao, K.V.; Reddy, A.R. Amelioration of plant responses to drought under elevated CO2 by rejuvenating photosynthesis and nitrogen use efficiency: Implications for future climate-resilient crops. Photosynth. Res. 2021, 150, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Ulfat, A.; Shokat, S.; Li, X.; Fang, L.; Großkinsky, D.K.; Majid, S.A.; Roitsch, T.; Liu, F. Elevated carbon dioxide alleviates the negative impact of drought on wheat by modulating plant metabolism and physiology. Agric. Water Manag. 2021, 250, 1–10. [Google Scholar] [CrossRef]
- Li, F.; Guo, D.; Gao, X.; Zhao, X. Water Deficit Modulates the CO2 Fertilization Effect on Plant Gas Exchange and Leaf-Level Water Use Efficiency: A Meta-Analysis. Front. Plant Sci. 2021, 12, 775477. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, H.; Qiao, Y.; Wang, Y.; Cai, Z.; Dong, B.; Shi, C.; Liu, Y.; Li, X.; Liu, M. Effects of elevated CO2 on the growth, seed yield, and water use efficiency of soybean (Glycine max (L.) Merr.) under drought stress. Agric. Water Manag. 2013, 129, 105–112. [Google Scholar] [CrossRef]
- Wang, A.; Lam, S.K.; Hao, X.; Li, F.Y.; Zong, Y.; Wang, H.; Li, P. Elevated CO2 reduces the adverse effects of drought stress on a high-yielding soybean (Glycine max (L.) Merr.) cultivar by increasing water use efficiency. Plant Physiol. Biochem. 2018, 132, 660–665. [Google Scholar] [CrossRef]
- Reddy, A.R.; Rasineni, G.K.; Raghavendra, A.S. The impact of global elevated CO2 concentration on photosynthesis and plant productivity. Curr. Sci. 2010, 99, 46–57. [Google Scholar]
- Pan, C.; Ahammed, G.J.; Li, X.; Shi, K. Elevated CO2 improves photosynthesis under high temperature by attenuating the functional limitations to energy fluxes, electron transport and redox homeostasis in tomato leaves. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.R.; Seemann, J.R.; Coleman, J.S. Growth in elevated CO2 protects photosynthesis against high temperature damage. Plant Cell Environ. 2000, 23, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.; Perez, P.; Morcuende, R.; Martinez-Carrasco, R. Future CO2 concentrations, though not warmer temperatures, enhance wheat photosynthesis temperature responses. Physiol. Plant. 2008, 132, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Dusenge, M.E.; Madhavji, S.; Way, D.A. Contrasting acclimation responses to elevated CO2 and warming between an evergreen and a deciduous boreal conifer. Glob. Chang. Biol. 2020, 26, 3639–3657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, S.; Kjaer, K.H.; Ottosen, C.O.; Rosenqvist, E.; Kumari Sharma, D.; Wollenweber, B. The alleviating effect of elevated CO2 on heat stress susceptibility of two wheat (Triticum aestivum L.) cultivars. J. Agron. Crop Sci. 2013, 199, 340–350. [Google Scholar] [CrossRef]
- Chavan, S.G.; Duursma, R.A.; Tausz, M.; Ghannoum, O. Elevated CO2 alleviates the negative impact of heat stress on wheat physiology but not on grain yield. J. Exp. Bot. 2019, 70, 6447–6459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Högy, P.; Wu, X.; Schmid, I.; Wang, X.; Schulze, W.X.; Jiang, D.; Fangmeier, A. Physiological and proteomic evidence for the interactive effects of post-anthesis heat stress and elevated CO2 on wheat. Proteomics 2018, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Prasch, C.M.; Sonnewald, U. Signaling events in plants: Stress factors in combination change the picture. Environ. Exp. Bot. 2015, 114, 4–14. [Google Scholar] [CrossRef]
- Wingler, A.; Lea, P.J.; Quick, W.P.; Leegood, R.C. Photorespiration: Metabolic pathways and their role in stress protection. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2000, 355, 1517–1529. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Niyogi, K.K. Safety valves for photosynthesis. Curr. Opin. Plant Biol. 2000, 3, 455–460. [Google Scholar] [CrossRef]
- Berry, J.; Björkman, O. Photosynthetic response and adaptation to temperature in higher plants. Annu. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Salvucci, M.E.; Crafts-Brandner, S.J. Inhibition of photosynthesis by heat stress: The activation state of Rubisco as a limiting factor in photosynthesis. Physiol. Plant. 2004, 120, 179–186. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Boyle, D.L.; Welti, R.; Jagadish, S.V.K.; Prasad, P.V.V. Decreased photosynthetic rate under high temperature in wheat is due to lipid desaturation, oxidation, acylation, and damage of organelles. BMC Plant Biol. 2018, 18, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ristic, Z.; Bukovnik, U.; Momčilović, I.; Fu, J.; Vara Prasad, P.V. Heat-induced accumulation of chloroplast protein synthesis elongation factor, EF-Tu, in winter wheat. J. Plant Physiol. 2008, 165, 192–202. [Google Scholar] [CrossRef]
- Perdomo, J.A.; Capo-Bauca, S.; Carmo-Silva, E.; Galmes, J. Rubisco and Rubisco Activase Play an Important Role in the Biochemical Limitations of Photosynthesis in Rice, Wheat, and Maize under High Temperature and Water Deficit. Front. Plant Sci. 2017, 8, 490. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero, R.M.; Mittler, R.; Blumwald, E.; Zandalinas, S.I. Developing climate-resilient crops: Improving plant tolerance to stress combination. Plant J. 2022, 109, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Rogers, A.; Nelson, R.; Long, S.P. Testing the “source-sink” hypothesis of down-regulation of photosynthesis in elevated [CO2] in the field with single gene substitutions in Glycine max. Agric. For. Meteorol. 2004, 122, 85–94. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Kizildeniz, T.; Pascual, I.; Irigoyen, J.J.; Morales, F. Future CO2, warming and water deficit impact white and red Tempranillo grapevine: Photosynthetic acclimation to elevated CO2 and biomass allocation. Physiol. Plant. 2021, 172, 1779–1794. [Google Scholar] [CrossRef]
- Tausz-Posch, S.; Tausz, M.; Bourgault, M. Elevated [CO2] effects on crops: Advances in understanding acclimation, nitrogen dynamics and interactions with drought and other organisms. Plant Biol. 2020, 22, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Silva, E.; Andralojc, P.J.; Scales, J.C.; Driever, S.M.; Mead, A.; Lawson, T.; Raines, C.A.; Parry, M.A.J. Phenotyping of field-grown wheat in the UK highlights contribution of light response of photosynthesis and flag leaf longevity to grain yield. J. Exp. Bot. 2017, 68, 3473–3486. [Google Scholar] [CrossRef]
- Driever, S.; Lawson, T.; Andralojc, P.J.; Raines, C.A.; Parry, M. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes. J. Exp. Bot. 2014, 65, 4959–4973. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kristiansen, K.; Rosenqvist, E.; Liu, F. Elevated CO2 modulates the effects of drought and heat stress on plant water relations and grain yield in wheat. J. Agron. Crop Sci. 2019, 205, 362–371. [Google Scholar] [CrossRef]
- Fitzgerald, G.J.; Tausz, M.; O’Leary, G.; Mollah, M.R.; Tausz-Posch, S.; Seneweera, S.; Mock, I.; Löw, M.; Partington, D.L.; McNeil, D.; et al. Elevated atmospheric [CO2] can dramatically increase wheat yields in semi-arid environments and buffer against heat waves. Glob. Chang. Biol. 2016, 22, 2269–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelhakim, L.O.A.; Palma, C.F.F.; Zhou, R.; Wollenweber, B.; Ottosen, C.O.; Rosenqvist, E. The effect of individual and combined drought and heat stress under elevated CO2 on physiological responses in spring wheat genotypes. Plant Physiol. Biochem. 2021, 162, 301–314. [Google Scholar] [CrossRef]
- Abdelhakim, L.O.A.; Mendanha, T.; Palma, C.F.F.; Vrobel, O.; Stefelova, N.; Cavar Zeljkovic, S.; Tarkowski, P.; De Diego, N.; Wollenweber, B.; Rosenqvist, E.; et al. Elevated CO2 Improves the Physiology but Not the Final Yield in Spring Wheat Genotypes Subjected to Heat and Drought Stress During Anthesis. Front. Plant Sci. 2022, 13, 824476. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Wen, J.; Jensen, N.B.; Dos Santos, T.M.; Wu, Z.; Rosenqvist, E.; Ottosen, C.O. Interactive effects of elevated CO2 concentration and combined heat and drought stress on tomato photosynthesis. BMC Plant Biol. 2020, 20, 260. [Google Scholar] [CrossRef]
- Zinta, G.; AbdElgawad, H.; Domagalska, M.A.; Vergauwen, L.; Knapen, D.; Nijs, I.; Janssens, I.A.; Beemster, G.T.; Asard, H. Physiological, biochemical, and genome-wide transcriptional analysis reveals that elevated CO2 mitigates the impact of combined heat wave and drought stress in Arabidopsis thaliana at multiple organizational levels. Glob. Chang. Biol. 2014, 20, 3670–3685. [Google Scholar] [CrossRef] [PubMed]
- Abo Gamar, M.I.; Kisiala, A.; Emery, R.J.N.; Yeung, E.C.; Stone, S.L.; Qaderi, M.M. Elevated carbon dioxide decreases the adverse effects of higher temperature and drought stress by mitigating oxidative stress and improving water status in Arabidopsis thaliana. Planta 2019, 250, 1191–1214. [Google Scholar] [CrossRef]
- Abo Gamar, M.I.; Dixon, S.L.; Qaderi, M.M. Single and interactive effects of temperature, carbon dioxide and watering regime on plant growth and reproductive yield of two genotypes of Arabidopsis thaliana. Acta Physiol. Plant. 2021, 43, 1–16. [Google Scholar] [CrossRef]
- Diksaityte, A.; Virsile, A.; Zaltauskaite, J.; Januskaitiene, I.; Juozapaitiene, G. Growth and photosynthetic responses in Brassica napus differ during stress and recovery periods when exposed to combined heat, drought and elevated CO2. Plant Physiol. Biochem. 2019, 142, 59–72. [Google Scholar] [CrossRef]
- Broughton, K.J.; Smith, R.A.; Duursma, R.A.; Tan, D.K.Y.; Payton, P.; Bange, M.P.; Tissue, D.T. Warming alters the positive impact of elevated CO2 concentration on cotton growth and physiology during soil water deficit. Funct. Plant Biol. 2017, 44, 267–278. [Google Scholar] [CrossRef]
- Zhang, J.; Deng, L.; Jiang, H.; Peng, C.; Huang, C.; Zhang, M.; Zhang, X. The effects of elevated CO2, elevated O3, elevated temperature, and drought on plant leaf gas exchanges: A global meta-analysis of experimental studies. Environ. Sci. Pollut. Res. 2021, 28, 15274–15289. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Farfan-Vignolo, E.R.; de Vos, D.; Asard, H. Elevated CO2 mitigates drought and temperature-induced oxidative stress differently in grasses and legumes. Plant Sci. 2015, 231, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias De Oliveira, E.; Bramley, H.; Siddique, K.H.M.; Henty, S.; Berger, J.; Palta, J.A. Can elevated CO2 combined with high temperature ameliorate the effect of terminal drought in wheat? Funct. Plant Biol. 2013, 40, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Addy, J.W.G.; Ellis, R.H.; Macdonald, A.J.; Semenov, M.A.; Mead, A. The impact of weather and increased atmospheric CO2 from 1892 to 2016 on simulated yields of UK wheat. J. R. Soc. Interface 2021, 18, 20210250. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, V.R.; Devi, M.J.; Timlin, D.J. Impact of water stress under ambient and elevated carbon dioxide across three temperature regimes on soybean canopy gas exchange and productivity. Sci. Rep. 2021, 11, 16511. [Google Scholar] [CrossRef] [PubMed]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Aranjuelo, I.; Erice, G.; Nogués, S.; Morales, F.; Irigoyen, J.J.; Sánchez-Díaz, M. The mechanism(s) involved in the photoprotection of PSII at elevated CO2 in nodulated alfalfa plants. Environ. Exp. Bot. 2008, 64, 295–306. [Google Scholar] [CrossRef]
- Tausz-Posch, S.; Borowiak, K.; Dempsey, R.W.; Norton, R.M.; Seneweera, S.; Fitzgerald, G.J.; Tausz, M. The effect of elevated CO2 on photochemistry and antioxidative defence capacity in wheat depends on environmental growing conditions—A FACE study. Environ. Exp. Bot. 2013, 88, 81–92. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Beemster, G.T.; Janssens, I.A.; Asard, H. Future Climate CO2 Levels Mitigate Stress Impact on Plants: Increased Defense or Decreased Challenge? Front. Plant Sci. 2016, 7, 556. [Google Scholar] [CrossRef] [Green Version]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Xu, Z.; Shimizu, H.; Yagasaki, Y.; Ito, S.; Zheng, Y.; Zhou, G. Interactive effects of elevated CO2, drought, and warming on plants. J. Plant Growth Regul. 2013, 32, 692–707. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes, 1st ed.; ASA-CSSA-SSSA: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biol. 2017, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Naudts, K.; Van den Berge, J.; Janssens, I.A.; Nijs, I.; Ceulemans, R. Combined effects of warming and elevated CO2 on the impact of drought in grassland species. Plant Soil 2013, 369, 497–507. [Google Scholar] [CrossRef]
- Uddling, J.; Gelang-Alfredsson, J.; Karlsson, P.E.; Selldén, G.; Pleijel, H. Source–sink balance of wheat determines responsiveness of grain production to increased [CO2] and water supply. Agric. Ecosyst. Environ. 2008, 127, 215–222. [Google Scholar] [CrossRef]
- Yang, C.; Fraga, H.; van Ieperen, W.; Trindade, H.; Santos, J.A. Effects of climate change and adaptation options on winter wheat yield under rainfed Mediterranean conditions in southern Portugal. Clim. Change 2019, 154, 159–178. [Google Scholar] [CrossRef] [Green Version]
- Semenov, M.A.; Stratonovitch, P.; Alghabari, F.; Gooding, M.J. Adapting wheat in Europe for climate change. J. Cereal Sci. 2014, 59, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Tausz, M.; Norton, R.M.; Tausz-Posch, S.; Löw, M.; Seneweera, S.; O’Leary, G.; Armstrong, R.; Fitzgerald, G.J. Can additional N fertiliser ameliorate the elevated CO2 induced depression in grain and tissue N concentrations of wheat on a high soil N background? J. Agron. Crop Sci. 2017, 203, 574–583. [Google Scholar] [CrossRef] [Green Version]
- Albert, K.R.; Ro-Poulsen, H.; Mikkelsen, T.N.; Michelsen, A.; Van Der Linden, L.; Beier, C. Effects of elevated CO2, warming and drought episodes on plant carbon uptake in a temperate heath ecosystem are controlled by soil water status. Plant Cell Environ. 2011, 34, 1207–1222. [Google Scholar] [CrossRef]
- Allen, L.H.; Kimball, B.A.; Bunce, J.A.; Yoshimoto, M.; Harazono, Y.; Baker, J.T.; Boote, K.J.; White, J.W. Fluctuations of CO2 in Free-Air CO2 Enrichment (FACE) depress plant photosynthesis, growth, and yield. Agric. For. Meteorol. 2020, 284, 107899. [Google Scholar] [CrossRef]
- Macabuhay, A.; Houshmandfar, A.; Nuttall, J.; Fitzgerald, G.J.; Tausz, M.; Tausz-Posch, S. Can elevated CO2 buffer the effects of heat waves on wheat in a dryland cropping system? Environ. Exp. Bot. 2018, 155, 578–588. [Google Scholar] [CrossRef]
- Ben Mariem, S.; Soba, D.; Zhou, B.; Loladze, I.; Morales, F.; Aranjuelo, I. Climate Change, Crop Yields, and Grain Quality of C3 Cereals: A Meta-Analysis of [CO2], Temperature, and Drought Effects. Plants 2021, 10, 1052. [Google Scholar] [CrossRef]
- Ribeiro, R.V.; Ottosen, C.O.; Rosenqvist, E.; Medanha, T.; Abdelhakim, L.; Machado, E.C.; Struik, P.C. Elevated CO2 concentration increases photosynthetic sensitivity to nitrogen supply of sorghum in a genotype-dependent manner. Plant Physiol. Biochem. 2021, 168, 202–210. [Google Scholar] [CrossRef]
- Li, P.; Hao, X.; Aryal, M.; Thompson, M.; Seneweera, S. Elevated carbon dioxide and nitrogen supply affect photosynthesis and nitrogen partitioning of two wheat varieties. J. Plant Nutr. 2019, 42, 1290–1300. [Google Scholar] [CrossRef]
- Zhang, L.; Li, G.; Li, C.; Liu, F. CO2 elevation and nutrient patchiness interactively affect morphology, nitrogen uptake, partitioning and use efficiency of Nicotiana tabacum L. (tobacco) during anthesis. J. Agron. Crop Sci. 2022, 208, 523–535. [Google Scholar] [CrossRef]
- Pleijel, H.; Broberg, M.C.; Hogy, P.; Uddling, J. Nitrogen application is required to realize wheat yield stimulation by elevated CO2 but will not remove the CO2-induced reduction in grain protein concentration. Glob. Chang. Biol. 2019, 25, 1868–1876. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Kasemsap, P.; Rubio-Asensio, J.S. Rising atmospheric CO2 concentration inhibits nitrate assimilation in shoots but enhances it in roots of C3 plants. Physiol. Plant. 2020, 168, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Bloom, A.J.; Plant, R.E.; Foyer, C. Wheat grain yield decreased over the past 35 years, but protein content did not change. J. Exp. Bot. 2021, 72, 6811–6821. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Armstrong, R.; Panozzo, J.; Partington, D.; Fitzgerald, G. Can nitrogen fertiliser maintain wheat (Triticum aestivum) grain protein concentration in an elevated CO2 environment? Soil Res. 2017, 55, 518–523. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Crops | eCO2, ppm | Combination of Stress | Focus/Responses | Mitigation Effect of eCO2 under Combined Stress | References |
|---|---|---|---|---|---|
| Wheat | 800 | eCO2, heat, and drought | Physiological level and yield | eCO2 increased the grain yield by 20.8% in wheat. | [67] |
| Wheat | 550 FACE | Semi-arid environments | Morpho-physiological levels and yield | Mitigation was reflected in increased final yield. | [68] |
| Wheat | 800 | eCO2, heat, and drought | Physiological level | eCO2 increased photosynthesis and maintained higher Fv/Fm in some genotypes under combined stress compared to aCO2. | [69] |
| Wheat | 800 | eCO2, heat, and drought | Physiological, biochemical levels and yield | No eCO2 mitigation effect on the final yield despite enhanced WUE. | [70] |
| Tomato | 800 | eCO2, heat, and drought | Physiological level | No eCO2 mitigation due to increased severity of water deficit. | [71] |
| Arabidopsis | 730 | eCO2, heat, and drought | Physiological, biochemical, and genome-wide transcriptional levels | Mitigation is associated with reduced photorespiration and increased content of different antioxidant mechanisms. | [72] |
| Arabidopsis | 700 | eCO2, heat, and drought | Physiological and molecular levels | eCO2 increased stress tolerance under combined drought and heat by mitigating oxidative stress and improving the water status of plants. | [73] |
| Arabidopsis | 700 | eCO2, heat, and drought | Physiological and molecular levels | eCO2 partially mitigated the adverse effect of stresses on some traits in the selected genotypes. | [74] |
| Brassica napus | 800 | eCO2, heat, and drought | Morpho-physiological levels | Mitigation is associated with improving plant water relations, but not during the recovery. | [75] |
| Cotton | 640 | eCO2, heat, and drought | Morpho-physiological levels | eCO2 enhanced biomass and WUE, but not under high temperatures and rapid water deficit, due to the stomatal limitation. | [76] |
| Plants/trees | Meta-analysis | eCO2, eO3, heat, and drought | Physiological level | Improved WUE by eCO2 mitigated the negative stress impacts on photosynthesis to some extent. | [77] |
| Two types of grass; Lolium perenne, Poa pratensis. Two legumes; Medicago lupulina, Lotus corniculatus. | 615 | eCO2, heat, and drought | Physiological and biochemical levels | Enhanced physiological responses by eCO2 under stress in the grass, whereas mitigation through regulating lipid peroxidation and H2O2 levels were more pronounced in legumes. | [78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelhakim, L.O.A.; Zhou, R.; Ottosen, C.-O. Physiological Responses of Plants to Combined Drought and Heat under Elevated CO2. Agronomy 2022, 12, 2526. https://doi.org/10.3390/agronomy12102526
Abdelhakim LOA, Zhou R, Ottosen C-O. Physiological Responses of Plants to Combined Drought and Heat under Elevated CO2. Agronomy. 2022; 12(10):2526. https://doi.org/10.3390/agronomy12102526
Chicago/Turabian StyleAbdelhakim, Lamis Osama Anwar, Rong Zhou, and Carl-Otto Ottosen. 2022. "Physiological Responses of Plants to Combined Drought and Heat under Elevated CO2" Agronomy 12, no. 10: 2526. https://doi.org/10.3390/agronomy12102526