
Microbial Consortium Inoculum with Rock Minerals Increased Wheat Grain Yield, Nitrogen-Use Efficiency, and Protein Yield Due to Larger Root Growth and Architecture


Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Properties
2.2. Experimental Design and Treatments
2.3. Plant Growth Conditions and Harvest
2.4. Plant Growth and Yield
2.5. Root Sampling and Assessment
2.6. Nitrogen Concentration, N-Use Efficiency and Grain Protein Concentration
2.7. Statistical Analysis
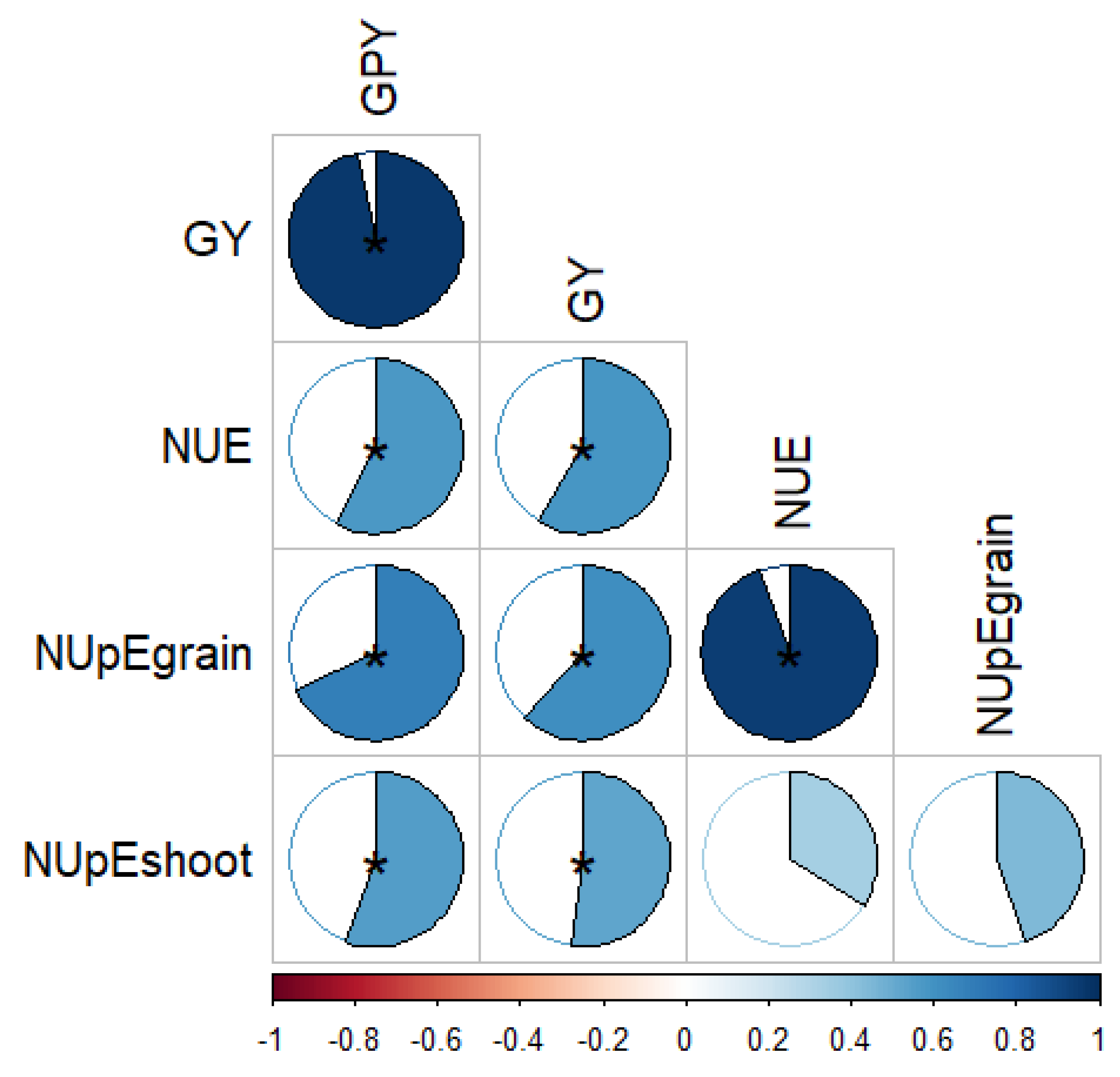
3. Results
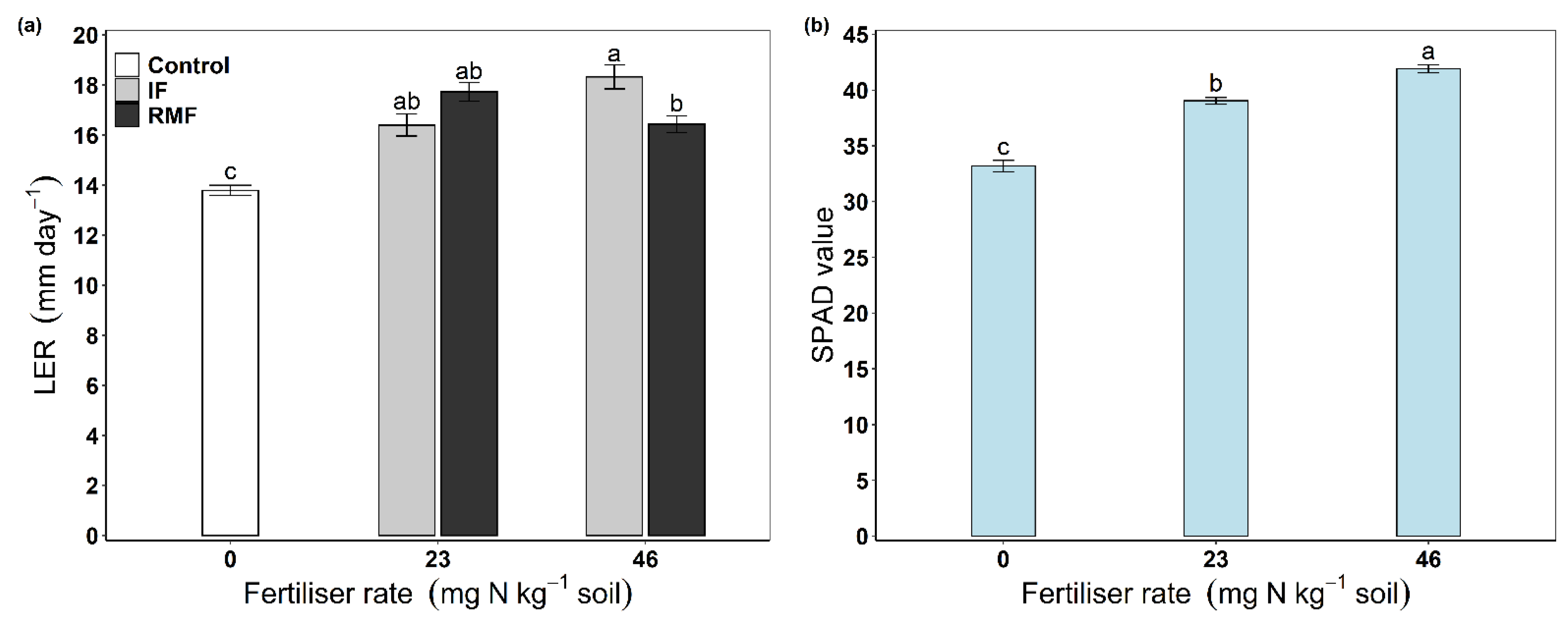
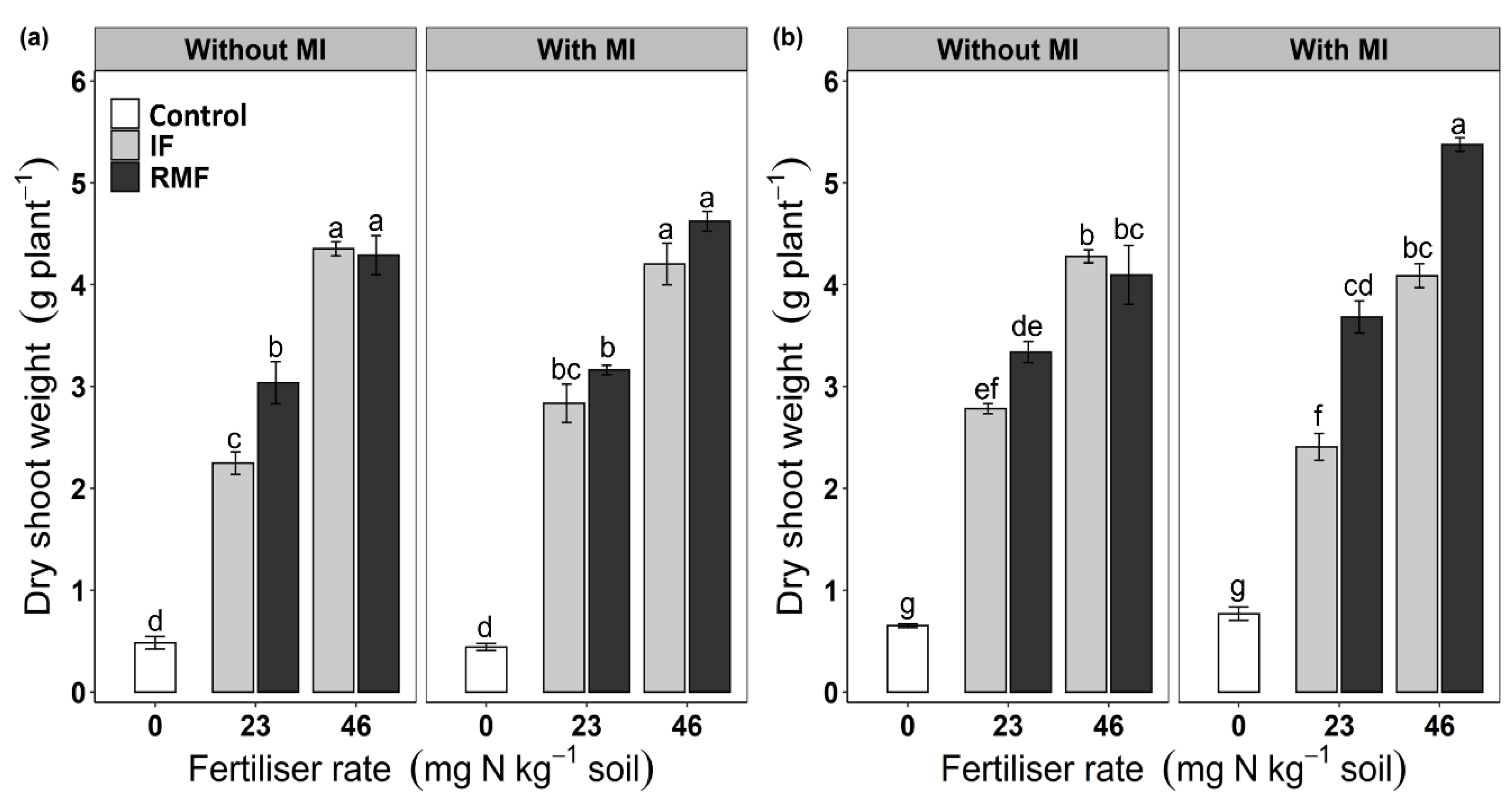
3.1. Leaf Extension Rate, Leaf Chlorophyll Content (SPAD Value) and Plant Biomass
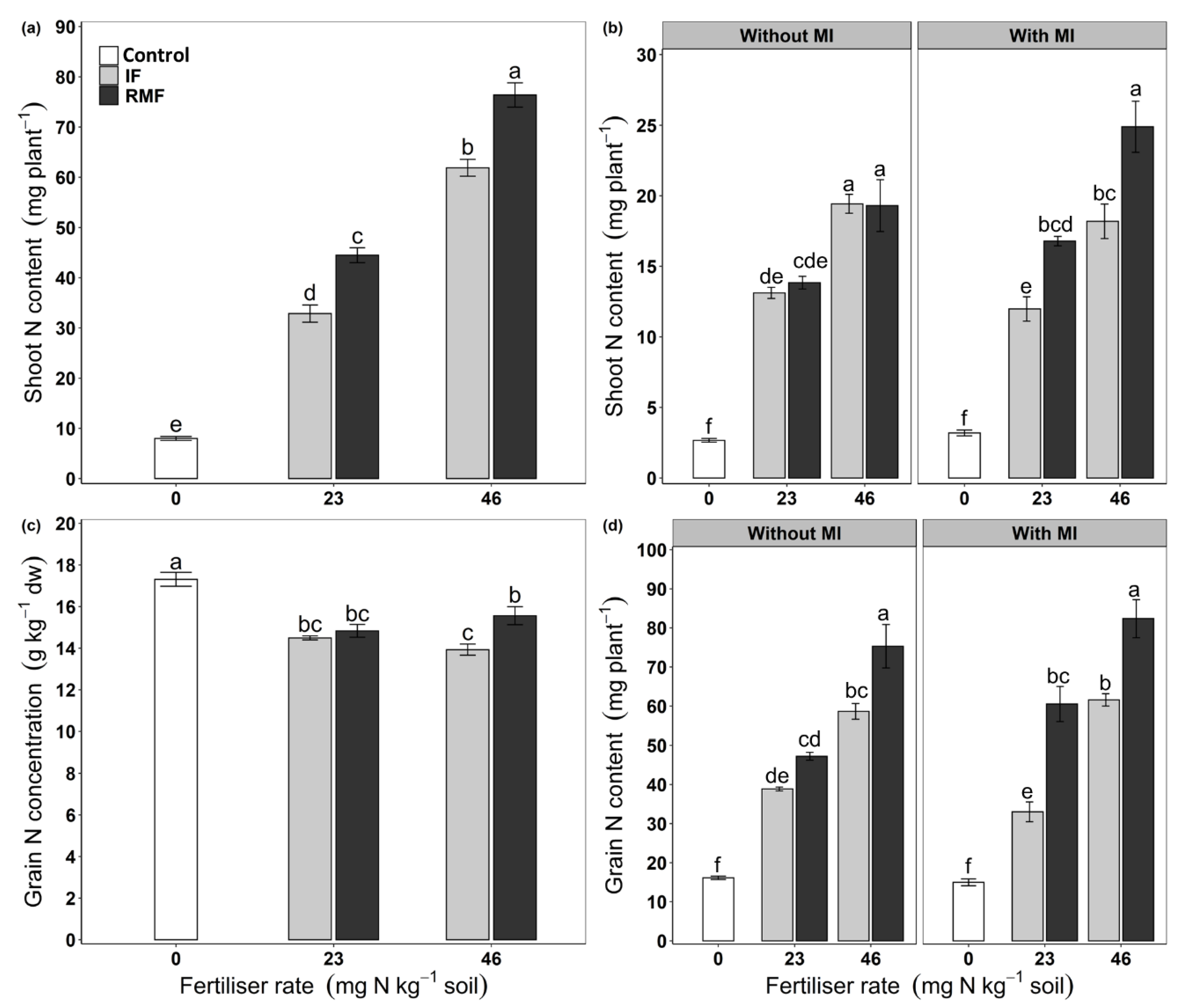
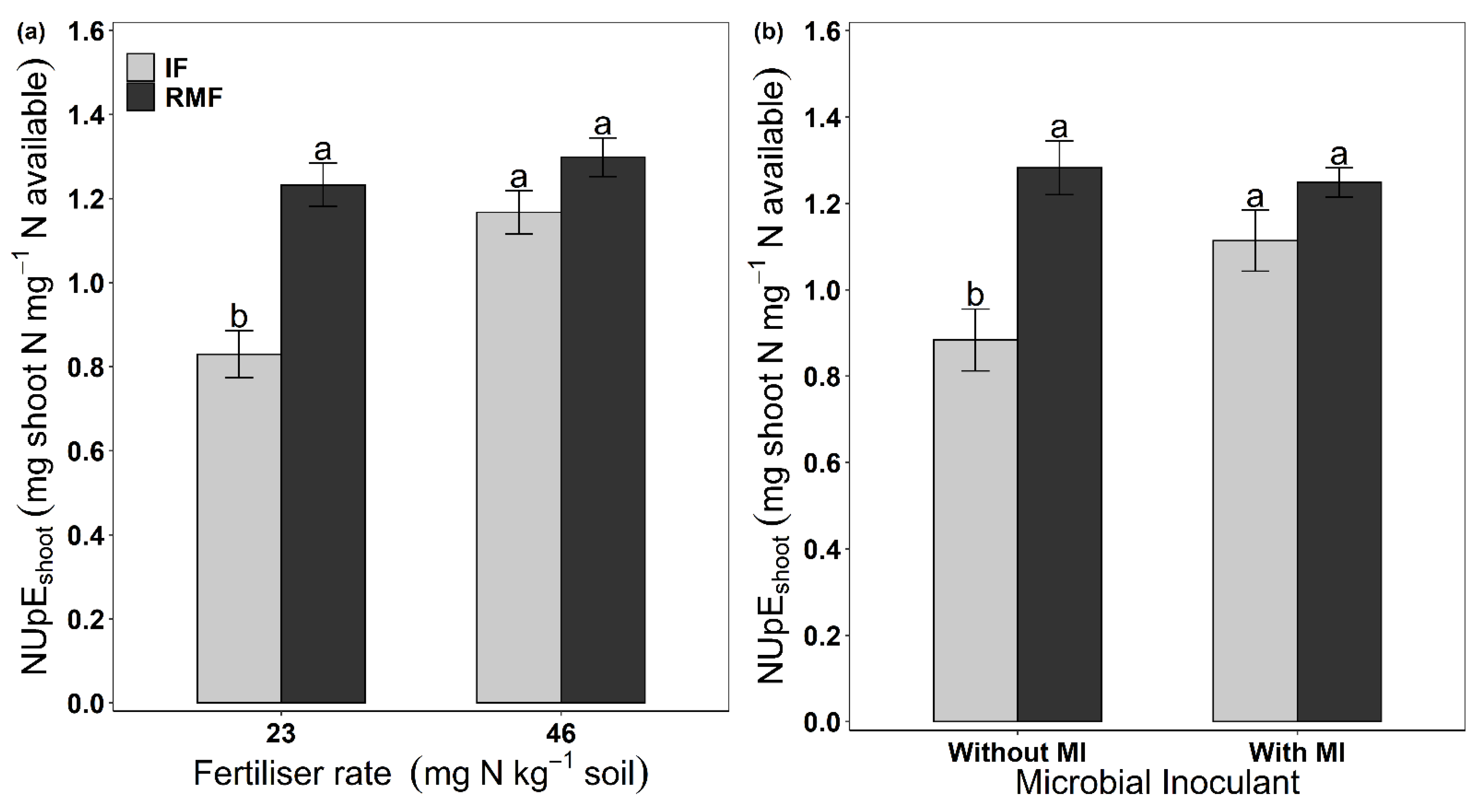
3.2. Nitrogen Content and N-Use Efficiency
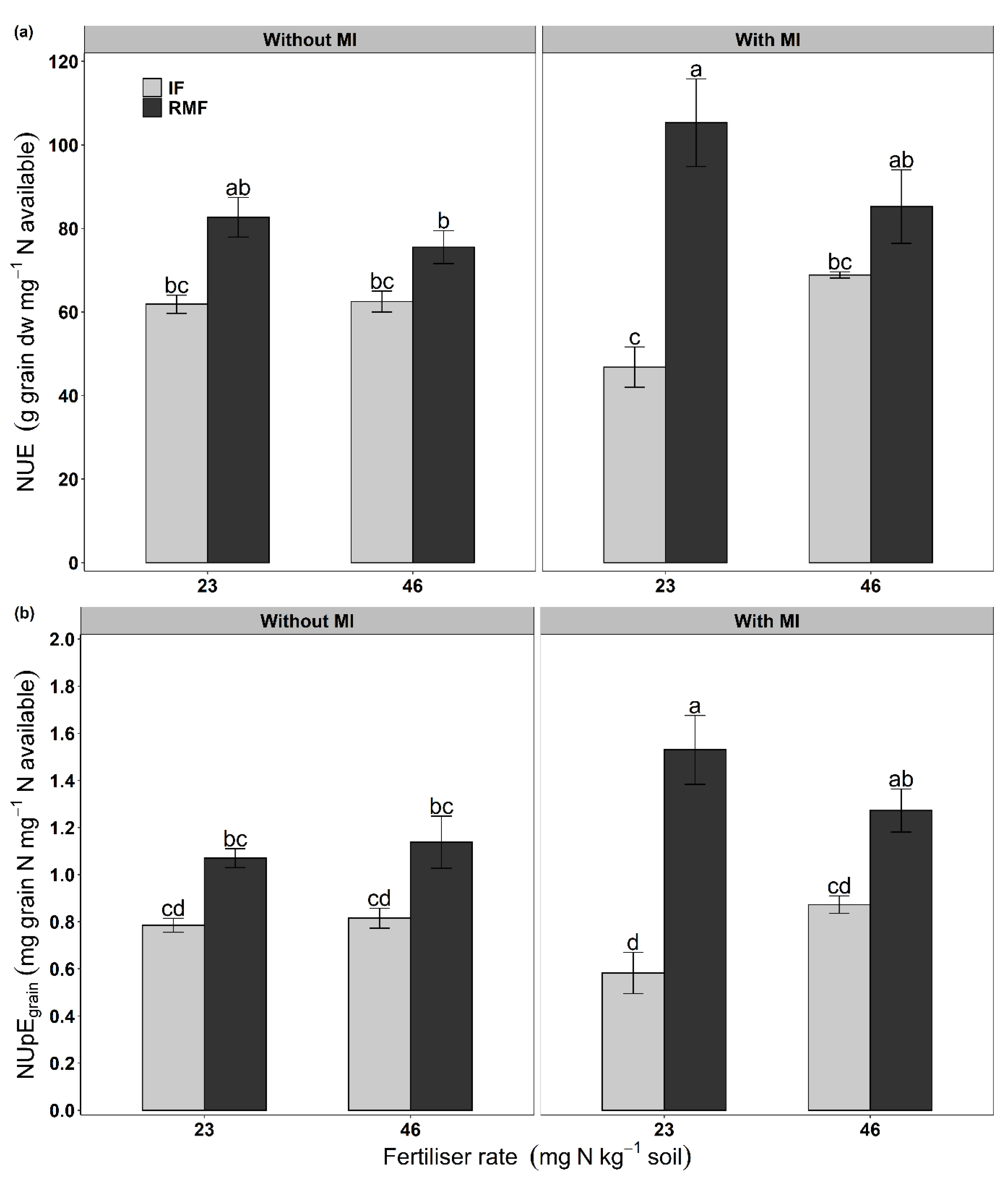
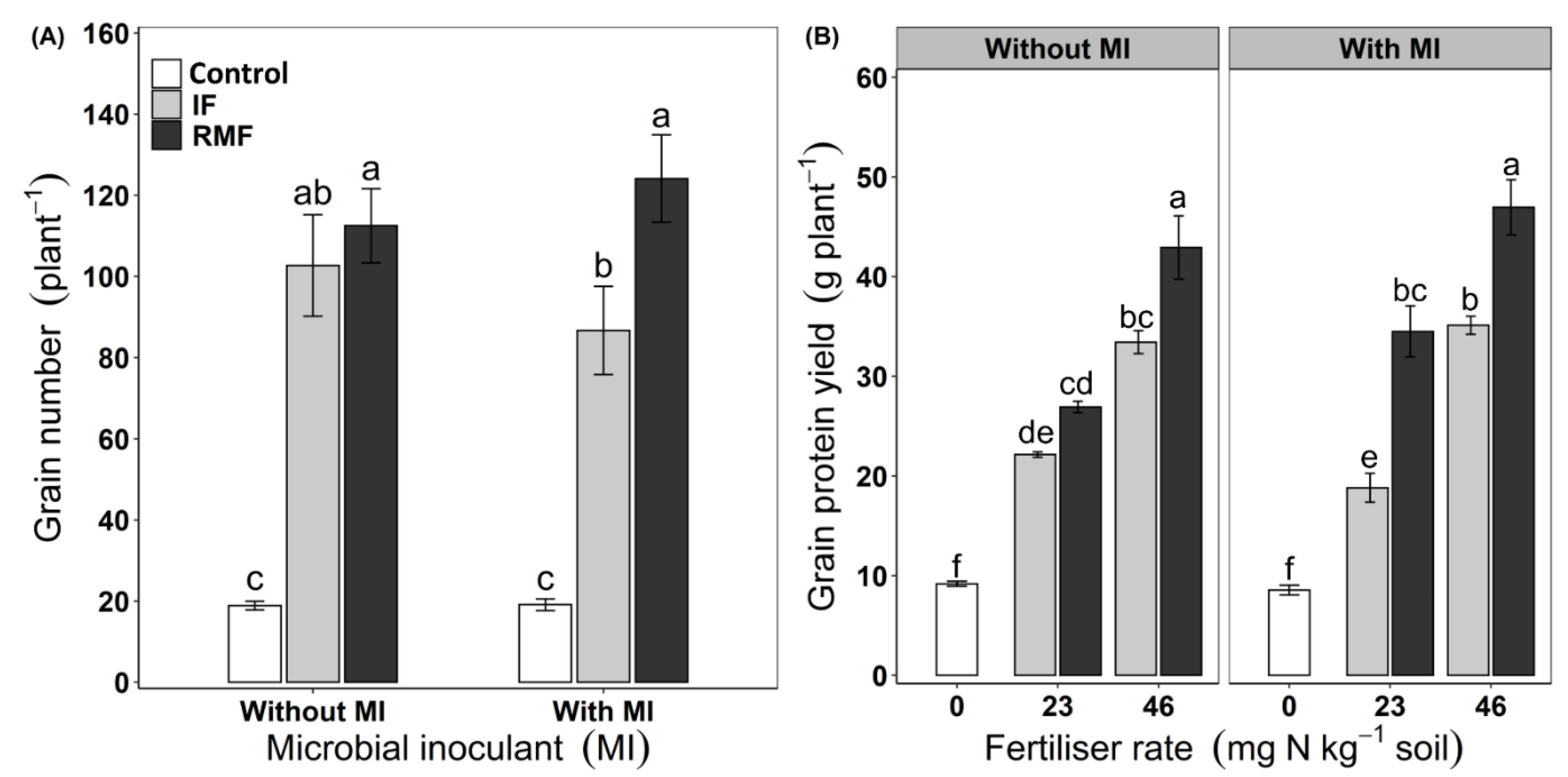
3.3. Grain Yield and Grain Protein Content
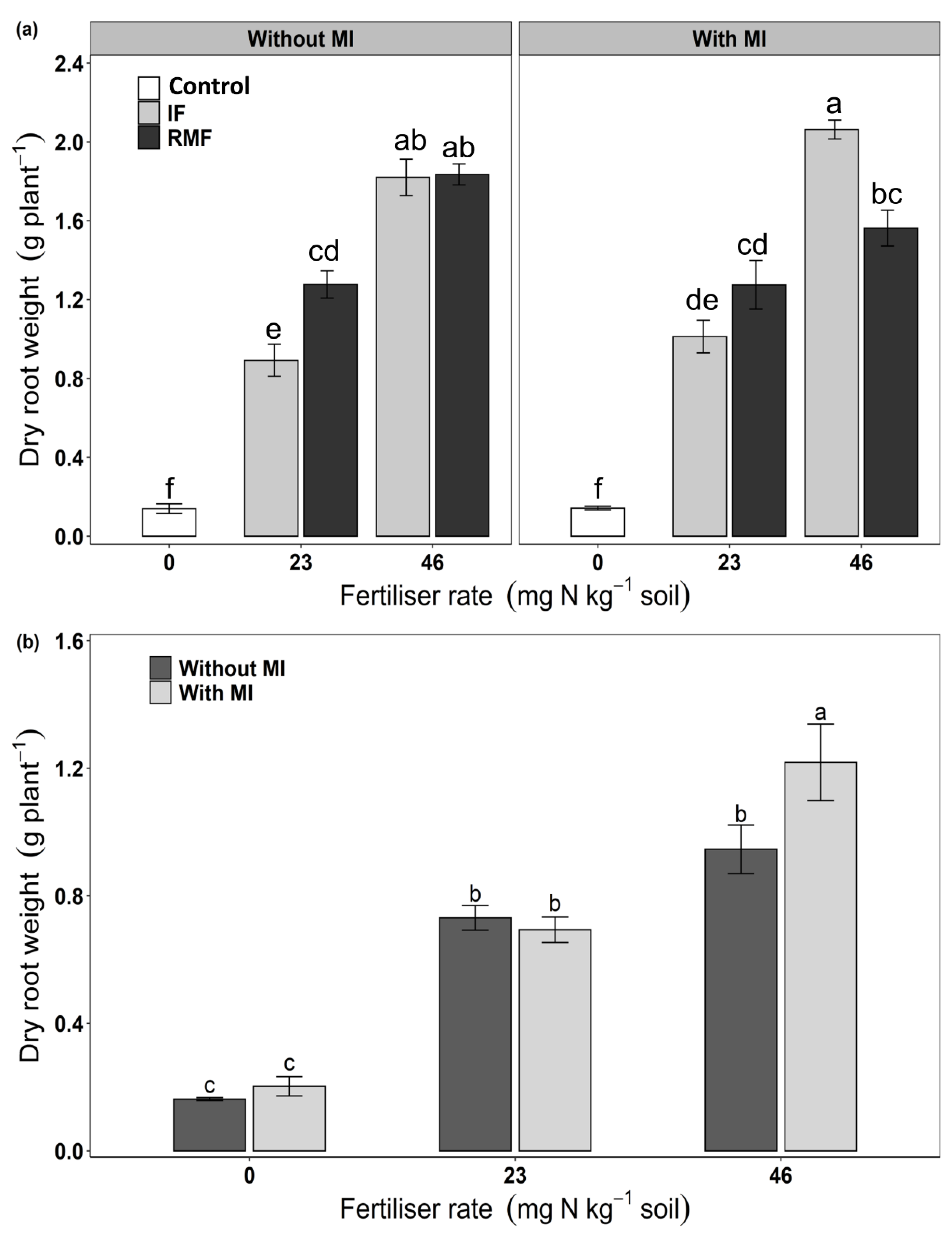
3.4. Root Morphology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, G.; Zhao, D.; Liu, S.; Liao, Y.; Han, J. Can controlled-release urea replace the split application of normal urea in China? A meta-analysis based on crop grain yield and nitrogen use efficiency. Field Crops Res. 2022, 275, 108343. [Google Scholar] [CrossRef]
- Teng, W.; He, X.; Tong, Y. Genetic control of efficient nitrogen use for high yield and grain protein concentration in wheat: A review. Plants 2022, 11, 492. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, S.; Liao, Y.; Han, J. A nitrogen fertilizer strategy for simultaneously increasing wheat grain yield and protein content: Mixed application of controlled-release urea and normal urea. Field Crops Res. 2022, 277, 108405. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. World Food and Agricultur-Statistical Yearbook 2021; FAO: Rome, Italy, 2021. [Google Scholar]
- Manschadi, A.M.; Soltani, A. Variation in traits contributing to improved use of nitrogen in wheat: Implications for genotype by environment interaction. Field Crops Res. 2021, 270, 108211. [Google Scholar] [CrossRef]
- Koryagin, Y.; Kulikova, E.; Efremova, S.; Sukhova, N. The influence of microbiological fertilisers on the productivity and quality of winter wheat. Plant Soil Environ. 2020, 66, 564–568. [Google Scholar] [CrossRef]
- Mălinaş, A.; Vidican, R.; Rotar, I.; Mălinaş, C.; Moldovan, C.M.; Proorocu, M. Current status and future prospective for nitrogen use efficiency in wheat (Triticum aestivum L.). Plants 2022, 11, 217. [Google Scholar] [CrossRef]
- Lollato, R.P.; Jaenisch, B.R.; Silva, S.R. Genotype-specific nitrogen uptake dynamics and fertilizer management explain contrasting wheat protein concentration. Crop Sci. 2021, 61, 2048–2066. [Google Scholar] [CrossRef]
- Tabak, M.; Lepiarczyk, A.; Filipek-Mazur, B.; Lisowska, A. Efficiency of nitrogen fertilization of winter wheat depending on sulfur fertilization. Agronomy 2020, 10, 1304. [Google Scholar] [CrossRef]
- Dawson, J.C.; Huggins, D.R.; Jones, S.S. Characterizing nitrogen use efficiency in natural and agricultural ecosystems to improve the performance of cereal crops in low-input and organic agricultural systems. Field Crops Res. 2008, 107, 89–101. [Google Scholar] [CrossRef]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef]
- Trenkel, M.E. Slow-and Controlled-Release and Stabilized Fertilizers: An Option For Enhancing Nutrient Use Efficiency in Agriculture; IFA, International fertilizer industry association: Paris, France, 2010. [Google Scholar]
- Ghafoor, I.; Habib-ur-Rahman, M.; Ali, M.; Afzal, M.; Ahmed, W.; Gaiser, T.; Ghaffar, A. Slow-release nitrogen fertilizers enhance growth, yield, NUE in wheat crop and reduce nitrogen losses under an arid environment. Environ. Sci. Pollut. Res. 2021, 28, 43528–43543. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhan, X.; Han, X.; Chen, K.; Peng, J.; Wang, X.; Shang, D. Slow-release nitrogen fertiliser suitable for one-time fertilisation of spring maize in Northeast China. Plant Soil Environ. 2021, 67, 164–172. [Google Scholar] [CrossRef]
- Vejan, P.; Khadiran, T.; Abdullah, R.; Ahmad, N. Controlled release fertilizer: A review on developments, applications and potential in agriculture. J. Control. Release 2021, 339, 321–334. [Google Scholar] [CrossRef]
- Beig, B.; Niazi, M.B.K.; Jahan, Z.; Hussain, A.; Zia, M.H.; Mehran, M.T. Coating materials for slow release of nitrogen from urea fertilizer: A review. J. Plant Nutr. 2020, 43, 1510–1533. [Google Scholar] [CrossRef]
- De Santis, M.A.; Giuliani, M.M.; Flagella, Z.; Reyneri, A.; Blandino, M. Impact of nitrogen fertilisation strategies on the protein content, gluten composition and rheological properties of wheat for biscuit production. Field Crops Res. 2020, 254, 1–10. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Enhanced plant nutrient use efficiency with PGPR and AMF in an integrated nutrient management system. Can. J. Microbiol. 2008, 54, 876–886. [Google Scholar] [CrossRef] [Green Version]
- Assainar, S.K.; Abbott, L.K.; Mickan, B.S.; Storer, P.J.; Whiteley, A.S.; Siddique, K.H.M.; Solaiman, Z.M. Polymer-coated rock mineral fertilizer has potential to substitute soluble fertilizer for increasing growth, nutrient uptake, and yield of wheat. Biol. Fertil. Soils 2020, 56, 381–394. [Google Scholar] [CrossRef]
- Assainar, S.K.; Abbott, L.K.; Mickan, B.S.; Whiteley, A.S.; Siddique, K.H.M.; Solaiman, Z.M. Response of wheat to a multiple species microbial inoculant compared to fertilizer application. Front. Plant Sci. 2018, 9, 1601. [Google Scholar] [CrossRef] [Green Version]
- Tshewang, S.; Rengel, Z.; Siddique, K.H.M.; Solaiman, Z.M. Growth and nutrient uptake of temperate perennial pastures are influenced by grass species and fertilisation with a microbial consortium inoculant. J. Plant Nutr. Soil Sci. 2020, 183, 530–538. [Google Scholar] [CrossRef]
- Mickan, B.S.; Alsharmani, A.R.; Solaiman, Z.M.; Leopold, M.; Abbott, L.K. Plant-dependent soil bacterial responses following amendment with a multispecies microbial biostimulant compared to rock mineral and chemical fertilizers. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G. Proposal for the division of plant growth-promoting Rhizobacteria into two classifications: Biocontrol-PGPB (Plant Growth-Promoting Bacteria) and PGPB. Soil Biol. Biochem. 1998, 30, 1225–1228. [Google Scholar] [CrossRef]
- Owen, D.; Williams, A.P.; Griffith, G.W.; Withers, P.J.A. Use of commercial bio-inoculants to increase agricultural production through improved phosphrous acquisition. Appl. Soil Ecol. 2015, 86, 41–54. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Caradonia, F.; Battaglia, V.; Righi, L.; Pascali, G.; La Torre, A. Plant biostimulant regulatory framework: Prospects in Europe and current situation at international level. J. Plant Growth Regul. 2019, 38, 438–448. [Google Scholar] [CrossRef]
- Sible, C.N.; Seebauer, J.R.; Below, F.E. Plant biostimulants: A categorical review, their implications for row crop production, and relation to soil health Indicators. Agronomy 2021, 11, 1297. [Google Scholar] [CrossRef]
- Tshewang, S.; Rengel, Z.; Siddique, K.H.M.; Solaiman, Z.M. Microbial consortium inoculant increases pasture grasses yield in low-phosphorus soil by influencing root morphology, rhizosphere carboxylate exudation and mycorrhizal colonisation. J. Sci. Food Agric. 2021, 102, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.; Flottmann, S.J.; Stefanovia, K.T.; Colmer, T.D. Approaches to scheduling water allocations to kikuyugrass grown on a water repellent soil in drying-climate. Agric. Water Manag. 2020, 230. [Google Scholar] [CrossRef]
- Rayment, G.E.; Lyons, D.J.; Shelley, B. Soil chemical methods-Australasia. In Australian Soil and Land Survey Handbooks Series; CSIRO Publishing: Melbourne, VIC, Australia, 2011. [Google Scholar]
- Tshering, K.; Rengel, Z.; Storer, P.J.; Solaiman, Z.M. Novel rock mineral fertiliser application with microbial consurtium inoculant enhances growth, yield and grain protein content of wheat (Triticum aestivum L.) in sandy soil. Arch. Agron. Soil Sci. 2022, 1–16. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Malik, A.I.; Veres, S.; Rengel, Z. Differential nitrogen-use efficiency in wheat parents of doubled-haploid mapping populations. Plant Soil 2016, 408, 311–325. [Google Scholar] [CrossRef]
- Xu, Q.; Krishnan, S.; Merewitz, E.; Xu, J.; Huang, B. Gibberellin-regulation and genetic variations in leaf elongation for tall fescue in association with differential gene expression controlling cell expansion. Sci. Rep. 2016, 6, 30258. [Google Scholar] [CrossRef]
- Khan, M.S.A.; Abbott, L.K.; Solaiman, Z.M.; Mawson, P.R.; Waite, I.S.; Jenkins, S.N. Complementary effect of zoo compost with mineral nitrogen fertilization increases wheat yield and nutrition in a low-nutrient soil. Pedosphere 2022, 32, 339–347. [Google Scholar] [CrossRef]
- Jensen, S.M.; Svensgaard, J.; Ritz, C. Estimation of the harvest index and the relative water content—Two examples of composite variables in agronomy. Eur. J. Agron. 2020, 112, 1–8. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Kibria, M.G.; Barton, L.; Rengel, Z. Foliar application of magnesium mitigates soil acidity stress in wheat. J. Agron. Crop Sci. 2021, 207, 378–389. [Google Scholar] [CrossRef]
- Bouma, T.J.; Nielsen, K.L.; Koutstaal, B.A.S. Sample preparation and scanning protocol for computerised analysis of root length and diameter. Plant Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- Dhillon, J.; Eickhoff, E.; Aula, L.; Omara, P.; Weymeyer, G.; Nambi, E.; Oyebiyi, F.; Carpenter, T.; Raun, W. Nitrogen management impact on winter wheat grain yield and estimated plant nitrogen loss. Agron. J. 2020, 112, 564–577. [Google Scholar] [CrossRef] [Green Version]
- Metho, L.A.; Taylor, J.R.N.; Hammes, P.S.; Randall, P.G. Effects of cultivar and soil fertility on grain protein yield, grain protein content, flour yield and breadmaking quality of wheat. J. Sci. Food Agric. 1999, 79, 1823–1831. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Griffiths, S. Exploiting genetic variation in nitrogen use efficiency for cereal crop improvement. Curr. Opin. Plant Biol. 2019, 49, 35–42. [Google Scholar] [CrossRef]
- Duncan, E.G.; O’ Sullivan, C.A.; Roper, M.M.; Palta, J.; Whisson, K.; Peoples, M.B. Yield and nitrogen use efficiency of wheat increased with root length and biomass due to nitrogen, phosphorus, and potassium interactions. J. Plant Nutri. Soil Sci. 2018, 181, 364–373. [Google Scholar] [CrossRef]
- Yang, Y.-C.; Zhang, M.; Zheng, L.; Cheng, D.-D.; Liu, M.; Geng, Y.-Q. Controlled release urea improved nitrogen use efficiency, yield, and quality of wheat. Agron. J. 2011, 103, 479–485. [Google Scholar] [CrossRef]
- Li, G.-H.; Cheng, G.-G.; Lu, W.-P.; Lu, D.-L. Differences of yield and nitrogen use efficiency under different applications of slow release fertilizer in spring maize. J. Integr. Agric. 2021, 20, 554–564. [Google Scholar] [CrossRef]
- Hett, J.; Neuhoff, D.; Döring, T.F.; Masoero, G.; Ercole, E.; Bevivino, A. Effects of multi-species mcrobial inoculants on early wheat growth and litterbag microbial activity. Agronomy 2022, 12, 899. [Google Scholar] [CrossRef]
- Rekowski, A.; Wimmer, M.A.; Hitzmann, B.; Hermannseder, B.; Hahn, H.; Zörb, C. Application of urease inhibitor improves protein composition and bread-baking quality of urea fertilized winter wheat. J. Plant Nutr. Soil Sci. 2020, 183, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Turner, N.C.; Poole, M.L. Source-sink balance and manipulating sink-source relations of wheat indicate that the yield potential of wheat is sink-limited in high-rainfall zones. Crop Pasture Sci. 2010, 61, 852–861. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, Z.; Zhang, M.; Shi, Y.; Zhu, Q.; Sun, Y.; Zhou, H.; Li, C.; Yang, Y.; Geng, J. Improving crop yields, nitrogen use efficiencies, and profits by using mixtures of coated controlled-released and uncoated urea in a wheat-maize system. Field Crops Res. 2017, 205, 106–115. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, M.; Zheng, G.; Yao, Y.; Tao, R.; Zhu, M.; Ding, J.; Li, C.; Guo, W.; Zhu, X. Twice-split application of controlled-release nitrogen fertilizer met the nitrogen demand of winter wheat. Field Crops Res. 2021, 267, 108163. [Google Scholar] [CrossRef]
- Geng, J.; Chen, J.; Sun, Y.; Zheng, W.; Tian, X.; Yang, Y.; Li, C.; Zhang, M. Controlled release urea improved nitrogen use efficiency and yield of wheat and corn. Agron. J. 2016, 108, 1666–1673. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef] [Green Version]
- Farmaha, B.S.; Sims, A.L. The influence of polymer-coated urea and urea fertilizer mixtures on spring wheat protein concentrations and economic returns. Agron. J. 2013, 105, 1328–1334. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Yao, C.; Zhang, Z.; Liu, Y.; Zhang, Y. Micro-sprinkling irrigation simultaneously improves grain yield and protein concentration of winter wheat in the North China Plain. Crop J. 2021, 9, 1397–1407. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Ft | Fr | MI | Ft × Fr | Ft × MI | Fr × MI | Ft × Fr × MI |
|---|---|---|---|---|---|---|---|
| Anthesis | |||||||
| Shoot dry weight | ** | *** | ns | * | ns | ns | * |
| Root dry weight | ns | *** | ns | *** | ns | * | * |
| Leaf chlorophyll content (SPAD value) | ns | *** | ns | ns | ns | ns | ns |
| 3rd leaf extension rate | ns | ns | ns | *** | ns | ns | ns |
| Shoot N concentration | ns | *** | * | ns | ns | ns | ns |
| Shoot N content | *** | *** | ns | *** | ns | ns | ns |
| Maturity | |||||||
| Shoot dry weight | *** | *** | ** | *** | *** | ** | *** |
| Root dry weight | ns | *** | ns | ns | ns | * | ns |
| Shoot N concentration | ns | * | ns | ns | ns | ns | ns |
| Shoot N content | *** | *** | * | * | ** | ns | * |
| Grain N concentration | * | *** | ns | * | ns | ns | ns |
| Grain N content | *** | *** | ns | *** | * | ns | * |
| NUE parameters | |||||||
| NUE (mg grain mg−1 N available) | *** | ns | ns | ** | * | ns | * |
| NUpEshoot (mg N in shoot mg−1 N available) | *** | *** | ** | ** | ** | ns | ns |
| NUpEgrain (mg N in grain mg−1 N available) | *** | ns | ns | * | ** | ns | * |
| Factor | Factor Levels | Shoot N Concentration (g kg−1 dw) | |
|---|---|---|---|
| Anthesis (Zadoks 65) | Maturity (Zadoks 92) | ||
| Fertiliser rate (mg N kg−1 soil) | 0 | 17 ± 0.7 a | 4 ± 0.1 b |
| 23 | 13 ± 0.3 b | 5 ± 0.1 a | |
| 46 | 16 ± 0.6 a | 5 ± 0.1 a | |
| p value | <0.001 | 0.012 | |
| Microbial inoculant (MI) | With MI | 17 ± 0.6 a | |
| Without MI | 15 ± 0.4 b | ||
| p value | 0.014 | 0.377 | |
| Variables | Ft | Fr | MI | Ft × Fr | Ft × MI | Fr × MI | Ft × Fr × MI |
|---|---|---|---|---|---|---|---|
| Grain number | *** | *** | ns | ** | * | ns | ns |
| Grain yield | *** | *** | ns | *** | ns | ns | ns |
| 1000-grain weight | ns | *** | ns | ns | ns | ns | ns |
| Harvest index | ns | *** | ns | ns | ns | ns | ns |
| Grain protein concentration | ** | *** | ns | * | ns | ns | ns |
| Grain protein yield | *** | *** | ns | *** | * | ns | * |
| Total root length | ns | *** | ns | *** | ns | ns | ns |
| Specific root length | ns | *** | ns | ns | ns | ns | ns |
| Root surface area | ns | *** | ns | *** | ns | ns | ns |
| Average root diameter (D) | ns | *** | ns | ns | ns | ns | ns |
| Fine root (0 < D ≤ 0.2 mm) length | ns | *** | ns | *** | ns | ns | ns |
| Coarse root (D > 0.2 mm) length | ns | *** | ns | ns | ns | ns | ns |
| Factor Level (mg N kg−1 Soil) | Grain Number (Plant−1) | Grain Yield (g Plant−1) | Grain Protein Concentration (g kg−1 dw) |
|---|---|---|---|
| Control | 19 ± 0.8 d | 0.9 ± 0.03 e | 98 ± 1.9 a |
| IF at 23 | 66 ± 3.5 c | 2.5 ± 0.11 d | 83 ± 0.6 bc |
| RMF at 23 | 101 ± 6.4 b | 3.6 ± 0.20 c | 85 ± 1.8 bc |
| IF at 46 | 123 ± 6.7 a | 4.3 ± 0.08 b | 79 ± 1.5 c |
| RMF at 46 | 136 ± 8.9 a | 5.1 ± 0.25 a | 89 ± 2.5 b |
| p value | 0.008 | <0.001 | 0.025 |
| Factor Level (mg N kg−1 Soil) | Total Root Length (m pot−1) | Root Surface Area (cm2 pot−1) | Fine Root (0 < D ≤ 0.2 mm) Length (m pot−1) |
|---|---|---|---|
| Control | 147 ± 14 e | 85 ± 8 d | 125 ± 12 d |
| IF at 23 | 465 ± 40 d | 499 ± 41 c | 330 ± 30 c |
| RMF at 23 | 665 ± 24 c | 730 ± 35 b | 487 ± 24 b |
| IF at 46 | 961 ± 34 a | 1052 ± 60 a | 674 ± 47 a |
| RMF at 46 | 802 ± 43 b | 900 ± 75 ab | 528 ± 58 ab |
| p value | <0.001 | <0.001 | <0.001 |
| Fertiliser Rate (mg N kg−1 Soil) | Specific Root Length (m g−1) | Average Root Diameter (mm) | Coarse Root (D > 0.2 mm) Length (m pot−1) |
|---|---|---|---|
| 0 | 529 ± 24 a | 0.18 ± 0.003 b | 22 ± 2 c |
| 23 | 254 ± 9 b | 0.23 ± 0.002 a | 157 ± 9 b |
| 46 | 242 ± 6 b | 0.24 ± 0.004 a | 261 ± 9 a |
| p value | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tshering, K.; Rengel, Z.; Storer, P.; Solaiman, Z.M. Microbial Consortium Inoculum with Rock Minerals Increased Wheat Grain Yield, Nitrogen-Use Efficiency, and Protein Yield Due to Larger Root Growth and Architecture. Agronomy 2022, 12, 2481. https://doi.org/10.3390/agronomy12102481
Tshering K, Rengel Z, Storer P, Solaiman ZM. Microbial Consortium Inoculum with Rock Minerals Increased Wheat Grain Yield, Nitrogen-Use Efficiency, and Protein Yield Due to Larger Root Growth and Architecture. Agronomy. 2022; 12(10):2481. https://doi.org/10.3390/agronomy12102481
Chicago/Turabian StyleTshering, Kinley, Zed Rengel, Paul Storer, and Zakaria M. Solaiman. 2022. "Microbial Consortium Inoculum with Rock Minerals Increased Wheat Grain Yield, Nitrogen-Use Efficiency, and Protein Yield Due to Larger Root Growth and Architecture" Agronomy 12, no. 10: 2481. https://doi.org/10.3390/agronomy12102481