
A Physiologically Based ODE Model for an Old Pest: Modeling Life Cycle and Population Dynamics of Bactrocera oleae (Rossi)

 ,
, 


Abstract
:1. Introduction
2. Materials and Methods
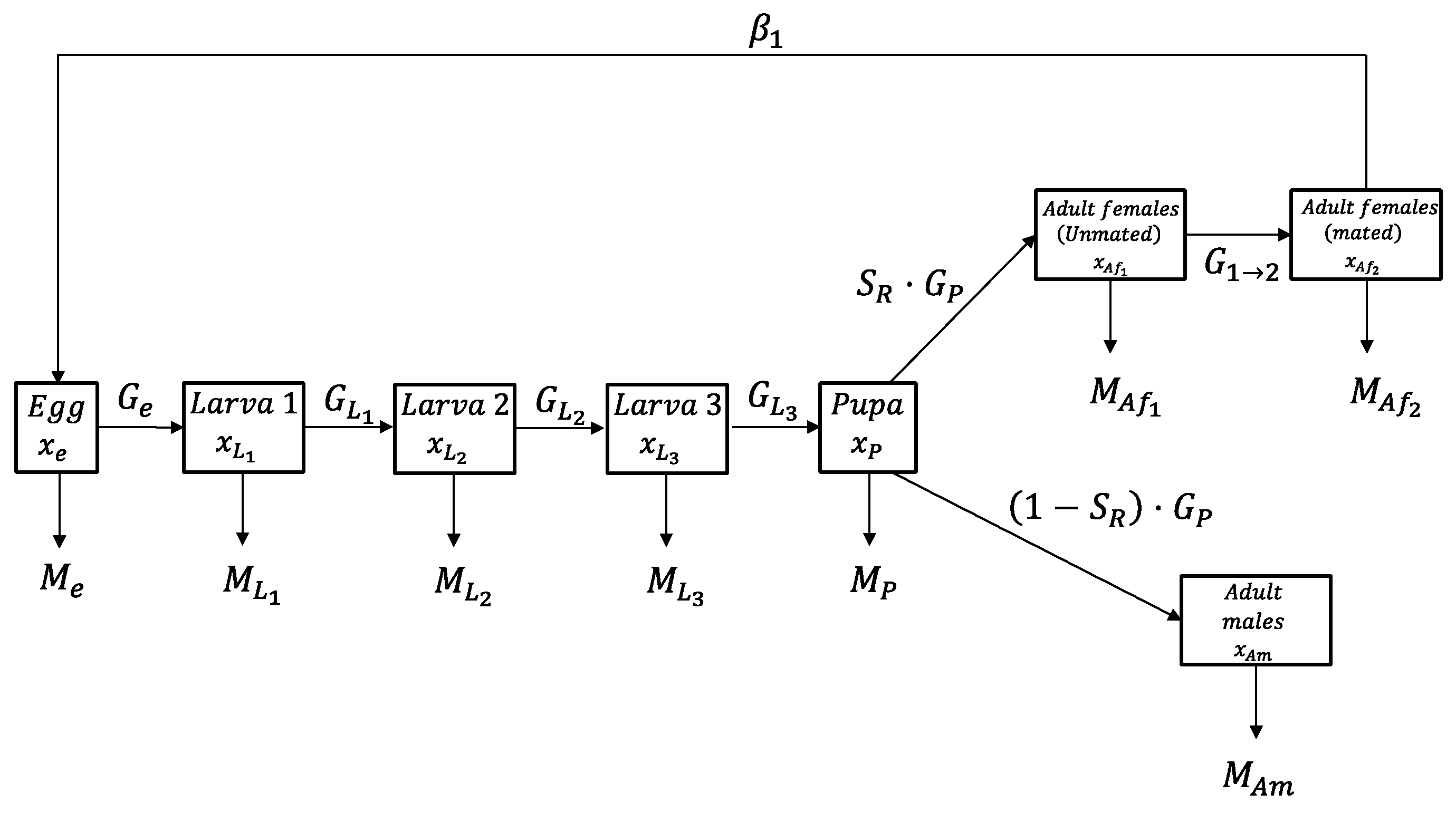
2.1. Population Density Model
2.2. Development, Fertility, and Mortality Rate Functions
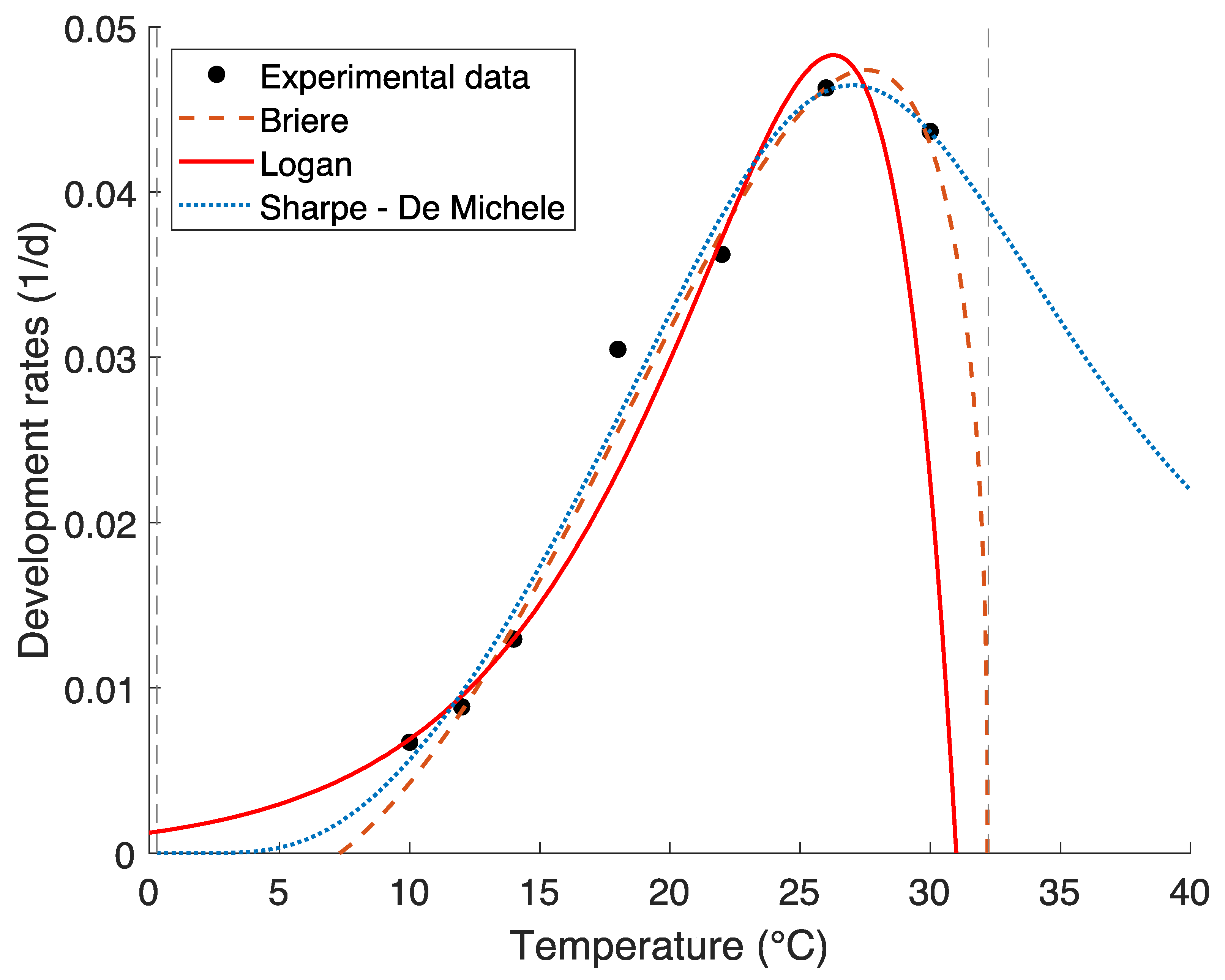
2.2.1. Development Rate Functions
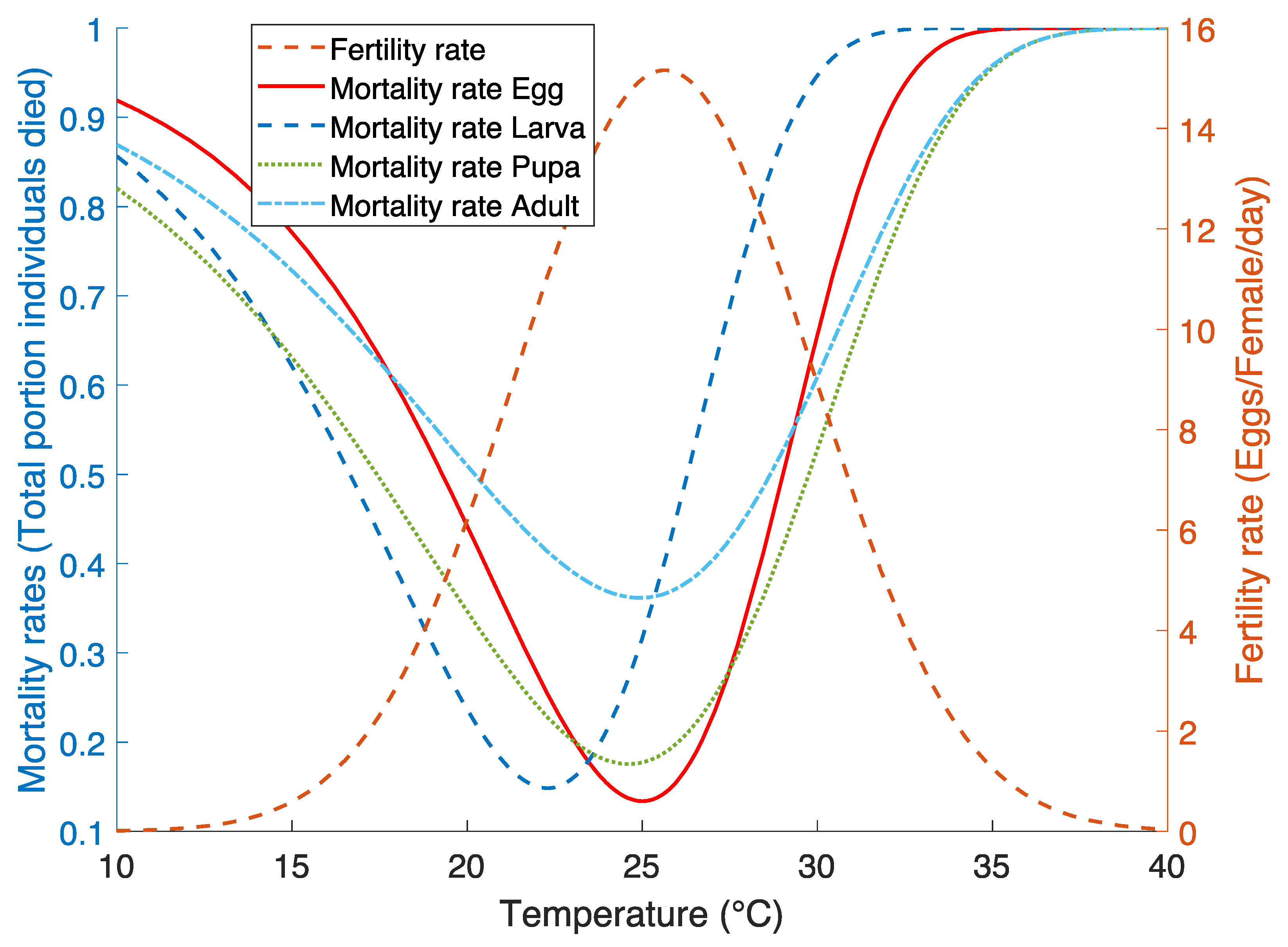
2.2.2. Mortality Rate Functions
2.2.3. Fertility Rate Functions
2.3. Field Data Collection for Model Validation
2.4. Model Evaluation and Comparison
3. Results
3.1. Development Rate Functions
3.2. Mortality Rate Functions
3.3. Fertility Rate Function
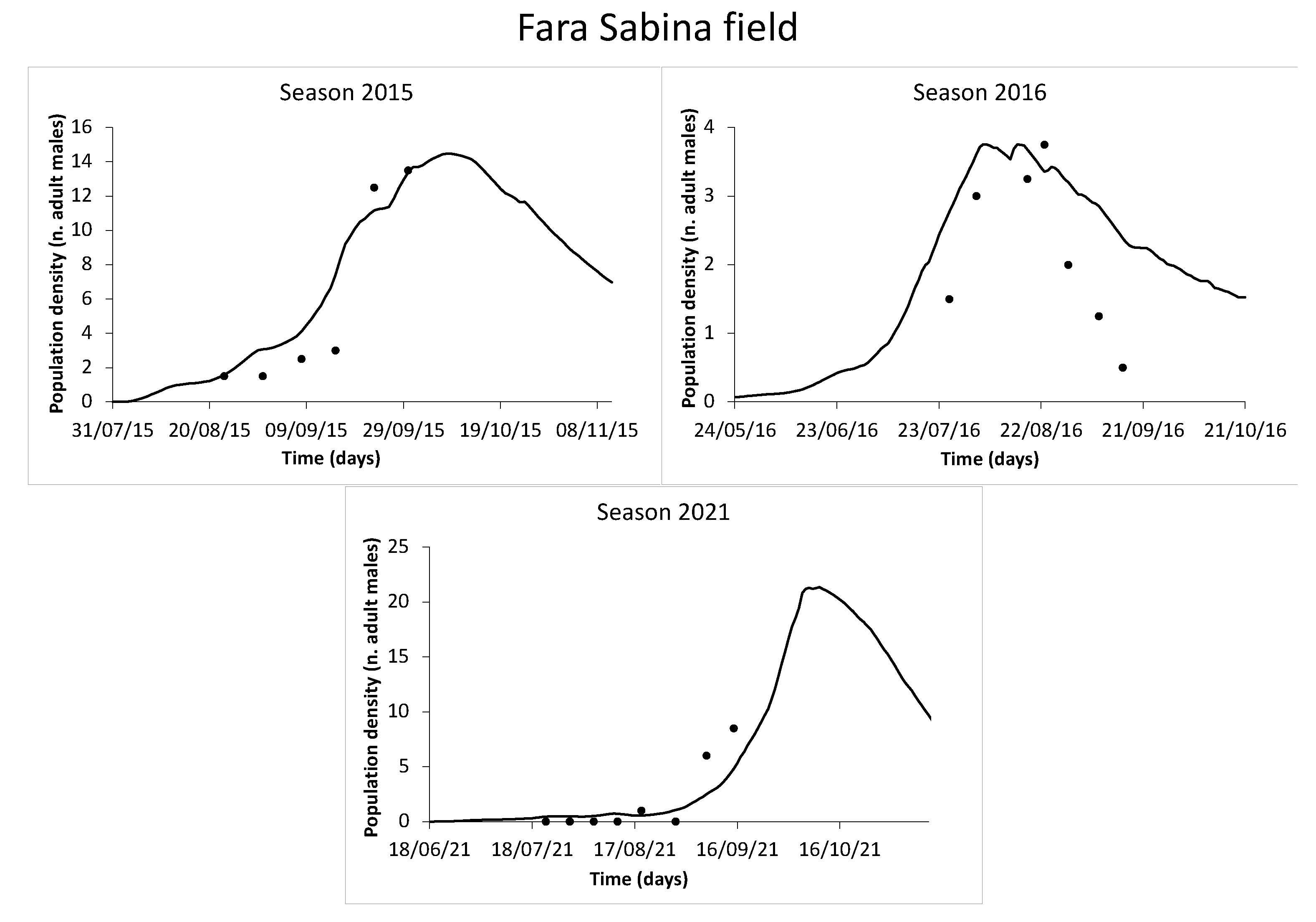
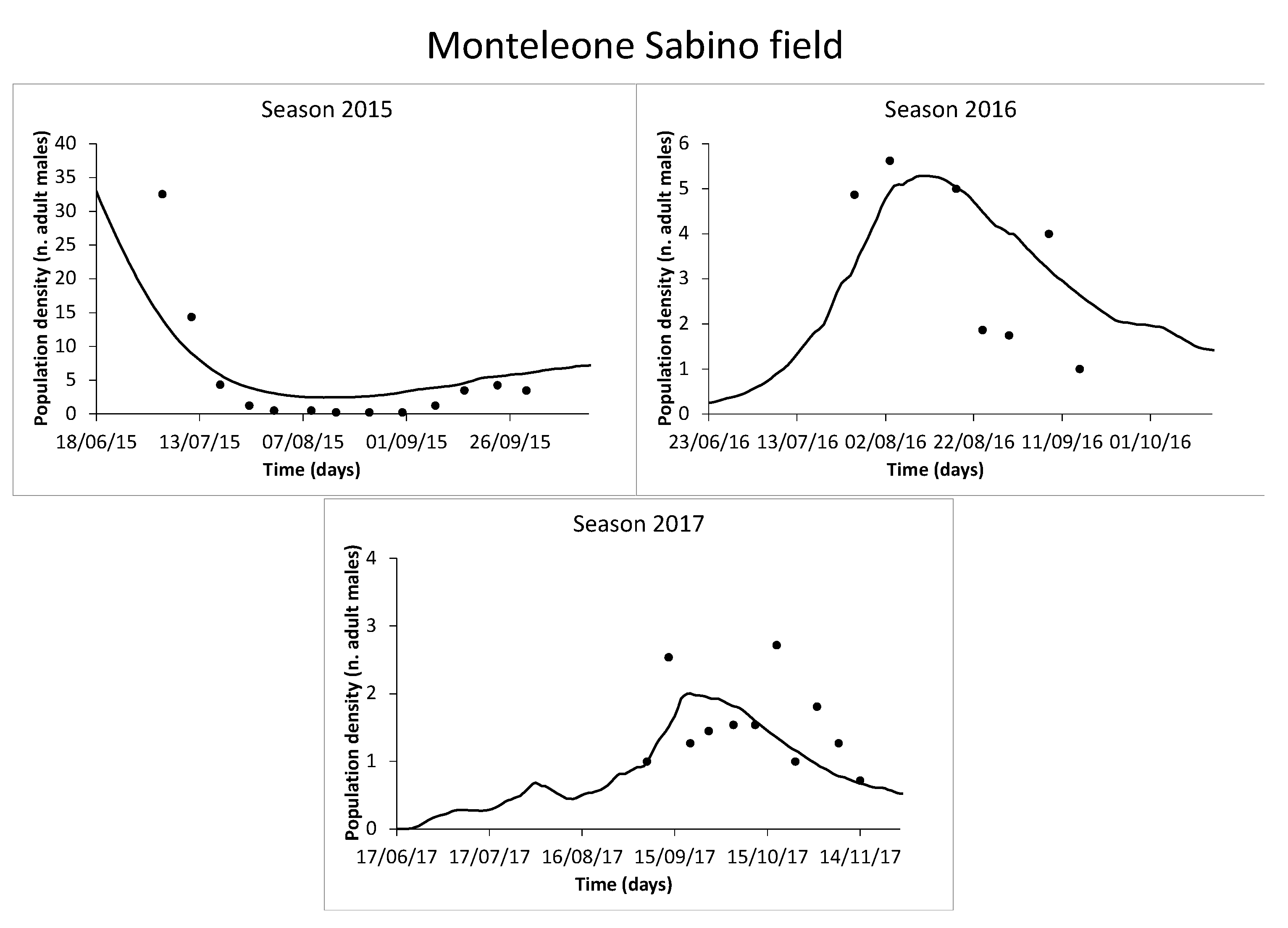
3.4. Model Validation
4. Discussion and Conclusions
4.1. Comparison with Other Existing Models
4.2. Development, Mortality, and Fertility Rates
4.3. Population Trend and Model Validation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutierrez, A.P.; Ponti, L.; Cossu, Q.A. Effects of climate warming on olive and olive fly (Bactrocera oleae (Gmelin)) in California and Italy. Clim. Change 2009, 95, 195–217. [Google Scholar] [CrossRef]
- Baratella, V.; Pucci, C.; Paparatti, B.; Speranza, S. Response of Bactrocera oleae to different photoperiods and temperatures using a novel method for continuous laboratory rearing. Biol. Control 2017, 110, 79–88. [Google Scholar] [CrossRef]
- Daane, K.M.; Johnson, M.W. Olive fruit fly: Managing an ancient pest in modern times. Annu. Rev. Entomol. 2010, 55, 151–169. [Google Scholar] [CrossRef]
- EPPO. EPPO Global Database. Available online: https://gd.eppo.int/taxon/ (accessed on 14 August 2022).
- Malheiro, R.; Casal, S.; Baptista, P.; Pereira, J.A. A review of Bactrocera oleae (Rossi) impact in olive products: From the tree to the table. Trends Food Sci. Technol. 2015, 44, 226–242. [Google Scholar] [CrossRef]
- Preu, M.; Friess, J.L.; Breckling, B.; Schröder, W. Case Study 1: Olive Fruit Fly (Bactrocera oleae); von Gleich, A., Schröder, W., Eds.; Springer International Publishing: Cham, Switzerland, 2020; ISBN 9783030389338. [Google Scholar]
- Pereira, J.A.; Alves, M.R.; Casal, S.; Oliveira, M.B.P.P. Effect of olive fruit fly infestation on the quality of olive oil from cultivars cobrançosa, madural and verdeal transmontana. Ital. J. Food Sci. 2004, 16, 355–365. [Google Scholar]
- Koprivnjak, O.; Dminić, I.; Kosić, U.; Majetić, V.; Godena, S.; Valenčič, V. Dynamics of oil quality parameters changes related to olive fruit fly attack. Eur. J. Lipid Sci. Technol. 2010, 112, 1033–1040. [Google Scholar] [CrossRef]
- Mraicha, F.; Ksantini, M.; Zouch, O.; Ayadi, M.; Sayadi, S.; Bouaziz, M. Effect of olive fruit fly infestation on the quality of olive oil from Chemlali cultivar during ripening. Food Chem. Toxicol. 2010, 48, 3235–3241. [Google Scholar] [CrossRef]
- Medjkouh, L.; Tamendjari, A.; Keciri, S.; Santos, J.; Nunes, M.A.; Oliveira, M.B.P.P. The effect of the olive fruit fly (Bactrocera oleae) on quality parameters, and antioxidant and antibacterial activities of olive oil. Food Funct. 2016, 7, 2780–2788. [Google Scholar] [CrossRef]
- Yokoyama, V.Y. Olive fruit fly (Diptera: Tephritidae) in California table olives, USA: Invasion, distribution, and management implications. J. Integr. Pest Manag. 2015, 6, 14. [Google Scholar] [CrossRef]
- Boccaccio, L.; Petacchi, R. Landscape effects on the complex of Bactrocera oleae parasitoids and implications for conservation biological control. BioControl 2009, 54, 607–616. [Google Scholar] [CrossRef]
- Pontikakos, C.M.; Tsiligiridis, T.A.; Drougka, M.E. Location-aware system for olive fruit fly spray control. Comput. Electron. Agric. 2010, 70, 355–368. [Google Scholar] [CrossRef]
- Voulgaris, S.; Stefanidakis, M.; Floros, A.; Avlonitis, M. Stochastic modeling and simulation of olive fruit fly outbreaks. Procedia Technol. 2013, 8, 580–586. [Google Scholar] [CrossRef]
- Kokkari, A.I.; Pliakou, O.D.; Floros, G.D.; Kouloussis, N.A.; Koveos, D.S. Effect of fruit volatiles and light intensity on the reproduction of Bactrocera (Dacus) oleae. J. Appl. Entomol. 2017, 141, 841–847. [Google Scholar] [CrossRef]
- Broumas, T.; Haniotakis, G.; Liaropoulos, C.; Tomazou, T.; Ragoussis, N. The efficacy of an improved form of the mass-trapping method, for the control of the olive fruit fly, Bactrocera oleae (Gmelin) (Dipt., Tephritidae): Pilot-scale feasibility studies. J. Appl. Entomol. 2002, 126, 217–223. [Google Scholar] [CrossRef]
- Tang, S.; Cheke, R.A. Models for integrated pest control and their biological implications. Math. Biosci. 2008, 215, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Kalamatianos, R.; Kermanidis, K.; Avlonitis, M.; Karydis, I. Environmental impact on predicting olive fruit fly population using trap measurements. In Artificial Intelligence Applications and Innovations, Proceedings of the 12th IFIP WG 12.5 International Conference and Workshops, AIAI 2016, Thessaloniki, Greece, 16–18 September 2016; Iliadis, L., Maglogiannis, I., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 475, pp. 180–190. [Google Scholar]
- Capinera, J.L. Handbook of Vegetable Pests; Capinera, J.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2001; ISBN 9780121588618. [Google Scholar]
- Rossini, L.; Severini, M.; Contarini, M.; Speranza, S. Use of ROOT to build a software optimized for parameter estimation and simulations with Distributed Delay Model. Ecol. Inform. 2019, 50, 184–190. [Google Scholar] [CrossRef]
- Kalamatianos, R.; Kermanidis, K.; Karydis, I.; Avlonitis, M. Treating stochasticity of olive-fruit fly’s outbreaks via machine learning algorithms. Neurocomputing 2018, 280, 135–146. [Google Scholar] [CrossRef]
- Pucci, C.; Spanedda, A.F. Performance comparison between two forecasting models of infestation caused by olive fruit fly (Bactrocera oleae Rossi). Pomol. Croat. 2006, 12, 193–202. [Google Scholar]
- Preti, M.; Verheggen, F.; Angeli, S. Insect pest monitoring with camera-equipped traps: Strengths and limitations. J. Pest Sci. 2021, 94, 203–217. [Google Scholar] [CrossRef]
- Shannon, C.E. Communication in the presence of noise. Proc. IRE 1949, 37, 10–21. [Google Scholar] [CrossRef]
- Pierpaoli, E.; Carli, G.; Pignatti, E.; Canavari, M. Drivers of precision agriculture technologies adoption: A literature review. Procedia Technol. 2013, 8, 61–69. [Google Scholar] [CrossRef]
- Capalbo, S.M.; Antle, J.M.; Seavert, C. Next generation data systems and knowledge products to support agricultural producers and science-based policy decision making. Agric. Syst. 2017, 155, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Rossini, L.; Rosselló, N.B.; Speranza, S.; Garone, E. A general ODE-based model to describe the physiological age structure of ectotherms: Description and application to Drosophila suzukii. Ecol. Model. 2021, 456, 109673. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef]
- Mirhosseini, M.A.; Fathipour, Y.; Reddy, G.V.P. Arthropod development’s response to temperature: A review and new software for modeling. Ann. Entomol. Soc. Am. 2017, 110, 507–520. [Google Scholar] [CrossRef]
- Schmalensee, L.; Gunnarsdóttir, K.H.; Näslund, J.; Gotthard, K.; Lehmann, P. Thermal performance under constant temperatures can accurately predict insect development times across naturally variable microclimates. Ecol. Lett. 2021, 24, 1633–1645. [Google Scholar] [CrossRef]
- Alilla, R.; Speranza, S.; Perovic, T.; Hrncic, S.; Pesolillo, S.; Pucci, C.; Severini, M. Modello a ritardo variabile per la simulazione della fenologia e della demografia della Bactrocera oleae (Gmel) (Diptera, Tephritidae) in due diversi ambienti olivicoli e in condizioni di aumento della temperatura. In Proceedings of the Quarte Giornate Studio su Metodi Numerici, Statistici e Informatici Nella Difesa Delle Colture Agrarie e Delle Foreste, Ricerca ed Applicazioni, Viterbo, Italy, 27–29 March 2007; pp. 48–50. [Google Scholar]
- Gilioli, G.; Pasquali, S. Use of individual-based models for population parameters estimation. Ecol. Model. 2007, 200, 109–118. [Google Scholar] [CrossRef]
- Petacchi, R.; Marchi, S.; Federici, S.; Ragaglini, G. Large-scale simulation of temperature-dependent phenology in wintering populations of Bactrocera oleae (Rossi). J. Appl. Entomol. 2015, 139, 496–509. [Google Scholar] [CrossRef]
- Rossini, L.; Rosselló, N.B.; Contarini, M.; Speranza, S.; Garone, E. Modelling ectotherms’ populations considering physiological age structure and spatial motion: A novel approach. Ecol. Inform. 2022, 70, 101703. [Google Scholar] [CrossRef]
- Wang, X.; Levy, K.; Son, Y.; Johnson, M.W.; Daane, K.M. Comparison of the thermal performance between a population of the olive fruit fly and its co-adapted parasitoids. Biol. Control 2012, 60, 247–254. [Google Scholar] [CrossRef]
- Rossini, L.; Contarini, M.; Giarruzzo, F.; Assennato, M.; Speranza, S. Modelling Drosophila suzukii adult male populations: A physiologically based approach with validation. Insects 2020, 11, 751. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Canale, A.; Bonsignori, G.; Ragni, G.; Stefanini, C.; Raspi, A. Male wing vibration in the mating behavior of the olive fruit fly Bactrocera oleae (Rossi) (Diptera: Tephritidae). J. Insect Behav. 2012, 25, 590–603. [Google Scholar] [CrossRef]
- Otero, M.; Solari, H.G.; Schweigmann, N. A stochastic population dynamics model for Aedes aegypti: Formulation and application to a city with temperate climate. Bull. Math. Biol. 2006, 68, 1945–1974. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Lewis, E.E. Modelling temperature-dependent development and survival of Otiorhynchus sulcatus (Coleoptera: Curculionidae). Agric. For. Entomol. 2005, 7, 201–209. [Google Scholar] [CrossRef]
- Shi, P.-J.; Reddy, G.V.P.; Chen, L.; Ge, F. Comparison of thermal performance equations in describing temperature-dependent developmental rates of insects: (I) Empirical models. Ann. Entomol. Soc. Am. 2017, 110, 113–120. [Google Scholar] [CrossRef]
- Pasquali, S.; Soresina, C.; Marchesini, E. Mortality estimate driven by population abundance field data in a stage-structured demographic model. The case of Lobesia botrana. Ecol. Model. 2022, 464, 109842. [Google Scholar] [CrossRef]
- Aguirre-Zapata, E.; Morales, H.; Dagatti, C.; di Sciascio, F.; Amicarelli, A.N. Semi physical growth model of Lobesia botrana under laboratory conditions for Argentina’s Cuyo Region. Ecol. Model. 2022, 464, 109803. [Google Scholar] [CrossRef]
- Aguirre, E.; Andreo, V.; Porcasi, X.; Lopez, L.; Guzman, C.; González, P.; Scavuzzo, C.M. Implementation of a proactive system to monitor Aedes aegypti populations using open access historical and forecasted meteorological data. Ecol. Inform. 2021, 64, 101351. [Google Scholar] [CrossRef]
- Severini, M.; Gilioli, G. Storia e filosofia dei modelli di simulazione nella difesa delle colture agrarie. Not. Sulla Prot. Delle Piante 2002, 15, 9–29. [Google Scholar]
- Damos, P.; Savopoulou-Soultani, M. Temperature-driven models for insect development and vital thermal requirements. Psyche 2012, 2012, 123405. [Google Scholar] [CrossRef]
- Quinn, B.K. A critical review of the use and performance of different function types for modeling temperature-dependent development of arthropod larvae. J. Therm. Biol. 2017, 63, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Ikemoto, T.; Kiritani, K. Novel method of specifying low and high threshold temperatures using thermodynamic SSI model of insect development. Environ. Entomol. 2019, 48, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Rossini, L.; Contarini, M.; Severini, M.; Talano, D.; Speranza, S. A modelling approach to describe the Anthonomus eugenii (Coleoptera: Curculionidae) life cycle in plant protection: A priori and a posteriori Analysis. Fla. Entomol. 2020, 103, 259–263. [Google Scholar] [CrossRef]
- Sharpe, P.J.H.; DeMichele, D.W. Reaction kinetics of poikilotherm development. J. Theor. Biol. 1977, 64, 649–670. [Google Scholar] [CrossRef]
- Wagner, T.L.; Wu, H.-I.; Sharpe, P.J.H.; Coulson, R.N. Modeling distributions of insect development time: A literature review and application of the Weibull function. Ann. Entomol. Soc. Am. 1984, 77, 475–483. [Google Scholar] [CrossRef]
- Schoolfield, R.M.; Sharpe, P.J.H.; Magnuson, C.E. Non-linear regression of biological temperature-dependent rate models based on absolute reaction-rate theory. J. Theor. Biol. 1981, 88, 719–731. [Google Scholar] [CrossRef]
- Rossini, L.; Severini, M.; Contarini, M.; Speranza, S. A novel modelling approach to describe an insect life cycle vis-à-vis plant protection: Description and application in the case study of Tuta absoluta. Ecol. Model. 2019, 409, 108778. [Google Scholar] [CrossRef]
- Rossini, L.; Speranza, S.; Severini, M.; Locatelli, D.P.; Limonta, L. Life tables and a physiologically based model application to Corcyra cephalonica (Stainton) populations. J. Stored Prod. Res. 2021, 91, 101781. [Google Scholar] [CrossRef]
- Logan, J.A.; Wollkind, D.J.; Hoyt, S.C.; Tanigoshi, L.K. An Analytic model for description of temperature dependent rate phenomena in arthropods. Environ. Entomol. 1976, 5, 1133–1140. [Google Scholar] [CrossRef]
- Briere, J.-F.; Pracros, P.; le Roux, A.-Y.; Pierre, J.-S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Rossini, L.; Severini, M.; Contarini, M.; Speranza, S. EntoSim, a ROOT-based simulator to forecast insects’ life cycle: Description and application in the case of Lobesia botrana. Crop Prot. 2020, 129, 105024. [Google Scholar] [CrossRef]
- Ponti, L.; Gutierrez, A.P.; de Campos, M.R.; Desneux, N.; Biondi, A.; Neteler, M. Biological invasion risk assessment of Tuta absoluta: Mechanistic versus correlative methods. Biol. Invasions 2021, 23, 3809–3829. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Marshall, K.E.; Sewell, M.A.; Levesque, D.L.; Willett, C.S.; Slotsbo, S.; Dong, Y.; Harley, C.D.G.; Marshall, D.J.; Helmuth, B.S.; et al. Can we predict ectotherm responses to climate change using thermal performance curves and body temperatures? Ecol. Lett. 2016, 19, 1372–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossini, L.; Virla, E.G.; Albarracín, E.L.; van Nieuwenhove, G.A.; Speranza, S. Evaluation of a physiologically based model to predict Dalbulus maidis occurrence in maize crops: Validation in two different subtropical areas of South America. Entomol. Exp. Appl. 2021, 169, 597–609. [Google Scholar] [CrossRef]
- Rossini, L.; Speranza, S.; Mazzaglia, A.; Turco, S. EntoSim, an insects life cycle simulator enclosing multiple models in a Docker container. Environ. Eng. Manag. J. 2021, 20, 1703–1710. [Google Scholar] [CrossRef]
- Kim, D.S.; Lee, J.H. Oviposition µodel of Carposina sasakii (Lepidoptera: Carposinidae). Ecol. Model. 2003, 162, 145–153. [Google Scholar] [CrossRef]
- Ryan, G.D.; Emiljanowicz, L.; Wilkinson, F.; Kornya, M.; Newman, J.A. Thermal tolerances of the spotted-wing drosophila Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2016, 109, 746–752. [Google Scholar] [CrossRef]
- Bellocchi, G.; Rivington, M.; Donatelli, M.; Matthews, K. Validation of biophysical models: Issues and methodologies. In Sustainable Agriculture; Springer: Dordrecht, The Netherlands, 2011; Volume 2, pp. 577–603. ISBN 9789048126651. [Google Scholar]
- Rossini, L.; Contarini, M.; Speranza, S. A Novel version of the Von Foerster equation to describe poikilothermic organisms including physiological age and reproduction rate. Ric. Mat. 2021, 70, 489–503. [Google Scholar] [CrossRef]
- Pappalardo, S.; Villa, M.; Santos, S.A.P.; Benhadi-Marín, J.; Pereira, J.A.; Venturino, E. A tritrophic interaction model for an olive tree pest, the olive moth—Prays oleae (Bernard). Ecol. Model. 2021, 462, 109776. [Google Scholar] [CrossRef]
- Brunetti, M.; Capasso, V.; Montagna, M.; Venturino, E. A Mathematical model for Xylella fastidiosa epidemics in the Mediterranean regions. Promoting good agronomic practices for their effective control. Ecol. Model. 2020, 432, 109204. [Google Scholar] [CrossRef]
- Pucci, C.; Iannotta, N.; Duro, N.; Jaupi, A.; Thomaj, F.; Speranza, S.; Paparatti, B. Application of a statistical forecast model on the olive fruit fly (Bactrocera oleae) infestation and oil analysis in Albania. Bull. Insectol. 2013, 66, 309–314. [Google Scholar]
- Caselli, A.; Petacchi, R. Climate change and major pests of Mediterranean olive orchards: Are we ready to face the global heating? Insects 2021, 12, 802. [Google Scholar] [CrossRef] [PubMed]
- Manetsch, T.J. Time-varying distributed delays and their use in aggregative models of large systems. IEEE Trans. Syst. Man Cybern. 1976, SMC-6, 547–553. [Google Scholar] [CrossRef]
- Vansickle, J. Attrition in distributed delay models. IEEE Trans. Syst. Man Cybern. 1977, 7, 635–638. [Google Scholar] [CrossRef]
- Bellagamba, V.; di Cola, G.; Cavalloro, R. Stochastic models in fruit-fly population dynamics. In Proceedings of the CEC/IOBC International Symposium “Fruit Flies of Economic Importance 87”, Rome, Italy, 7–10 April 1987; pp. 91–98. [Google Scholar]
- Pappas, M.L.; Broufas, G.D.; Koufali, N.; Pieri, P.; Koveos, D.S. Effect of heat stress on survival and reproduction of the olive fruit fly Bactocera (Dacus) oleae. J. Appl. Entomol. 2011, 135, 359–366. [Google Scholar] [CrossRef]
- Wang, X.-G.; Johnson, M.W.; Daane, K.M.; Nadel, H. High summer temperatures affect the survival and reproduction of olive fruit fly (Diptera: Tephritidae). Environ. Entomol. 2009, 38, 1496–1504. [Google Scholar] [CrossRef]
- Mansour, A.A.; Kahime, K.; Chemseddine, M.; Boumezzough, A. Study of the population dynamics of the olive fly, Bactrocera oleae Rossi (Diptera, Tephritidae) in the region of Essaouira. Open J. Ecol. 2015, 5, 174–186. [Google Scholar] [CrossRef]
- Marchi, S.; Guidotti, D.; Ricciolini, M.; Petacchi, R. Towards understanding temporal and spatial dynamics of Bactrocera oleae (Rossi) infestations using decade-long agrometeorological time series. Int. J. Biometeorol. 2016, 60, 1681–1694. [Google Scholar] [CrossRef]
- Ponti, L.; Cossu, A.; Gutierrez, A.P. Climate warming effects on the Olea europaea—Bactrocera oleae system in Mediterranean islands: Sardinia as an example. Glob. Chang. Biol. 2009, 15, 2874–2884. [Google Scholar] [CrossRef]
- Ordano, M.; Engelhard, I.; Rempoulakis, P.; Nemny-Lavy, E.; Blum, M.; Yasin, S.; Lensky, I.M.; Papadopoulos, N.T.; Nestel, D. Olive fruit fly (Bactrocera oleae) population dynamics in the eastern Mediterranean: Influence of exogenous uncertainty on a monophagous frugivorous insect. PLoS ONE 2015, 10, e0127798. [Google Scholar] [CrossRef]
- Doebeli, M.; de Jong, G. Genetic variability in sensitivity to population density affects the dynamics of simple ecological models. Theor. Popul. Biol. 1999, 55, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Severini, M.; Baumgärtner, J.; Ricci, M. Theory and practice of parameter estimation of distributed delay models for insect and plant phenologies. In Meteorology and Environmental Sciences; World Scientific: Singapore, 1990; pp. 674–719. [Google Scholar]
- Di Cola, G.; Gilioli, G.; Baumgärtner, J. Mathematical Models for Age-structured population dynamics: An overview. In Proceedings of the 20th International Congress of Entomology, Florence, Italy, 25–31 August 1996; pp. 45–61. [Google Scholar]
- Baumgärtner, J.; Gutierrez, A.P.; Pesolillo, S.; Severini, M. A model for the overwintering process of European grapevine moth Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Tortricidae) populations. J. Entomol. Acarol. Res. 2012, 44, 2. [Google Scholar] [CrossRef]
- Nance, J.; Fryxell, R.T.; Lenhart, S. Modeling a single season of Aedes albopictus populations based on host-seeking data in response to temperature and precipitation in eastern Tennessee. J. Vector Ecol. 2018, 43, 138–147. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; You, M.; Atlıhan, R.; Smith, C.L.; Kavousi, A.; Özgökçe, M.S.; Güncan, A.; Tuan, S.-J.; Fu, J.-W.; Xu, Y.-Y.; et al. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Broufas, G.D.; Pappas, M.L.; Koveos, D.S. Effect of relative humidity on longevity, ovarian maturation, and egg production in the olive fruit fly (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2009, 102, 70–75. [Google Scholar] [CrossRef]
- Podgornik, M.; Vuk, I.; Arbeiter, A.; Hladnik, M.; Bandelj, D. Population fluctuation of adult males of the olive fruit fly Bactrocera oleae (Rossi) analysis in olive orchards in relation to abiotic factors. Entomol. News 2013, 123, 15–25. [Google Scholar] [CrossRef]
- Noce, M.E.; Perri, E.; Scalercio, S.; Iannotta, N. Phenolic compounds and susceptibility of olive cultivar to Bactrocera oleae (Diptera: Tephritidae) infestations and complementary aspects: A Review. Acta Hortic. 2014, 1057, 177–183. [Google Scholar] [CrossRef]
- Grasso, F.; Coppola, M.; Carbone, F.; Baldoni, L.; Alagna, F.; Perrotta, G.; Pérez-Pulido, A.J.; Garonna, A.; Facella, P.; Daddiego, L.; et al. The transcriptional response to the olive fruit fly (Bactrocera oleae) reveals extended differences between tolerant and susceptible olive (Olea europaea L.) varieties. PLoS ONE 2017, 12, e0183050. [Google Scholar] [CrossRef]
- Bjeliš, M.; Masten, T.; Mladen, M. Olive fruit infestation by olive fruit fly Bactrocera oleae Gmel. in dry and irrigated growing conditions in Dalmacija. In Proceedings of the VII Alps-Adria Scientific Workshop, Stara Lesna, Slovakia, 28 April–2 May 2008; Volume 36. [Google Scholar]
- González-Zamora, J.E.; Alonso-López, M.T.; Gómez-Regife, Y.; Ruiz-Muñoz, S. Decreased water use in a super-intensive olive orchard mediates arthropod populations and pest damage. Agronomy 2021, 11, 1337. [Google Scholar] [CrossRef]
- Bono Rossello, N.; Rossini, L.; Speranza, S.; Garone, E. State estimation of pest populations subject to intermittent measurements. In Proceedings of the Sensing, Control and Automation Technologies for Agriculture—7th AGRICONTROL 2022, Munich, Germany, 14–16 September 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Development Rate Function | Best Fit Parameters (±SE) | Goodness of Fit | |
|---|---|---|---|
| Sharpe and De Michele (4) | |||
| Logan (5) | |||
| Briére (6) | |||
| Life Stage | Best Fit Parameters (±SE) | Goodness of Fit | |
|---|---|---|---|
| Egg | |||
| Larva | |||
| Pupa | |||
| Adult | |||
| Best Fit Parameters (±SE) | Goodness of Fit | |
|---|---|---|
| Area | Year of Survey | Initial Conditions * | Time Range of Simulations | Model Evaluation Parameters |
|---|---|---|---|---|
| Fara Sabina | 2015 | 17 August–30 September | ||
| 2016 | 19 July–15 September | |||
| 2021 | 15 July–15 September | |||
| Monteleone Sabino | 2015 | 27 June–30 September | ||
| 2016 | 19 July–15 September | |||
| 2017 | 30 August–14 November | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossini, L.; Bruzzone, O.A.; Contarini, M.; Bufacchi, L.; Speranza, S. A Physiologically Based ODE Model for an Old Pest: Modeling Life Cycle and Population Dynamics of Bactrocera oleae (Rossi). Agronomy 2022, 12, 2298. https://doi.org/10.3390/agronomy12102298
Rossini L, Bruzzone OA, Contarini M, Bufacchi L, Speranza S. A Physiologically Based ODE Model for an Old Pest: Modeling Life Cycle and Population Dynamics of Bactrocera oleae (Rossi). Agronomy. 2022; 12(10):2298. https://doi.org/10.3390/agronomy12102298
Chicago/Turabian StyleRossini, Luca, Octavio Augusto Bruzzone, Mario Contarini, Livio Bufacchi, and Stefano Speranza. 2022. "A Physiologically Based ODE Model for an Old Pest: Modeling Life Cycle and Population Dynamics of Bactrocera oleae (Rossi)" Agronomy 12, no. 10: 2298. https://doi.org/10.3390/agronomy12102298