
Changes in Soil Properties, Bacterial Communities and Wheat Roots Responding to Subsoiling in South Loess Plateau of China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Design
2.2. Collection of Wheat Root and Soil Samples
2.3. Determination of Soil Physical and Chemical Properties
2.4. Extraction and PCR Amplification of Soil Microbial Total DNA
2.5. Purifying PCR Product and High-Throughput Sequencing Analysis
2.6. Sequencing Data Processing
2.7. Statistical Analysis
3. Results
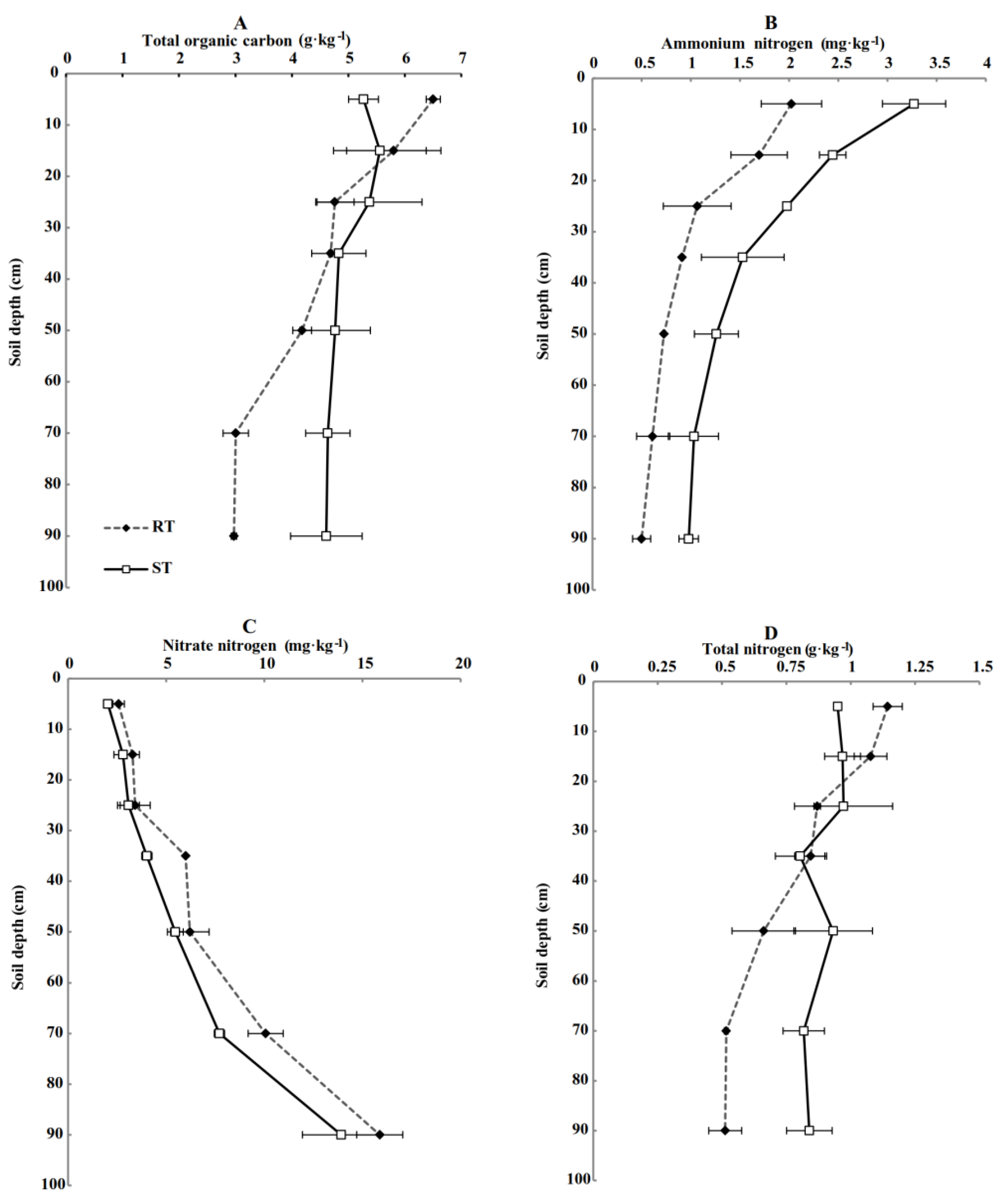
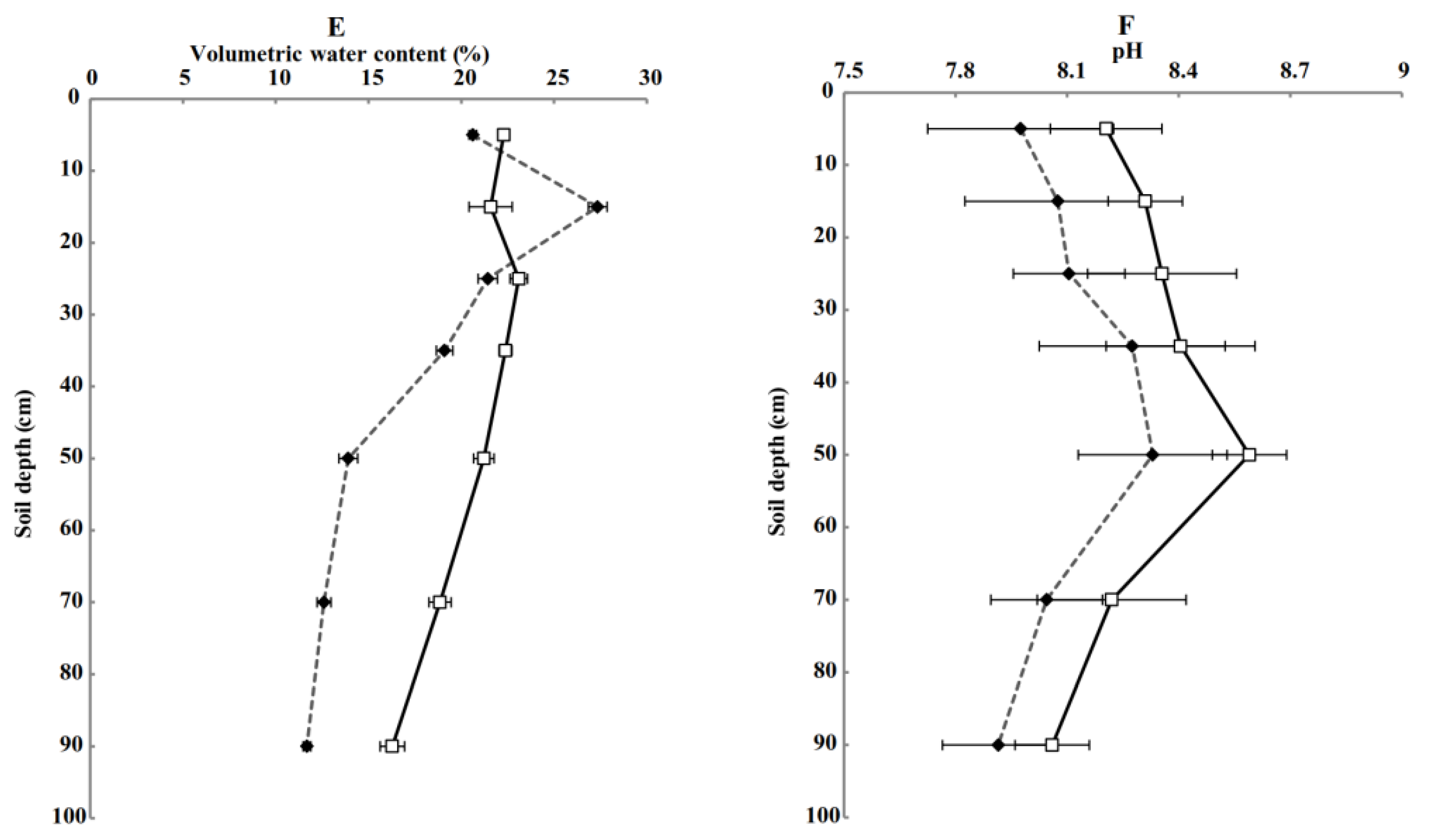
3.1. Soil Physical and Chemical Properties of Different Treated Soil Profiles (0–100 cm)
3.2. Wheat Root Length Density in Soil Profile (0–100 cm) under Different Treatments
3.3. Bacterial Community Composition in Surface Layer (0–20 cm) of Different Treatments
3.4. Soil Bacterial Diversity Index of Surface Layer (0–20 cm) under Different Treatments
3.5. Relationship between Bacterial Community Structure in Surface Layer (0–20 cm) of Different Treatments and Soil Physical and Chemical Properties and Root Length Density
4. Discussion
4.1. Effects of Subsoiling + Rotary Tillage on Physical and Chemical Properties and Root Length Density of Soil Profile (0–100 cm)
4.2. Effects of Subsoiling + Rotary Tillage on Bacterial Diversity in Topsoil (0–20 cm)
4.3. Differences in Bacterial Community Composition and Structure in Surface Layer under Different Tillage Treatments and Their Relationship with Physical and Chemical Properties and Root Length Density
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.L.; Gao, P.C.; Tong, Y.N.; Norse, D.; Lu, Y.L.; Powlson, D. Overcoming nitrogen fertilizer over-use through technical and advisory approaches: A case study from Shaanxi Province, northwest China. Agric. Ecosyst. Environ. 2015, 209, 89–99. [Google Scholar] [CrossRef]
- Wang, S.B.; Guo, L.L.; Zhou, P.C.; Wang, X.J.; Shen, Y.; Han, H.F.; Nang, T.Y.; Han, K. Effect of subsoiling depth on soil physical properties and summer maize (Zea mays L.) yield. Plant Soil Environ. 2019, 65, 131–137. [Google Scholar] [CrossRef]
- Moraes, M.T.D.; Debiasi, H.Q.; Franchini, J.C.; Mastroberti, A.A.; Levien, R.; Leitner, D.; Schnepf, A. Soil compaction impacts soybean root growth in an Oxisol from subtropical Brazil. Soil Tillage Res. 2020, 200, 104611. [Google Scholar] [CrossRef]
- Wiesmeier, M.; Hubner, R.; Barthold, F.; Sporlein, P.; Geub, U.; Hangen, E.; Reischl, A.; Schilling, B.; Lutzow, M.V.; Knabner, L.K. Amount, distribution and driving factors of soil organic carbon and nitrogen in cropland and grassland soils of southeast Germany (Bavaria). Agric. Ecosyst. Environ. 2013, 176, 39–52. [Google Scholar] [CrossRef]
- Han, E.; Dresboll, D.B.; Kristensen, K.T. Tracing deep P uptake potential in arable subsoil using radioactive 33P isotope. Plant Soil 2022, 472, 91–104. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Lilley, J.M.; Howe, G.N.; Graham, M. Impact of subsoil water use on wheat yield. Aust. J. Agric. Res. 2007, 58, 303–315. [Google Scholar] [CrossRef]
- Hussein, M.A.; Muche, H.; Schmitter, P.; Nakawuka, P.; Tilahun, S.A.; Langan, S.; Barron, J.; Steenhuis, T.S. Deep tillage improves degraded soils in the (sub)humid ethiopian highlands. Land 2019, 8, 159. [Google Scholar] [CrossRef]
- Schneider, F.; Don, A.; Hennings, I.; Schmittmann, O.; Seidel, S.J. The effect of deep tillage on crop yield—What do we really know? Soil Tillage Res. 2017, 174, 193–204. [Google Scholar] [CrossRef]
- Gourley, C.J.P.; Sale, P.W.G. Chemical and physical amelioration of subsoils has limited production benefits for perennial pastures in two contrasting soils. Soil Tillage Res. 2014, 144, 41–52. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, P.; Sodhi, G.P.S. Soil organic carbon and biological indicators of uncultivated vis-à-vis intensively cultivated soils under rice–wheat and cotton–wheat cropping systems in South-Western Punjab. Carbon Manag. 2020, 11, 681–695. [Google Scholar] [CrossRef]
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Sharma, S.; Khare, P.K.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers. Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A.; et al. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Abbadie, L.; Clays-Josserand, A.; Degrange, V.; Grayston, S.J.; Guillaumaud, N.; Loiseau, P.; Louault, F.; Mahmood, S.; Nazaret, S.; et al. Effects of management regime and plant species on the enzyme activity and genetic structure of N-fixing, denitrifying and nitrifying bacterial communities in grassland soils. Environ. Microbiol. 2006, 8, 1005–1016. [Google Scholar] [CrossRef]
- Yang, Q.Y.; Norton, J.M.; Stark, J.M.; Reeve, J.R.; Habteselassie, M.Y. Ammonia-oxidizing bacteria are more responsive than archaea to nitrogen source in an agricultural soil. Soil Biol. Biochem. 2016, 96, 4–15. [Google Scholar] [CrossRef]
- Chaurasia, A.; Meena, B.R.; Tripathi, A.N.; Pandey, K.K.; Rai, A.B.; Singh, B. Actinomycetes: An unexplored microorganism for plant growth promotion and biocontrol in vegetable crops. World J. Microbiol. Biotechnol. 2018, 34, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Asmiaty, S.; Ambo, A.; Baharuddin, P.; Elkawakib, S. Effect of seed inoculation with Actinomycetes and Rhizobium isolated from indigenous soybean and rhizosphere on nitrogen fixation, growth, and yield of soybean. Int. J. Agron. 2018, 2018, 4371623. [Google Scholar] [CrossRef]
- Laudicina, V.A.; Palazzolo, E.; Catania, P.; Vallone, M.; Garcia, A.G.; Badalucco, L. Soil quality indicators as affected by shallow tillage in a vineyard grown in a semiarid Mediterranean environment. Land Degrad. Dev. 2017, 28, 1038–1046. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Fu, G.Z.; Zhao, Y.G. Rotary tillage in rotation with plowing tillage improves soil properties and crop yield in a wheat-maize cropping system. PLoS ONE 2018, 13, e0198193. [Google Scholar] [CrossRef]
- Heinze, S.; Rauber, R.; Joergensen, R.G. Influence of mouldboard plough and rotary harrow tillage on microbial biomass and nutrient stocks in two long-term experiments on loess derived Luvisols. Appl. Soil Ecol. 2010, 46, 405–412. [Google Scholar] [CrossRef]
- Moreira, B.C.; Júnior, P.P.; Dell, B.; Kasuya, M.C.M. Roots and Beneficial Interactions with Soil Microbes. In Subsoil Constraints for Crop Production; Oliveira, T.S.D., Bell, R.W., Eds.; Springer: Cham, Switzerland, 2022; pp. 263–287. [Google Scholar] [CrossRef]
- Six, J.; Feller, C.; Denef, K.; Ogle, S.; Sa, J.C.D.M.; Albrecht, A. Soil organic matter, biota and aggregation in temperate and tropical soils—Effects of no-tillage. Agron. EDP Sci. 2002, 22, 755. [Google Scholar] [CrossRef]
- Li, Y.Y.; Zhai, Z.; Cong, P.; Zhang, Y.T.; Pang, H.C.; Dong, G.H.; Gao, J.S. Effect of plough pan thickness on crop growth parameters, nitrogen uptake and greenhouse gas (CO2 and N2O) emissions in a wheat-maize double-crop rotation in the Northern China Plain: A one-year study. Agric. Water Manag. 2019, 213, 534–545. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.K.; Guo, Z.; Li, J.B.; Tian, C.; Hua, D.W.; Shi, C.D.; Wang, H.Y.; Han, J.C.; Xu, Y. Effects of conservation tillage on soil physicochemical properties and crop yield in an arid Loess Plateau, China. Sci. Rep. 2020, 10, 4716. [Google Scholar] [CrossRef] [PubMed]
- Izumi, Y.; Yoshida, T.; Iijima, M. Effects of subsoiling to the non-tilled field of wheat-soybean rotation on the root system development, water uptake, and yield. Plant Prod. Sci. 2009, 12, 327–335. [Google Scholar] [CrossRef]
- Yin, J.D.; Zhang, X.C.; Ma, Y.F.; Yu, X.F.; Hou, H.Z.; Wang, H.L.; Fang, Y.J. Vertical rotary sub-soiling under ridge–furrow with plastic mulching system increased crops yield by efficient use of deep soil moisture and rainfall. Agric. Water Manag. 2022, 271, 107767. [Google Scholar] [CrossRef]
- Xia, Q.; Liu, X.L.; Gao, Z.Q.; Wang, J.M.; Yang, Z.P. Responses of rhizosphere soil bacteria to 2-year tillage rotation treatments during fallow period in semiarid southeastern Loess Plateau. PeerJ 2020, 8, e8853. [Google Scholar] [CrossRef]
- Ji, B.Y.; Hu, H.; Zhao, Y.L.; Mu, X.Y.; Liu, K.; Li, C.H. Effects of deep tillage and straw returning on soil microorganism and enzyme activities. Sci. World J. 2014, 2014, 451493. [Google Scholar] [CrossRef]
- Hu, R.W.; Liu, Y.J.; Chen, T.; Zheng, Z.Y.; Peng, G.J.; Zou, Y.D.; Tang, C.G.; Shan, X.H.; Zhou, Q.M.; Li, J. Responses of soil aggregates, organic carbon, and crop yield to short-term intermittent deep tillage in Southern China. J. Clean. Prod. 2021, 298, 126767. [Google Scholar] [CrossRef]
- Song, K.; Zheng, X.Q.; Lv, W.Q.; Qin, Q.; Sun, L.J.; Zhang, H.L.; Xue, Y. Effects of tillage and straw return on water-stable aggregates, carbon stabilization and crop yield in an estuarine alluvial soil. Sci. Rep. 2019, 9, 4586. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Q.Q.; Hu, J.L.; Wei, H.Y.; Zhang, H.C.; Zhu, J.Y. Relationship between plant roots, rhizosphere microorganisms, and nitrogen and its special focus on rice. Agriculture 2021, 11, 234. [Google Scholar] [CrossRef]
- Preusser, S.; Liebmann, P.; Stucke, A.; Wirsching, J.; Muller, K.; Mikutta, R.; Guggenberger, G.; Don, A.; Kalbitz, K.; Bachmann, J.; et al. Microbial utilisation of aboveground litter-derived organic carbon within a sandy dystric cambisol profile. Front. Soil Sci. 2021, 1, 666950. [Google Scholar] [CrossRef]
- Shokralla, S.; Spall, J.; Gibson, J.F.; Hajibabaei, M. Next-generation sequencing technologies for environmental DNA research. Mol. Ecol. 2012, 21, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Kaurin, A.; Mihelic, R.; Kastelec, D.; Grcman, H.; Bru, D.; Philippot, L.; Suhadolc, M. Resilience of bacteria, archaea, fungi and N-cycling microbial guilds under plough and conservation tillage, to agricultural drought. Soil Biol. Biochem. 2018, 120, 233–245. [Google Scholar] [CrossRef]
- Sun, R.B.; Li, W.Y.; Dong, W.X.; Tian, Y.P.; Hu, C.S.; Liu, B.B. Tillage changes vertical distribution of soil bacterial and fungal communities. Front. Microbiol. 2018, 9, 699. [Google Scholar] [CrossRef]
- Beretta, A.N.; Silbermann, A.V.; Paladino, L.; Torres, D.; Kassahun, D.; Musselli, R.; Lamohte, A.G. Soil texture analyses using a hydrometer: Modification of the Bouyoucos method. Cienc. Investig. Agrar. 2014, 41, 263–271. Available online: https://dialnet.unirioja.es/descarga/articulo/5316082.pdf (accessed on 6 February 2020). [CrossRef]
- Ahmed, F.; Arthur, E.; Liu, H.; Andersen, M.N. New rootsnap sensor reveals the ameliorating effect of biochar on in situ root growth dynamics of maize in sandy soil. Front. Plant Sci. 2020, 11, 949. [Google Scholar] [CrossRef]
- Mantoura, R.F.C.; Woodward, E.M.S. Optimization of the indophenol blue method for the automated determination of ammonia in estuarine waters. Estuar. Coast. Shelf Sci. 1983, 17, 219–224. [Google Scholar] [CrossRef]
- Norman, R.J.; Edberg, J.C.; Stucki, J.W. Determination of nitrate in soil extracts by dual-wavelength ultraviolet spectrophotometry. Soil Sci. Soc. Am. J. 1985, 49, 1182–1185. [Google Scholar] [CrossRef]
- Kuczynskin, J.; Lauber, C.L.; Walters, W.A.; Parfrey, L.W.; Clemente, J.C.; Gevers, D.; Knight, R. Experimental and analytical tools for studying the human microbiome. Nat. Rev. Genet. 2011, 13, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Peiffer, J.A.; Spor, A.; Koren, O.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Weber, C.F. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Devis, J.; Young, A.C.; Program, N.C.S.; Bouffard, G.G.; Blakesley, R.W.; Blakesley, R.W.; et al. Topographical and temporal diversity of the human skin microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Xue, P.P.; Carrillo, Y.; Pino, V.; Minasny, B.; Mcbratney, A.B. Soil properties drive microbial community structure in a large scale transect in south eastern Australia. Sci. Rep. 2018, 8, 11725. [Google Scholar] [CrossRef] [Green Version]
- Eilers, K.G.; Debenport, S.; Anderson, S.; Fierer, N. Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. Biochem. 2012, 50, 58–65. [Google Scholar] [CrossRef]
- Li, W.J.; Jiang, L.M.; Zhang, Y.; Teng, D.X.; Wang, H.F.; Wang, J.L.; Lv, G.H. Structure and driving factors of the soil microbial community associated with Alhagi sparsifolia in an arid desert. PLoS ONE 2021, 16, e0254065. [Google Scholar] [CrossRef] [PubMed]
- Roesch, L.F.W.; Fulthorpe, R.R.; Riva, A.; Casella, G.; Hadwin, A.K.M.; Kent, A.D.; Daroub, S.H.; Camargo, F.A.O.; Farmerie, W.G.; Triplett, E.W. Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J 2007, 1, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, Z.M.; Liu, Y.H.; Wang, F.; Liu, Y.R.; Zhao, J.T.; Li, Y.F.; Gao, Y.X.; Zhu, N.L. Roles of plant-associated microorganisms in regulating the fate of Hg in croplands: A perspective on potential pathways in maintaining sustainable agriculture. Sci. Total Environ. 2022, 834, 155204. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lan, L.Y.; Jin, Y.; Yu, N.; Wang, D.; Wang, E. Mechanisms underlying legume-rhizobium symbioses. J. Integr. Plant Biol. 2021, 64, 244–267. [Google Scholar] [CrossRef]
- Przemieniecki, W.S.; Kurowski, P.T.; Karwowska, A. Plant growth promoting potential of Pseudomonas sp. SP0113 isolated from potable water from a closed water well. Arch. Biol. Sci. 2015, 67, 663–673. [Google Scholar] [CrossRef]
- George, P.; Gupta, A.; Gopal, M.; Thomas, L.; Thomas, G.V. Multifarious beneficial traits and plant growth promoting potential of Serratia marcescens KiSII and Enterobacter sp. RNF 267 isolated from the rhizosphere of coconut palms (Cocos nucifera L.). World J. Microbiol. Biotechnol. 2013, 29, 109–117. [Google Scholar] [CrossRef]
- Yao, Q.; Peng, D.C. Nitrite oxidizing bacteria (NOB) dominating in nitrifying community in full-scale biological nutrient removal wastewater treatment plants. AMB Express 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.X.; Suh, J.W. Effects of Actinobacteria on plant disease suppression and growth promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef]
- Bollmann, A.; Sedlacek, C.J.; Norton, J.; Laanbroek, H.J.; Suwa, Y.; Stein, L.Y.; Klotz, M.G.; Arp, D.; Sayavedra-Soto, L.; Lu, M.; et al. Complete genome sequence of Nitrosomonas sp. Is79, an ammonia oxidizing bacterium adapted to low ammonium concentrations. Stand Genom. Sci. 2013, 7, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil Acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition-current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Paśmionka, I.B.; Bulski, K.; Boligłowa, E. The participation of microbiota in the transformation of nitrogen compounds in the soil—A review. Agronomy 2021, 11, 977. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Ayangbenro, A.S.; Glick, B.R.; Babalola, O.O. Plant health: Feedback effect of root exudates-rhizobiome interactions. Appl. Microbiol. Biotechnol. 2019, 103, 1155–1166. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Zhao, S.; Mao, Y.; Zhang, J.; Pan, X.; He, F.; van der Ploeg, M. Impact of long-term sub-soiling tillage on soil porosity and soil physical properties in the soil profile. Land Degrad. Dev. 2021, 32, 2892–2905. [Google Scholar] [CrossRef]
- He, D.; Lu, C.; Tong, Z.; Zhong, G.; Ma, X. Research progress of minimal tillage method and machine in China. AgriEngineering 2021, 3, 633–647. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Types | Rotary Tillage (0–10 cm) RTa | Subsoiling and Rotary Tillage (0–10 cm) STa | Rotary Tillage (10–20 cm) RTb | Subsoiling and Rotary Tillage (10–20 cm) STb |
|---|---|---|---|---|
| o_Sphingomonadales | 5.9130 ± 0.1532 b | 6.9066 ± 0.1526 a | 3.8256 ± 0.2437 c | 2.8571 ± 0.1459 d |
| o_Xanthomonadales | 3.9411 ± 0.3192 b | 7.5155 ± 0.1482 a | 4.5856 ± 0.2921 b | 4.4502 ± 0.2271 b |
| o_Rhodospirillales | 3.6096 ± 0.0760 b | 2.6035 ± 0.1552 c | 8.9473 ± 0.5699 a | 3.7447 ± 0.1912 b |
| o_Rhizobiales | 2.5410 ± 0.1592 b | 3.0216 ± 0.1693 a | 1.9061 ± 0.1214 c | 2.5232 ± 0.1289 b |
| o_Nitrosomonadales | 3.3418 ± 0.2391 a | 2.6010 ± 0.1526 b | 3.7302 ± 0.2376 a | 3.2833 ± 0.1676 a |
| o_Flavobacteriales | 0.2149 ± 0.0109 bc | 7.9082 ± 0.1510 a | 0.3880 ± 0.0247 b | 0.1166 ± 0.0062 c |
| o_Nitrospirales | 2.0583 ± 0.2735 b | 1.8522 ± 0.1974 b | 1.8945 ± 0.1207 b | 5.4689 ± 0.2791 a |
| o_Burkholderiales | 1.0182 ± 0.0135 b | 2.3415 ± 0.0295 a | 0.7225 ± 0.0460 c | 2.2937 ± 0.1172 a |
| o_Gaiellales | 1.4707 ± 0.0541 d | 2.0693 ± 0.0516 c | 2.8266 ± 0.1801 b | 6.0688 ± 0.3097 a |
| g_Bacillus | 0.2384 ± 0.0108 a | 0.0305 ± 0.0022 c | 0.1953 ± 0.0047 b | 0.0146 ± 0.0004 c |
| g_Pseudomonas | 0.0672 ± 0.0021 b | 0.2544 ± 0.0091 a | 0.0294 ± 0.0038 c | 0.0789 ± 0.0021 b |
| g_Serratia | 0.0445 ± 0.0027 b | 0.3189 ± 0.0128 a | N | N |
| Soil Depth (cm) | Sample | Operational Taxonomic Units (OTUs) | Chao1 Index | Shannon Index | Simpson Index | Coverage |
|---|---|---|---|---|---|---|
| 0–10 | RTa | 3632 ± 36 a | 4022.20 ± 39.53 a | 6.5965 ± 0.0794 a | 0.0042 ± 0.0001 c | 0.9953 ± 0.0009 a |
| STa | 3664 ± 38 a | 4062.84 ± 42.60 a | 6.5772 ± 0.0382 a | 0.0044 ± 0.0002 c | 0.9951 ± 0.0008 a | |
| 10–20 | RTb | 2934 ± 115 b | 3164.35 ± 124.68 b | 6.3918 ± 0.0573 b | 0.0051 ± 0.0002 b | 0.9965 ± 0.0011 a |
| STb | 2126 ± 54 c | 2387.48 ± 61.72 c | 6.0779 ± 0.0436 c | 0.0059 ± 0.0002 a | 0.9958 ± 0.0010 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Zhang, D.; He, J.; Wang, L.; Ren, J.; Zhang, S.; Bai, W.; Song, J.; Lv, G.; Li, J. Changes in Soil Properties, Bacterial Communities and Wheat Roots Responding to Subsoiling in South Loess Plateau of China. Agronomy 2022, 12, 2288. https://doi.org/10.3390/agronomy12102288
Wang H, Zhang D, He J, Wang L, Ren J, Zhang S, Bai W, Song J, Lv G, Li J. Changes in Soil Properties, Bacterial Communities and Wheat Roots Responding to Subsoiling in South Loess Plateau of China. Agronomy. 2022; 12(10):2288. https://doi.org/10.3390/agronomy12102288
Chicago/Turabian StyleWang, Hanbo, Dasheng Zhang, Jiuxing He, Lijuan Wang, Jiameng Ren, Shuantang Zhang, Wenbo Bai, Jiqing Song, Guohua Lv, and Jiusheng Li. 2022. "Changes in Soil Properties, Bacterial Communities and Wheat Roots Responding to Subsoiling in South Loess Plateau of China" Agronomy 12, no. 10: 2288. https://doi.org/10.3390/agronomy12102288