
Salinity Effect on Plant Growth Parameters and Fruit Bioactive Compounds of Two Strawberry Cultivars, Coupled with Environmental Conditions Monitoring
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Physical Characteristics Determination
2.3. Evaluation of Plant Growth Parameters and Toxicity Symptoms
2.4. Total Soluble Solids (TSS), Total Titratable Acidity (TA) and pH Determinations
2.5. Total Phenol Content, Total Anthocyanin Content and Antioxidant Capacity Determination
2.6. Carbohydrates Determination
2.7. Organic Acids Determination
2.8. Statistical Analyses
3. Results
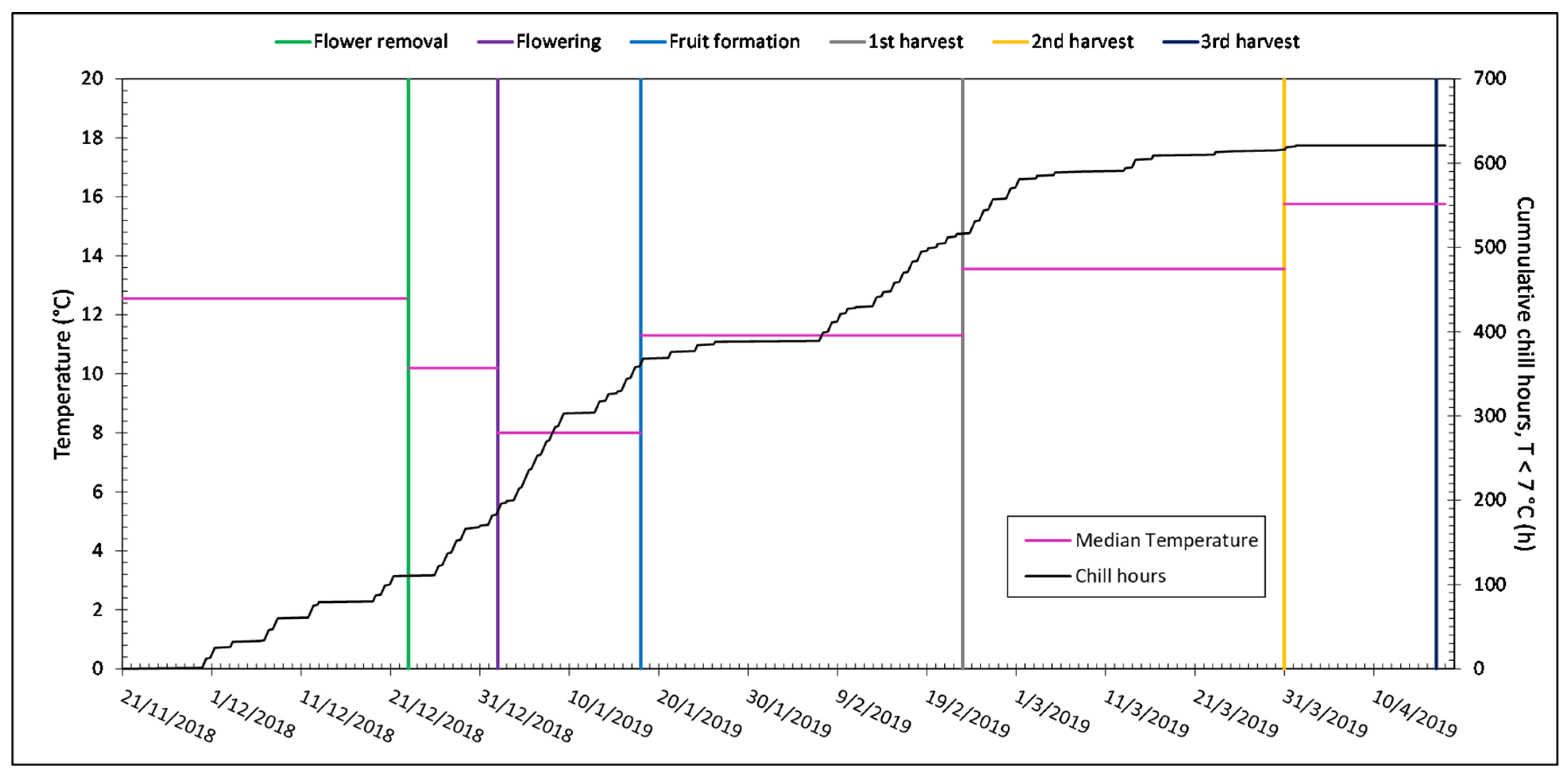
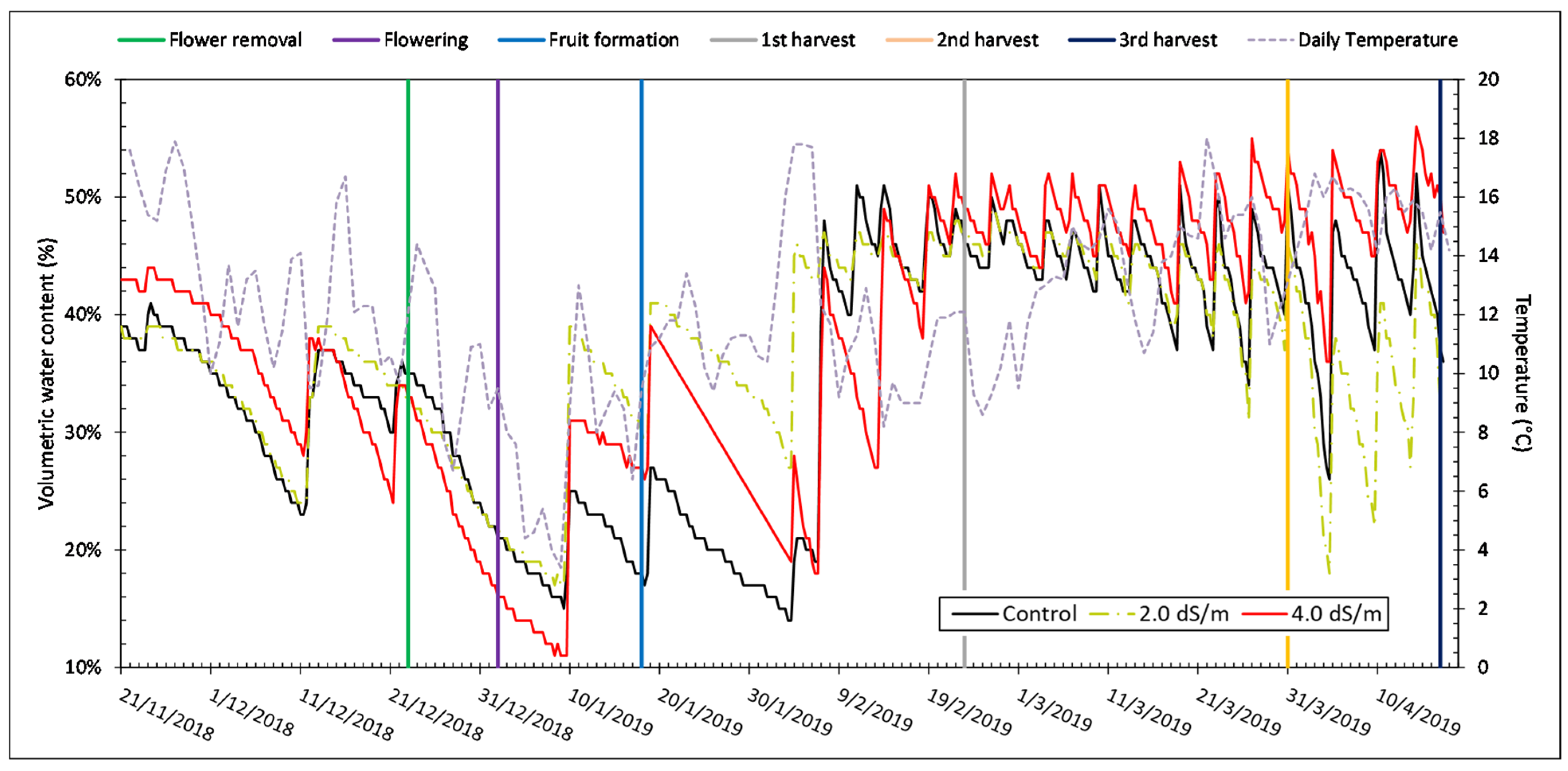
3.1. Microclimate and Soil Moisture Data
3.2. Effect of Salt Stress on Strawberry Fruit Physical and Quality Parameters
3.3. Effect of Salt Stress on the Phenolic and Anthocyanin Content of Strawberry Fruits and Antioxidant Capacity
3.4. Effect of Salt Stress on Strawberry Fruit Carbohydrate and Organic Acids Concentration
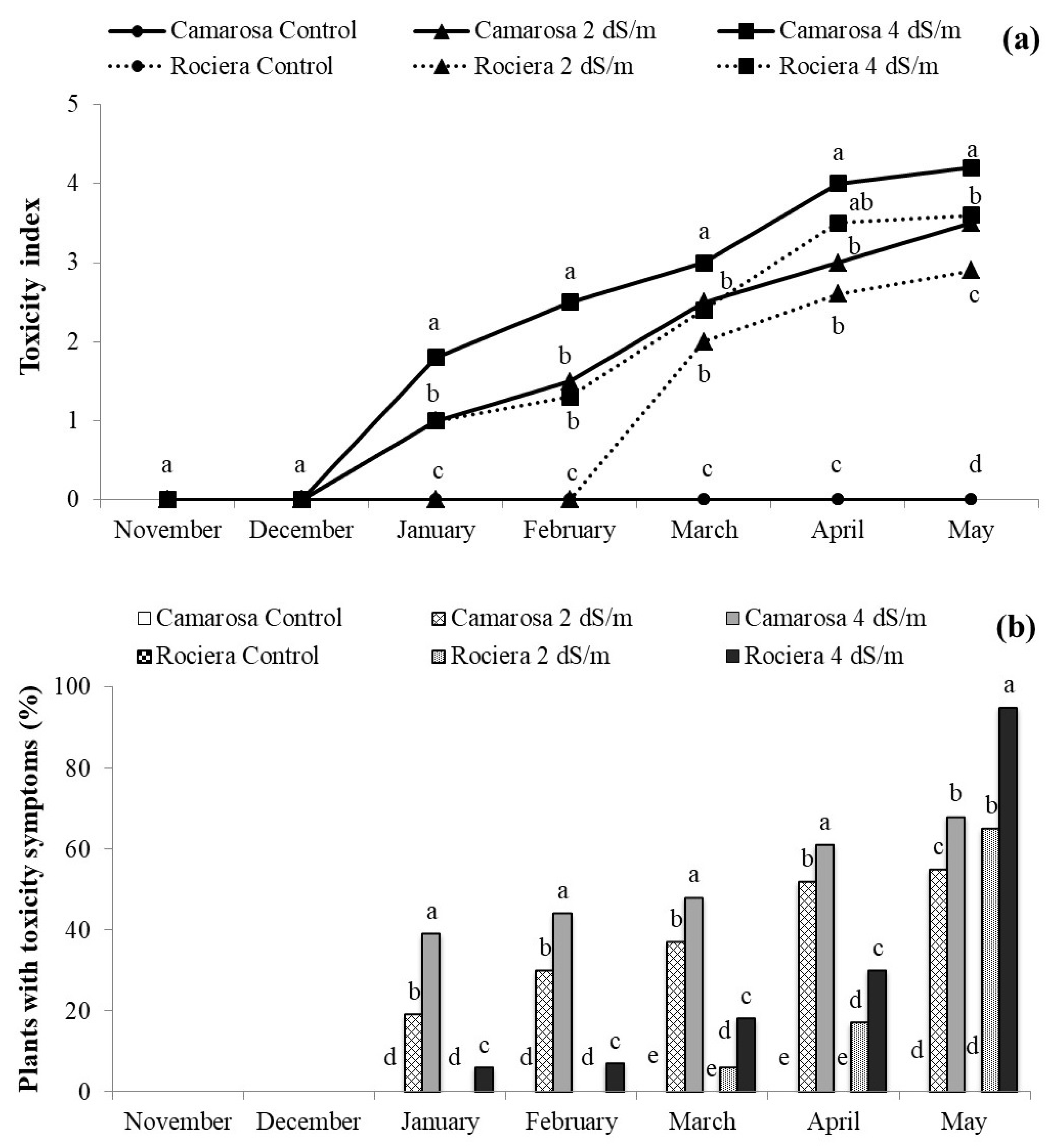
3.5. Effect of Salt Stress on Strawberry Growth Parameters and Toxicity Symptoms
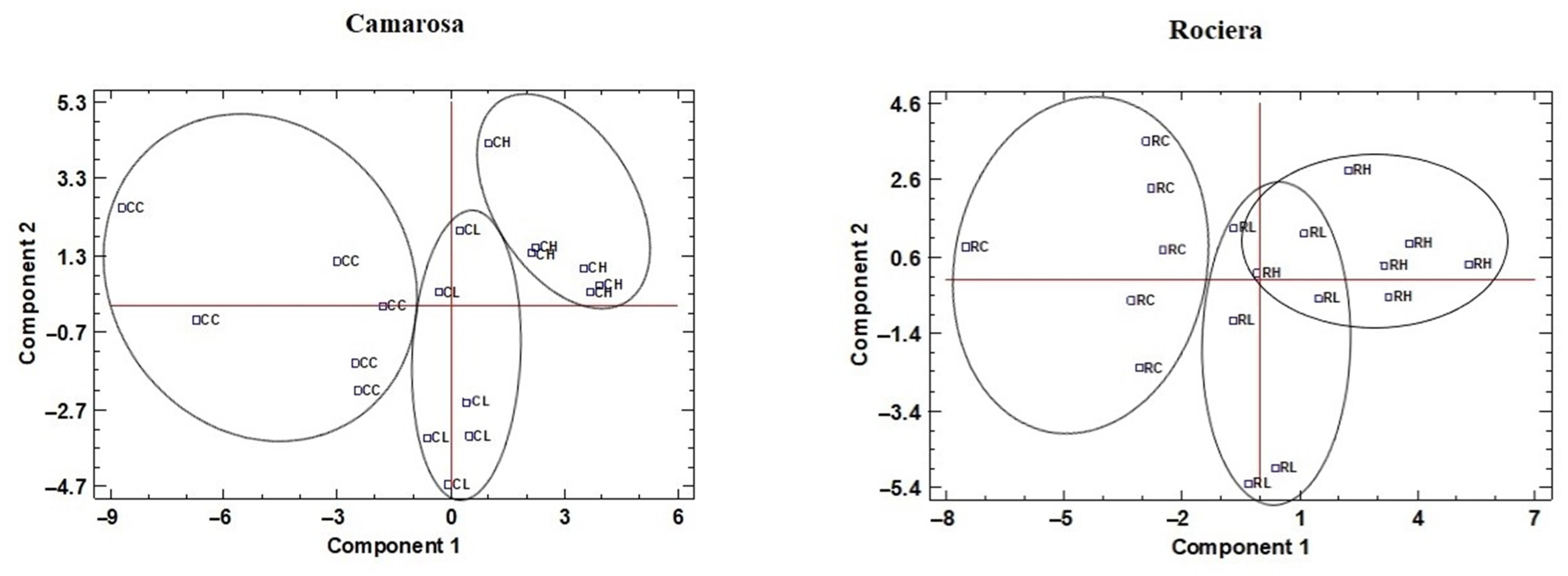
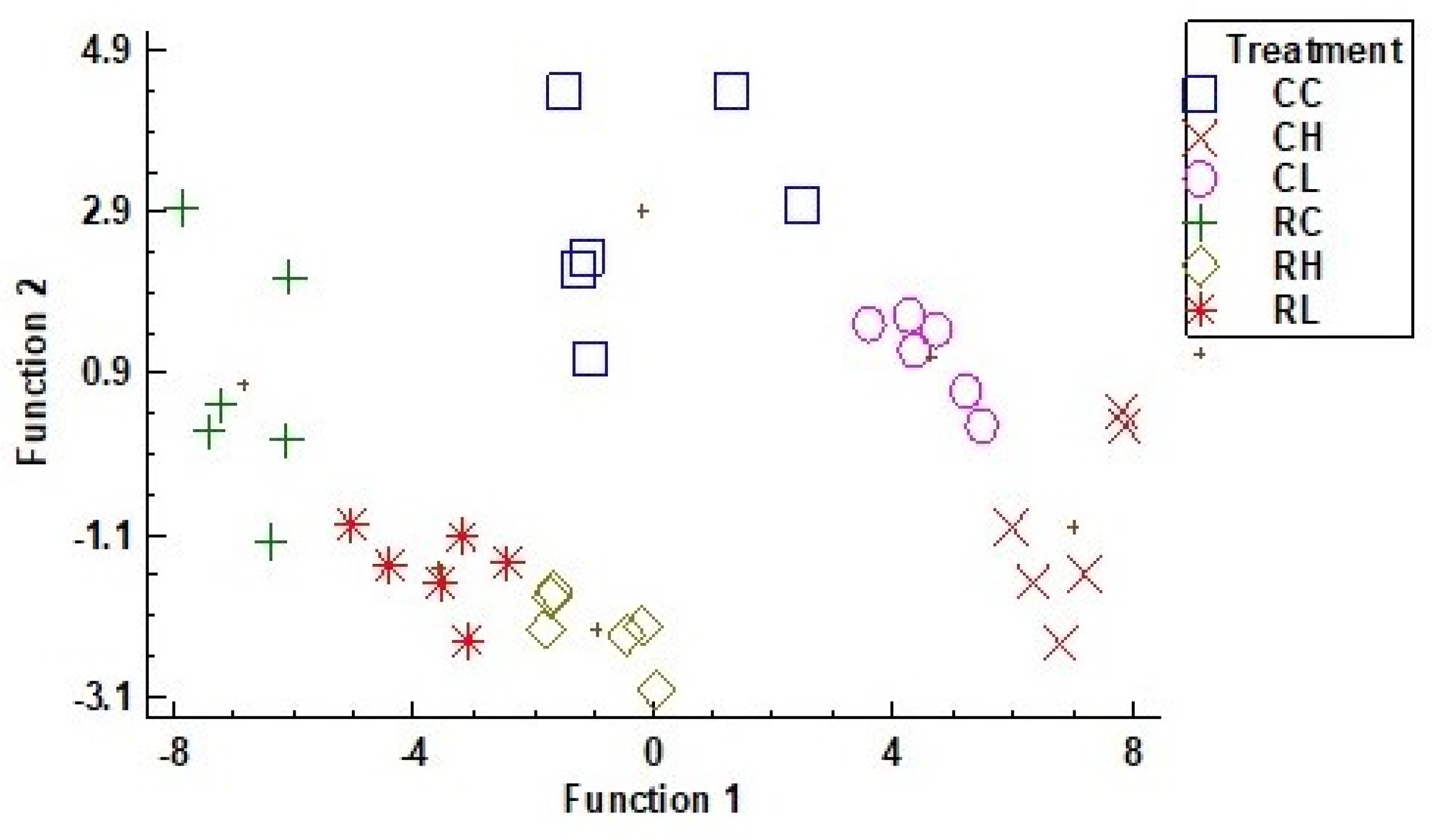
3.6. Principal Components Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, A.; Fasiha Amjad, S.; Hamzah Saleem, M.; Yasmin, H.; Imran, M.; Riaz, M.; Ali, Q.; Ahmad Joyia, F.; Mobeen; Ahmed, S.; et al. Foliar application of ascorbic acid enhances salinity stress tolerance in barley (Hordeum vulgare L.) through modulation of morpho-physio-biochemical attributes, ions uptake, osmo-protectants and stress response genes expression. Saudi J. Biol. Sci. 2021, 28, 4276–4290. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Saleem, M.H.; Ali, S.; Shahid, M.; Sagir, M.; Tahir, M.B.; Qureshi, K.A. Mitigation of salinity stress in barley genotypes with variable salt tolerance by application of zinc oxide nanoparticles. Front. Plant Sci. 2022, 13, 973782. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.G.; Silberbush, M.; Lips, S.H. Physiological studies on salinity and nitrogen interaction in alfalfa. i. biomass production and root development. J. Plant Nutr. 1994, 17, 657–668. [Google Scholar] [CrossRef]
- Quesada, V.; Ponce, M.R.; Micol, J.L. Genetic analysis of salt-tolerant mutants in Arabidopsis thaliana. Genetics 2000, 154, 421–436. [Google Scholar] [CrossRef]
- Blumwald, E.; Grober, A. Salt tolerance. In Plant Biotechnology: Current and Future Uses of Genetically Modified Crops; Halfor, N.G., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2006; pp. 206–224. [Google Scholar]
- Szabolcs, I. Soil and salinization. In Handbook of Plant and Crop Stress; Pessarakli, M., Ed.; Marcel Dekker: New York, NY, USA, 1994; pp. 3–11. [Google Scholar]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef]
- Al-Shorafa, W.; Mahadeen, A.; Al-Absi, K. Evaluation for salt stress tolerance in two strawberry cultivars. Am. J. Agric. Biol. Sci. 2014, 9, 334–341. [Google Scholar] [CrossRef]
- Kerbab, S.; Silini, A.; Bouket, A.C.; Cherif-Silini, H.; Eshelli, M.; Rabhi, N.E.H.; Belbahri, L. Mitigation of NaCl stress in wheat by rhizosphere engineering using salt habitat adapted PGPR halotolerant bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Magallon, K.J.; Dinneny, J.R. Environmental Stress: Salinity Ruins a Plant’s Day in the Sun. Curr. Biol. 2019, 29, R360–R362. [Google Scholar] [CrossRef]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [Green Version]
- Manaf, H.H. Beneficial effects of exogenous selenium, glycine betaine and seaweed extract on salt stressed cowpea plant. Ann. Agric. Sci. 2016, 61, 41–48. [Google Scholar] [CrossRef]
- Hu, L.; Hu, T.; Zhang, X.; Pang, H.; Fu, J. Exogenous glycine betaine Ameliorates the adverse effect of salt stress on perennial ryegrass. J. Am. Soc. Hortic. Sci. 2012, 137, 38–46. [Google Scholar] [CrossRef]
- Sofy, M.R.; Elhawat, N.; Alshaal, T. Glycine betaine counters salinity stress by maintaining high K+/Na+ ratio and antioxidant defense via limiting Na+ uptake in common bean (Phaseolus vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200, 110732. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Garriga, M.; Muñoz, C.A.; Caligari, P.D.S.; Retamales, J.B. Effect of salt stress on genotypes of commercial (Fragaria x ananassa) and Chilean strawberry (F. chiloensis). Sci. Hortic. 2015, 195, 37–47. [Google Scholar] [CrossRef]
- Jamali, B.; Eshghi, S. Salicylic acid-induced salinity redressal in hydroponically grown strawberry. Commun. Soil Sci. Plant Anal. 2015, 46, 1482–1493. [Google Scholar] [CrossRef]
- Ghaderi, N.; Hatami, M.; Mozafari, A.; Siosehmardeh, A. Change in antioxidant enzymes activity and some morpho-physiological characteristics of strawberry under long-term salt stress. Physiol. Mol. Biol. Plants 2018, 24, 833–843. [Google Scholar] [CrossRef]
- Karlidag, H.; Yildirim, E.; Turan, M. Role of 24-epibrassinolide in mitigating the adverse effects of salt stress on stomatal conductance, membrane permeability, and leaf water content, ionic composition in salt stressed strawberry (Fragaria × ananassa). Sci. Hortic. 2011, 130, 133–140. [Google Scholar] [CrossRef]
- Cogo, S.L.P.; Chaves, F.C.; Schirmer, M.A.; Zambiazi, R.C.; Nora, L.; Silva, J.A.; Rombaldi, C.V. Low soil water content during growth contributes to preservation of green colour and bioactive compounds of cold-stored broccoli (Brassica oleraceae L.) florets. Postharvest Biol. Technol. 2011, 60, 158–163. [Google Scholar] [CrossRef]
- Ripoll, J.; Urban, L.; Staudt, M.; Lopez-Lauri, F.; Bidel, L.P.R.; Bertin, N. Water shortage and quality of fleshy fruits-making the most of the unavoidable. J. Exp. Bot. 2014, 65, 4097–4117. [Google Scholar] [CrossRef] [Green Version]
- Awang, Y.B.; Atherton, J.G.; Taylor, A.J. Salinity effects on strawberry plants grown in rockwool. I. Growth and leaf water relations. J. Hortic. Sci. Biotechnol. 1993, 68, 783–790. [Google Scholar] [CrossRef]
- Sato, S.; Sakaguchi, S.; Furukawa, H.; Ikeda, H. Effects of NaCl application to hydroponic nutrient solution on fruit characteristics of tomato (Lycopersicon esculentum Mill.). Sci. Hortic. 2006, 109, 248–253. [Google Scholar] [CrossRef]
- Galli, V.; da Silva Messias, R.; Perin, E.C.; Borowski, J.M.; Bamberg, A.L.; Rombaldi, C.V. Mild salt stress improves strawberry fruit quality. LWT Food Sci. Technol. 2016, 73, 693–699. [Google Scholar] [CrossRef]
- Yildirim, E.; Karlidag, H.; Turan, M. Mitigation of salt stress in strawberry by foliar K, Ca and Mg nutrient supply. Plant Soil Environ. 2009, 55, 213–221. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef]
- Kepenek, K.; Koyuncu, F. Studies on the salt tolerance of some strawberry cultivars under glasshouse. Acta Hortic. 2002, 573, 297–304. [Google Scholar] [CrossRef]
- Saied, A.S.; Keutgen, A.J.; Noga, G. The influence of NaCl salinity on growth, yield and fruit quality of strawberry cvs. “Elsanta” and “Korona”. Sci. Hortic. 2005, 103, 289–303. [Google Scholar] [CrossRef]
- Gulen, H.; Turhan, E.; Eris, A. Changes in peroxidase activities and soluble proteins in strawberry varieties under salt-stress. Acta Physiol. Plant. 2006, 28, 109–116. [Google Scholar] [CrossRef]
- Turhan, E.; Eriş, A. Growth and stomatal behaviour of two stawberry cultivars under long-term salinity stress. Turkish J. Agric. For. 2007, 31, 55–61. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Quality and nutritional value of strawberry fruit under long term salt stress. Food Chem. 2008, 107, 1413–1420. [Google Scholar] [CrossRef]
- Orsini, F.; Alnayef, M.; Bona, S.; Maggio, A.; Gianquinto, G. Low stomatal density and reduced transpiration facilitate strawberry adaptation to salinity. Environ. Exp. Bot. 2012, 81, 1–10. [Google Scholar] [CrossRef]
- Sun, Y.; Niu, G.; Wallace, R.; Masabni, J.; Gu, M. Relative salt tolerance of seven strawberry cultivars. Horticulturae 2015, 1, 27–43. [Google Scholar] [CrossRef]
- Roussos, P.A.; Denaxa, N.K.; Ntanos, E.; Tsafouros, A.; Mavrikou, S.; Kintzios, S. Organoleptic, nutritional and anti-carcinogenic characteristics of the fruit and rooting performance of cuttings of black mulberry (Morus nigra L.) genotypes. J. Berry Res. 2020, 10, 77–93. [Google Scholar] [CrossRef]
- Denaxa, N.K.; Vemmos, S.N.; Roussos, P.A. The role of endogenous carbohydrates and seasonal variation in rooting ability of cuttings of an easy and a hard to root olive cultivars (Olea europaea L.). Sci. Hortic. 2012, 143, 19–28. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Modifications of strawberry fruit antioxidant pools and fruit quality under NaCl stress. J. Agric. Food Chem. 2007, 55, 4066–4072. [Google Scholar] [CrossRef]
- Turhan, E.; Eriş, A. Changes of Growth, Amino Acids, and Ionic Composition in Strawberry Plants under Salt Stress Conditions. Commun. Soil Sci. Plant Anal. 2009, 40, 3308–3322. [Google Scholar] [CrossRef]
- Pirlak, L.; Eşitken, A. Salinity effects on growth, proline and ion accumulation in strawberry plants. Acta Agric. Scand. Sect. B Soil Plant Sci. 2004, 54, 189–192. [Google Scholar] [CrossRef]
- Khayyat, M.; Tafazoli, E.; Rajaee, S.; Vazifeshenas, M.; Mahmoodabadi, M.R.; Sajjadinia, A. Effects of NaCl and Supplementary Potassium on Gas Exchange, Ionic Content, and Growth of Salt-Stressed Strawberry Plants. J. Plant Nutr. 2009, 32, 907–918. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Pawelzik, E. Impacts of NaCl stress on plant growth and mineral nutrient assimilation in two cultivars of strawberry. Environ. Exp. Bot. 2009, 65, 170–176. [Google Scholar] [CrossRef]
- Yilmaz, H.; Kina, A. The influence of NaCl salinity on some vegetative and chemical changes of strawberries (Fragaria x ananassa L.). Afr. J. Biotechnol. 2008, 7, 3299–3305. [Google Scholar]
- Ntanos, E.; Kekelis, P.; Assimakopoulou, A.; Gasparatos, D.; Denaxa, N.K.; Tsafouros, A.; Roussos, P.A. Amelioration effects against salinity stress in strawberry by bentonite–zeolite mixture, glycine betaine, and bacillus amy-loliquefaciens in terms of plant growth, nutrient content, soil properties, yield, and fruit quality characteristics. Appl. Sci. 2021, 11, 8796. [Google Scholar] [CrossRef]
- Yoon, J.; Hamayun, M.; Lee, S.; Lee, I. Methyl jasmonate alleviated salinity stress in soybean. J. Crop Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Kusvuran, S.; Ellialtioglu, S.; Yasar, F.; Abak, K. Effects of salt stress on ion accumulation and activity of some antioxidant enzymes in melon (Cucumis melo L.). J. Food Agric. Environ. 2007, 5, 351–354. [Google Scholar]
- Talei, D.; Kadir, M.A.; Yusop, M.K.; Valdiani, A.; Abdullah, M.P. Growth indices and salinity tolerance threshold in a medicinal plant Andrographis paniculata Nees. J. Med. Plants Res. 2013, 7, 104–110. [Google Scholar] [CrossRef]
- Haghshenas, M.; Javad Nazarideljou, M.; Shokoohian, A. Phytochemical and Quality Attributes of Strawberry Fruit under Osmotic Stress of Nutrient Solution and Foliar Application of Putrescine and Salicylic Acid Introduction. Int. J. Hortic. Sci. Technol. 2020, 7, 263–278. [Google Scholar] [CrossRef]
- Warrence, B.N.J.; Bauder, J.W.; Pearson, K.E. Basics of Salinity and Sodicity Effects on Soil Physical Properties; Montana State University-Bozeman: Bozeman, MT, USA, 2002. [Google Scholar]
- Garriga, M.; Caligari, P.; Retamales, J. Salt stress induces differential responses in yield, fruit quality, vegetative growth and physiological parameters in commercial (Fragaria × ananassa) and Chilean strawberry (F. chiloensis) genotypes. Acta Hortic. 2017, 1156, 419–424. [Google Scholar] [CrossRef]
- Kaya, C.; Kirnak, H.; Higgs, D.; Saltali, K. Supplementary calcium enhances plant growth and fruit yield in strawberry cultivars grown at high (NaCL) salinity. Sci. Hortic. 2002, 93, 65–74. [Google Scholar] [CrossRef]
- Khayyat, M.; Tafazoli, E.; Eshghi, S.; Rahemi, M.; Rajaee, S. Salinity, Supplementary Calcium and Potassium Effects on Fruit Yield and Quality of Strawberry (Fragaria ananassa Duch.). Am. Eurasian J. Agric. Environ. Sci. 2007, 2, 539–544. [Google Scholar]
- Ondrašek, G.; Romić, D.; Romić, M.; Duralija, B.; Mustač, I. Strawberry growth and fruit yield in a saline environment. Agric. Conspec. Sci. 2006, 71, 155–158. [Google Scholar]
- Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Fruit yield and survival of five commercial strawberry cultivars under field cultivation and salinity stress. Sci. Hortic. 2019, 243, 401–410. [Google Scholar] [CrossRef]
- Bybordi, A.; Saadat, S.; Zargaripour, P. The effect of zeolite, selenium and silicon on qualitative and quantitative traits of onion grown under salinity conditions. Arch. Agron. Soil Sci. 2018, 64, 520–530. [Google Scholar] [CrossRef]
- Awang, Y.B.; Atherton, J.G.; Taylor, A.J. Salinity effects on strawberry plants grown in rockwool. II. Fruit quality. J. Hortic. Sci. Biotechnol. 1993, 68, 791–795. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Abadía, J.; Marjani, M. Melatonin foliar sprays elicit salinity stress tolerance and enhance fruit yield and quality in strawberry (Fragaria × ananassa Duch.). Plant Physiol. Biochem. 2020, 149, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Kader, A.A. Flavor quality of fruits and vegetables. J. Sci. Food Agric. 2008, 88, 1863–1868. [Google Scholar] [CrossRef]
- Rouphael, Y.; Petropoulos, S.A.; Cardarelli, M.; Colla, G. Salinity as eustressor for enhancing quality of vegetables. Sci. Hortic. 2018, 234, 361–369. [Google Scholar] [CrossRef]
- Keutgen, A.; Pawelzik, E. Modifications of taste-relevant compounds in strawberry fruit under NaCl salinity. Food Chem. 2007, 105, 1487–1494. [Google Scholar] [CrossRef]
- D’Anna, F.; Incalcaterra, G.; Moncada, A.; Miceli, A. Effects of different electrical conductivity levels on strawberry grown in soilless culture. Acta Hortic. 2003, 609, 355–360. [Google Scholar] [CrossRef]
- Jamalian, S.; Tehranifar, A.; Tafazoli, E.; Eshghi, S.; Davarynejad, G.H. Paclobutrazol application ameliorates the negative effect of salt stress on reproductive growth, yield, and fruit quality of strawberry plants. Hortic. Environ. Biotechnol. 2008, 49, 203–208. [Google Scholar]
- Cardeñosa, V.; Medrano, E.; Lorenzo, P.; Sánchez-Guerrero, M.C.; Cuevas, F.J.; Pradas, I.; Moreno-Rojas, J.M. Effects of salinity and nitrogen supply on the quality and health-related compounds of strawberry fruits (Fragaria × ananassa cv. Primoris). J. Sci. Food Agric. 2015, 95, 2924–2930. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Cordenunsi, B.R.; Nascimento, J.R.O.; Lajolo, F.M. Physico-chemical changes related to quality of five strawberry fruit cultivars during cool-storage. Food Chem. 2003, 83, 167–173. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Pilbeam, D.J.; Gunes, A. Interactive effects of salicylic acid and silicon on oxidative damage and antioxidant activity in spinach (Spinacia oleracea L. cv. Matador) grown under boron toxicity and salinity. Plant Growth Regul. 2008, 55, 207–219. [Google Scholar] [CrossRef]
- Keutgen, A.J.; Keutgen, N. Influence of NaCl salinity stress on fruit quality in strawberry. Acta Hortic. 2003, 609, 155–157. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Relative Humidity (%) | Solar Radiation (W m−2) | |
|---|---|---|---|
| November | 18.9 | 63.5 | 130.6 |
| December | 15.5 | 59.8 | 128.1 |
| January | 12.6 | 66.1 | 124.1 |
| February | 15.3 | 54.0 | 181.1 |
| March | 17.9 | 59.0 | 261.4 |
| April | 18.6 | 64.2 | 260.6 |
| Temperature (°C) | Relative Humidity (%) | |
|---|---|---|
| November | 13.3 | 84.0 |
| December | 8.9 | 83.3 |
| January | 6.9 | 86.5 |
| February | 8.1 | 78.3 |
| March | 9.8 | 87.8 |
| April | 12.3 | 88.3 |
| Cultivar | Salt Treatment | Weight (g) | Diameter (mm) | Length (mm) | Diameter/ Length | Firmness (N) | Dry/Fresh Weight |
|---|---|---|---|---|---|---|---|
| Camarosa | Control | 19.20 c | 36.71 c | 33.89 b | 1.08 b | 1.68 a | 0.11 a |
| 2 dS m−1 | 18.60 d | 35.61 c | 34.81 ab | 1.02 b | 1.70 a | 0.08 b | |
| 4 dS m−1 | 15.19 d | 31.82 d | 31.41 c | 1.01 b | 1.62 a | 0.06 b | |
| Rociera | Control | 24.34 a | 45.60 a | 36.58 a | 1.25 a | 1.75 a | 0.12 a |
| 2 dS m−1 | 23.01 ab | 43.44 b | 35.91 ab | 1.21 a | 1.72 a | 0.09 b | |
| 4 dS m−1 | 20.69 b | 41.70 b | 34.31 b | 1.22 a | 1.69 a | 0.07 b |
| Cultivar | Salt Treatment | Nb Fruits/Plant | Extra (%) | I and II (%) | Total Yield/Plant (g) | Total Yield (kg) |
|---|---|---|---|---|---|---|
| Camarosa | Control | 9.21 ab | 97.82 a | 2.18 b | 130.74 ab | 4.31 d |
| 2 dS m−1 | 8.24 ab | 94.12 b | 5.88 ab | 103.13 b | 3.40 e | |
| 4 dS m−1 | 6.87 b | 92.82 b | 7.18 a | 92.85 b | 3.17 e | |
| Rociera | Control | 10.97 a | 97.02 a | 2.98 b | 277.26 a | 9.15 a |
| 2 dS m−1 | 10.52 a | 96.91 ab | 3.09 b | 229.08 ab | 7.56 b | |
| 4 dS m−1 | 9.91 ab | 93.06 b | 6.94 a | 214.13 ab | 7.07 c |
| Cultivar | Salt Treatment | pH | TSS (°Brix) | TA (% Citric Acid) | TSS:TA | L* | Chroma | Hue |
|---|---|---|---|---|---|---|---|---|
| Camarosa | Control | 3.34 a | 8.80 a | 1.12 a | 8.65 a | 36.48 a | 43.20 a | 33.75 a |
| 2 dS m−1 | 3.30 a | 8.39 a | 1.06 a | 8.67 a | 36.10 a | 43.17 a | 33.58 a | |
| 4 dS m−1 | 3.33 a | 8.28 a | 1.19 a | 7.63 a | 35.75 a | 40.80 a | 32.60 a | |
| Rociera | Control | 3.31 a | 9.00 a | 1.06 a | 9.45 a | 36.89 a | 41.32 a | 32.54 a |
| 2 dS m−1 | 3.26 a | 8.09 a | 0.98 a | 8.91 a | 35.85 a | 40.94 a | 32.64 a | |
| 4 dS m−1 | 3.27 a | 8.06 a | 1.12 a | 8.15 a | 34.90 a | 40.25 a | 32.59 a |
| Cultivar | Salt Treatment | Total Phenols | Total o-diphenols | Total Flavanols | Total Flavonoids | Total Anthocyanins | FRAP | DPPH |
|---|---|---|---|---|---|---|---|---|
| Camarosa | Control | 3.43 b | 5.22 a | 0.42 a | 0.46 b | 0.54 b | 20.43 b | 11.02 b |
| 2 dS m−1 | 4.02 ab | 3.67 a | 0.43 a | 0.54 ab | 0.59 a | 20.77 b | 12.29 ab | |
| 4 dS m−1 | 4.56 a | 4.50 a | 0.51 a | 0.59 a | 0.61 a | 24.20 a | 13.89 a | |
| Rociera | Control | 4.45 a | 3.30 a | 0.52 a | 0.58 a | 0.34 d | 19.37 b | 12.21 ab |
| 2 dS m−1 | 4.83 a | 4.01 a | 0.55 a | 0.62 a | 0.38 c | 25.50 a | 13.98 a | |
| 4 dS m−1 | 4.21 ab | 4.76 a | 0.46 a | 0.59 a | 0.40 c | 24.74 a | 13.63 a |
| Cultivar | Salt Treatment | Sucrose | Glucose | Fructose | Total Sugars | SI |
|---|---|---|---|---|---|---|
| Camarosa | Control | 17.49 bc | 9.30 b | 9.85 c | 36.64 c | 55.56 c |
| 2 dS m−1 | 23.64 a | 9.33 b | 12.23 abc | 45.21 ab | 69.37 bc | |
| 4 dS m−1 | 24.39 a | 10.21 ab | 12.91 ab | 47.51 a | 72.83 a | |
| Rociera | Control | 19.70 b | 11.90 a | 13.95 a | 45.55 ab | 70.58 ab |
| 2 dS m−1 | 18.72 bc | 9.17 b | 12.09 abc | 39.98 bc | 62.25 bc | |
| 4 dS m−1 | 16.13 c | 8.34 c | 10.58 bc | 35.05 c | 54.45 c |
| Cultivar | Salt Treatment | Ascorbic Acid | Malic Acid | Citric Acid | Total Acids |
|---|---|---|---|---|---|
| Control | 0.50 ab | 3.64 a | 6.38 b | 10.54 c | |
| Camarosa | 2 dS m−1 | 0.45 b | 4.26 a | 8.72 ab | 13.43 b |
| 4 dS m−1 | 0.39 b | 4.57 a | 9.27 a | 14.23 a | |
| Control | 0.60 a | 4.87 a | 5.41 b | 10.88 c | |
| Rociera | 2 dS m−1 | 0.57 a | 5.04 a | 6.72 b | 12.35 b |
| 4 dS m−1 | 0.51 ab | 5.21 a | 8.13 ab | 13.86 b |
| Cultivar | Salt Treatment | Root FW | Root DW | AGPM FW | AGPM DW | Total Plant FW | Total Plant DW | Root/AGPM FW | Root/AGPM DW |
|---|---|---|---|---|---|---|---|---|---|
| Camarosa | Control | 140.67 a | 27.44 a | 162.35 a | 49.07 a | 303.01 a | 76.51 a | 0.85 a | 0.55 a |
| 2 dS m−1 | 86.64 ab | 16.95 ab | 145.54 ab | 40.12 ab | 232.18 b | 57.07 bc | 0.63 ab | 0.46 a | |
| 4 dS m−1 | 73.24 ab | 13.98 ab | 126.43 bc | 37.22 ab | 199.66 bc | 51.20 bc | 0.61 ab | 0.39 a | |
| Rociera | Control | 89.78 ab | 17.69 ab | 148.51 ab | 46.38 ab | 238.29 b | 64.07 ab | 0.65 ab | 0.42 a |
| 2 dS m−1 | 70.28 b | 15.16 ab | 123.11 bc | 38.51 ab | 193.40 bc | 53.67 bc | 0.58 ab | 0.40 a | |
| 4 dS m−1 | 51.41 b | 10.33 b | 107.50 c | 33.61 b | 158.92 c | 43.95 c | 0.50 b | 0.32 a |
| Cultivar | Salt Treatment | Nb of Leaves | AGPM Water Content (g) | Root Water Content (g) | Plant Water Content (g) | Tolerance Index (%) |
|---|---|---|---|---|---|---|
| Camarosa | Control | 18 a | 113.28 a | 113.22 a | 226.50 a | 100 a |
| 2 dS m−1 | 14 c | 105.43 ab | 69.68 ab | 175.11 b | 74.59 b | |
| 4 dS m−1 | 11 d | 89.20 bc | 59.26 ab | 148.46 bc | 66.91 b | |
| Rociera | Control | 19 a | 102.13 ab | 72.09 ab | 174.22 b | 100 a |
| 2 dS m−1 | 16 b | 84.60 bc | 55.13 b | 139.73 bc | 83.76 ab | |
| 4 dS m−1 | 13 c | 73.89 c | 41.09 b | 114.97 c | 68.58 b |
| Camarosa | Rociera | |||
|---|---|---|---|---|
| Factors | Component 1 | Component 2 | Component 1 | Component 2 |
| FRAP | 0.11 | 0.228 | 0.0674 | −0.29 |
| DPPH | 0.128 | 0.148 | 0.0382 | −0.0931 |
| Total anthocyanins | 0.227 | 0.0508 | 0.132 | −0.124 |
| Total phenols | 0.187 | 0.201 | −0.0989 | 0.00807 |
| Total o-diphenols | −0.109 | 0.154 | 0.0479 | 0.0644 |
| Total flavonoids | 0.205 | 0.0574 | −0.0556 | −0.203 |
| Total flavanols | 0.121 | 0.18 | −0.146 | −0.134 |
| Salt tolerance index | −0.279 | 0.0969 | −0.288 | −0.114 |
| Nb of leaves | −0.192 | −0.0939 | −0.272 | 0.048 |
| AGPM water content | −0.21 | 0.101 | −0.253 | 0.0459 |
| Root water content | −0.242 | 0.109 | −0.184 | −0.201 |
| Plant water content | −0.268 | 0.123 | −0.289 | −0.113 |
| Root FW | −0.245 | 0.108 | −0.177 | −0.204 |
| Root DW | −0.253 | 0.106 | −0.134 | −0.198 |
| AGPM FW | −0.222 | 0.0926 | −0.261 | 0.0363 |
| AGPM DW | −0.229 | 0.0608 | −0.267 | 0.00987 |
| Plant FW | −0.273 | 0.119 | −0.292 | −0.114 |
| Plant DW | −0.279 | 0.0969 | −0.288 | −0.114 |
| pH | −0.0245 | 0.163 | −0.0793 | 0.0717 |
| TSS | −0.034 | −0.06 | −0.0686 | 0.103 |
| TA | 0.0473 | 0.0844 | −0.0487 | −0.178 |
| Total yield | −0.117 | −0.0179 | −0.0526 | −0.204 |
| Total yield/plant | −0.117 | −0.0179 | −0.0526 | −0.204 |
| Malic | 0.0983 | 0.00574 | 0.00304 | 0.0812 |
| ASA | −0.0464 | 0.119 | 0.104 | −0.04 |
| Citric | −0.1 | −0.16 | −0.134 | −0.0762 |
| Total organic acids | 0.0788 | −0.0689 | −0.00655 | 0.0405 |
| Fructose | 0.0265 | 0.303 | −0.197 | 0.257 |
| Glucose | 0.0647 | 0.282 | −0.136 | 0.302 |
| Sucrose | 0.195 | 0.164 | −0.0167 | 0.328 |
| Total sugars | 0.145 | 0.285 | −0.137 | 0.343 |
| SI | 0.135 | 0.299 | −0.154 | 0.332 |
| Weight | 0.0143 | −0.286 | 0.178 | −0.0434 |
| Length | −0.00334 | −0.335 | 0.131 | 0.0556 |
| Diameter | 0.0861 | −0.253 | 0.216 | −0.0737 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denaxa, N.-K.; Nomikou, A.; Malamos, N.; Liveri, E.; Roussos, P.A.; Papasotiropoulos, V. Salinity Effect on Plant Growth Parameters and Fruit Bioactive Compounds of Two Strawberry Cultivars, Coupled with Environmental Conditions Monitoring. Agronomy 2022, 12, 2279. https://doi.org/10.3390/agronomy12102279
Denaxa N-K, Nomikou A, Malamos N, Liveri E, Roussos PA, Papasotiropoulos V. Salinity Effect on Plant Growth Parameters and Fruit Bioactive Compounds of Two Strawberry Cultivars, Coupled with Environmental Conditions Monitoring. Agronomy. 2022; 12(10):2279. https://doi.org/10.3390/agronomy12102279
Chicago/Turabian StyleDenaxa, Nikoleta-Kleio, Aggeliki Nomikou, Nikolaos Malamos, Eleni Liveri, Petros Anargyrou Roussos, and Vasileios Papasotiropoulos. 2022. "Salinity Effect on Plant Growth Parameters and Fruit Bioactive Compounds of Two Strawberry Cultivars, Coupled with Environmental Conditions Monitoring" Agronomy 12, no. 10: 2279. https://doi.org/10.3390/agronomy12102279