
Biomass Allocation and Competitive Ability of a Semiarid Perennial Grass and a Legume in Mixtures under Periodical Soil Water Decreasing Conditions
 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pot Experiment
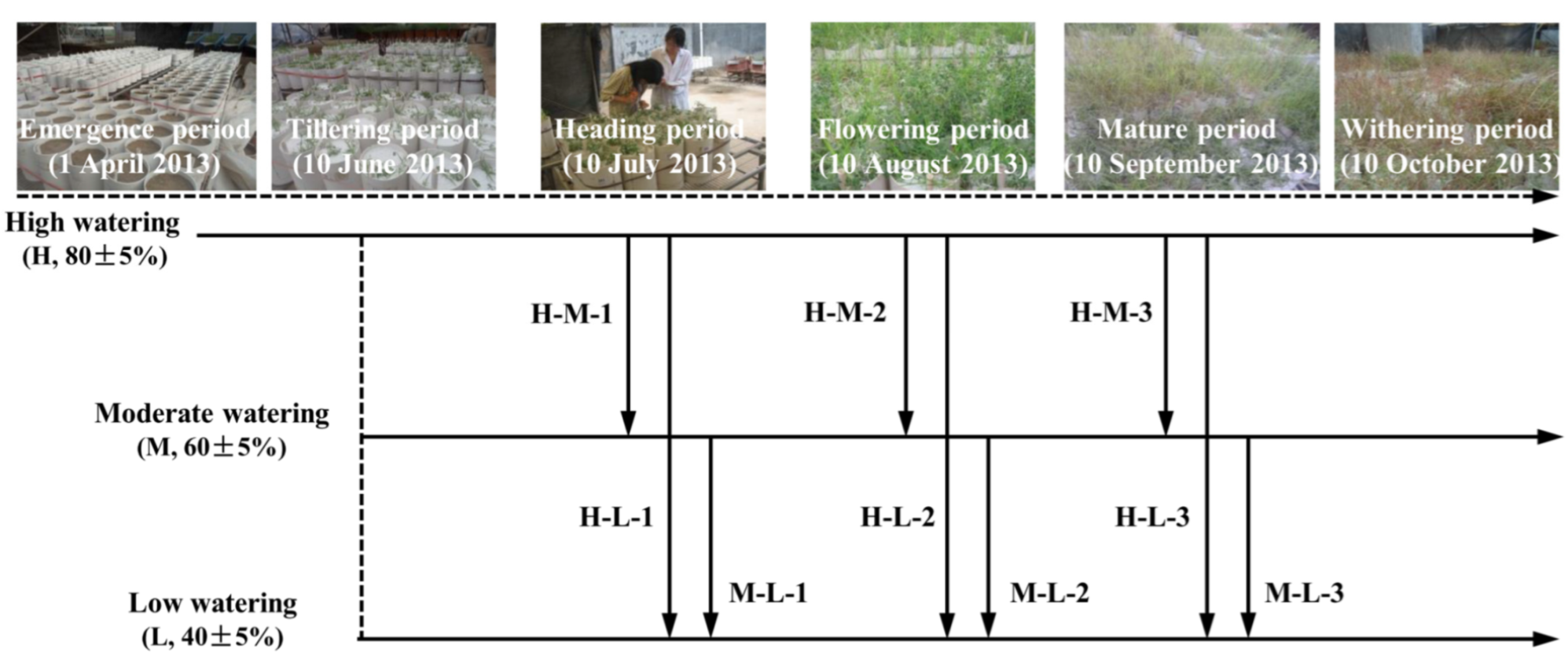
2.2. Plant Mixtures and Watering Treatments
2.3. Biomass Harvest
2.4. Water Use Efficiency
2.5. Competitive Indices
2.6. Statistical Analysis
3. Results
3.1. Plant Biomass Production (BP)
3.2. Root/Shoot Ratio (RSR)
3.3. Relative Yield Total (RYT)
3.4. Relative Competition Intensity (RCI)
3.5. Competitive Balance (CB)
3.6. Water Use Efficiency (WUE)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, R.; Fu, B.; Liu, G.; Liu, S. Soil carbon sequestration potential for “Grain for Green” project in Loess Plateau, China. Environ. Manag. 2011, 48, 1158–1172. [Google Scholar] [CrossRef]
- Jia, X.; Shao, M.; Wei, X. Richness and composition of herbaceous species in restored shrubland and grassland ecosystems in the northern Loess Plateau of China. Biodivers. Conserv. 2011, 20, 3435–3452. [Google Scholar] [CrossRef]
- Noy-Meir, I. Desert ecosystems: Environment and producers. Annu. Rev. Ecol. Evol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Yang, L.; Wei, W.; Mo, B.; Chen, L. Soil water deficit under different artificial vegetation restoration in the semi-arid hilly region of the Loess Plateau. Acta Ecol. Sin. 2011, 31, 3060–3068. [Google Scholar]
- Zhang, X.; Zhao, W.; Liu, Y.; Fang, X.; Feng, Q. The relationships between grasslands and soil moisture on the Loess Plateau of China: A review. Catena 2016, 145, 56–67. [Google Scholar] [CrossRef]
- Peng, S.; Gang, C.; Cao, Y.; Chen, Y. Assessment of climate change trends over the Loess Plateau in China from 1901 to 2100. Int. J. Climatol. 2018, 38, 2250–2264. [Google Scholar] [CrossRef]
- Miao, C.; Sun, Q.; Duan, Q.; Wang, Y. Joint analysis of changes in temperature and precipitation on the Loess Plateau during the period 1961–2011. Clim. Dyn. 2016, 47, 3221–3234. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y.; Xu, G.; Zou, T. Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation. Plant Cell Environ. 2007, 30, 399–409. [Google Scholar] [CrossRef]
- Anjum, S.A.; Xie, X.Y.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Niu, F.; Duan, D.; Chen, J.; Xiong, P.; Zhang, H.; Wang, Z.; Xu, B. Eco-physiological responses of dominant species to watering in a natural grassland community on the semi-arid Loess Plateau of China. Front. Plant Sci. 2016, 7, 663. [Google Scholar] [CrossRef] [PubMed]
- Xiong, P.; Shu, J.; Zhang, H.; Jia, Z.; Song, J.; Palta, J.A.; Xu, B. Small rainfall pulses affected leaf photosynthesis rather than biomass production of dominant species in semiarid grassland community on Loess Plateau of China. Funct. Plant Biol. 2017, 44, 1229–1242. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Wang, F.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T. Adaptation to and recovery from drought stress at vegetative stages in wheat (Triticum aestivum) cultivars. Funct. Plant Biol. 2016, 43, 1159–1169. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Xu, W.; Deng, X.; Xu, B.; Palta, J.A.; Chen, Y. Soil water availability changes in amount and timing favor the growth and competitiveness of grass than a co-dominant legume in their mixtures. Front. Plant Sci. 2021, 12, 723839. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Martin-Guay, M.O.; Paquette, A.; Dupras, J.; Rivest, D. The new green revolution: Sustainable intensification of agriculture by intercropping. Sci. Total Environ. 2018, 615, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Zhao, Y.; Li, X.; He, F.; Wan, L.; Wang, D.; Han, D. Relationships between grasses and legumes in mixed grassland: A review. Acta Prataculturae Sin. 2013, 22, 284–296. [Google Scholar]
- Xu, B.; Gao, Z.; Wang, J.; Xu, W.; Palta, J.A.; Chen, Y. N:P ratio of the grass Bothriochloa ischaemum mixed with the legume Lespedeza davurica under varying water and fertilizer supplies. Plant Soil 2016, 400, 67–79. [Google Scholar] [CrossRef]
- Nyfeler, D.; Huguenin-Elie, O.; Suter, M.; Frossard, E.; Connolly, J.; Lüscher, A. Strong mixture effects among four species in fertilized agricultural grassland led to persistent and consistent transgressive overyielding. J. Appl. Ecol. 2009, 46, 683–691. [Google Scholar] [CrossRef]
- Horton, J.L.; Hart, S.C. Hydraulic lift: A potentially important ecosystem process. Trends Ecol. Evol. 1998, 13, 232–235. [Google Scholar] [CrossRef]
- Ta, T.C.; Faris, M.A. Species variation in the fixation and transfer of nitrogen from legumes to associated grasses. Plant Soil 1987, 98, 265–274. [Google Scholar] [CrossRef]
- White, P.J.; George, T.S.; Gregory, P.J.; Bengough, A.G.; Hallett, P.D.; McKenzie, B.M. Matching roots to their environment. Ann. Bot. 2013, 112, 207–222. [Google Scholar] [CrossRef]
- Nyfeler, D.; Huguenin-Elie, O.; Suter, M.; Frossard, E.; Lüscher, A. Grass–legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources. Agric. Ecosyst. Environ. 2011, 140, 155–163. [Google Scholar] [CrossRef]
- Aasim, M. Yield and competition indices of intercropping cotton (Gossypium hirsutum L.) using different planting patterns. J. Agric. Sci. 2008, 14, A-5949. [Google Scholar]
- Yin, W.; Chai, Q.; Zhao, C.; Yu, A.; Fan, Z.; Hu, F.; Fan, H.; Guo, Y.; Coulter, J.A. Water utilization in intercropping: A review. Agric. Water Manag. 2020, 241, 106335. [Google Scholar] [CrossRef]
- Xu, B.C.; Xu, W.Z.; Huang, J.; Shan, L.; Li, F.M. Biomass allocation, relative competitive ability and water use efficiency of two dominant species in semiarid Loess Plateau under water stress. Plant Sci. 2011, 181, 644–651. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, W.; Chen, Z.; Jia, Z.; Huang, J.; Wen, Z.; Chen, Y.; Xu, B. Soil moisture availability at early growth stages strongly affected root growth of Bothriochloa ischaemum when mixed with Lespedeza davurica. Front. Plant Sci. 2018, 9, 1050. [Google Scholar] [CrossRef]
- Sammul, M.; Kull, K.; Oksanen, L.; Veromann, P. Competition intensity and its importance: Results of field experiments with Anthoxanthum odoratum. Oecologia 2000, 125, 18–25. [Google Scholar] [CrossRef]
- Wilson, J.B. Shoot competition and root competition. J. Appl. Ecol. 1988, 25, 279–296. [Google Scholar] [CrossRef]
- Williams, A.C.; Mccarthy, B.C. A new index of interspecific competition for replacement and additive designs. Ecol. Res. 2001, 16, 29–40. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P.; Xu, G.C.; Xiao, L.; Ren, Z.P.; Li, Z.B. Growth, morphological, and physiological responses to drought stress in Bothriochloa ischaemum. Front. Plant Sci. 2017, 8, 230. [Google Scholar] [CrossRef]
- Yang, L.; Lai, L.; Zhou, J.; Li, Q.; Yi, S.; Sun, Q.; Zheng, Y. Changes in levels of enzymes and osmotic adjustment compounds in key species and their relevance to vegetation succession in abandoned croplands of a semiarid sandy region. Ecol. Evol. 2020, 10, 2269–2280. [Google Scholar] [CrossRef]
- Xu, W.Z.; Deng, X.P.; Xu, B.C.; Gao, Z.J.; Ding, W.L. Photosynthetic activity and efficiency of Bothriochloa ischaemum and Lespedeza davurica in mixtures across growth stages under water stress. Acta Physiol. Plant. 2014, 36, 1033–1044. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, H.; Peng, Z.; Dai, J.; Zhao, F.; Chen, Z. Root system plays an important role in responses of plant to drought in the steppe of China. Land Degrad. Dev. 2021, 32, 3498–3506. [Google Scholar] [CrossRef]
- Eck, H.V.; Mathers, A.C.; Musick, J.T. Plant water stress at various growth stages and growth and yield of soybeans. Field Crops Res. 1987, 17, 1–16. [Google Scholar] [CrossRef]
- Wagg, C.; Hann, S.; Kupriyanovich, Y.; Li, S. Timing of short period water stress determines potato plant growth, yield and tuber quality. Agric. Water Manag. 2021, 247, 106731. [Google Scholar] [CrossRef]
- Schulze, E.D. Whole-plant responses to drought. Funct. Plant Biol. 1986, 13, 127–141. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S.; Mooney, H.A. Resource limitation in plants—An economic analogy. Annu. Rev. Ecol. Evol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Duan, D.; Jiang, F.; Lin, W.; Tian, Z.; Wu, N.; Feng, X.; Chen, T.; Nan, Z. Effects of drought on the growth of Lespedeza davurica through the alteration of soil microbial communities and nutrient availability. J. Fungi 2022, 8, 384. [Google Scholar] [CrossRef]
- Fry, E.L.; Evans, A.L.; Sturrock, C.J.; Bullock, J.M.; Bardgett, R.D. Root architecture governs plasticity in response to drought. Plant Soil 2018, 433, 189–200. [Google Scholar] [CrossRef]
- Xu, B.C.; Niu, F.R.; Duan, D.P.; Xu, W.Z.; Huang, J. Root morphological characteristics of Lespedeza davurica (L.) intercropped with Bothriochloa ischaemum (L.) Keng under water stress and P application conditions. Pak. J. Bot. 2012, 44, 1857–1864. [Google Scholar]
- Zhang, B.; Cadotte, M.W.; Chen, S.; Tan, X.; You, C.; Ren, T.; Chen, M.; Wang, S.; Li, W.; Chu, C.; et al. Plants alter their vertical root distribution rather than biomass allocation in response to changing precipitation. Ecology 2019, 100, e02828. [Google Scholar] [CrossRef]
- Mucheru-Muna, M.; Pypers, P.; Mugendi, D.; Kung’u, J.; Mugwe, J.; Merckx, R.; Vanlauwe, B. A staggered maize–legume intercrop arrangement robustly increases crop yields and economic returns in the highlands of Central Kenya. Field Crops Res. 2010, 115, 132–139. [Google Scholar] [CrossRef]
- Harris, D.; Natarajan, M.; Willey, R.W. Physiological basis for yield advantage in a sorghum/groundnut intercrop exposed to drought. 1. Dry-matter production, yield, and light interception. Field Crops Res. 1987, 17, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.K.; Mohanty, M.; Bandyopadhyay, K.K.; Painuli, D.K.; Misra, A.K. Growth, competition, yield advantage and economics in soybean/pigeonpea intercropping system in semi-arid tropics of India: I. Effect of subsoiling. Field Crops Res. 2006, 96, 80–89. [Google Scholar] [CrossRef]
- Zhu, S.G.; Cheng, Z.G.; Batool, A.; Wang, Y.B.; Wang, J.; Zhou, R.; Khan, A.; Zhu, S.Y.; Yang, Y.M.; Wang, W.; et al. Plant facilitation shifts along with soil moisture and phosphorus gradients via rhizosphere interaction in the maize-grass pea intercropping system. Ecol. Indic. 2022, 139, 108. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef]
- Van Duivenbooden, N.; Pala, M.; Studer, C.; Bielders, C.L.; Beukes, D.J. Cropping systems and crop complementarity in dryland agriculture to increase soil water use efficiency: A review. NJAS Wagening. J. Life Sci. 2000, 48, 213–236. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | df | Total BP | WUE | ||
|---|---|---|---|---|---|
| F | p | F | p | ||
| SWD | 2 | 674.90 | <0.001 | 79.05 | <0.001 |
| GS | 2 | 145.31 | <0.001 | 13.41 | <0.001 |
| MR | 6 | 873.69 | <0.001 | 615.56 | <0.001 |
| SWD × GS | 4 | 15.65 | <0.001 | 1.86 | 0.140 |
| SWD × MR | 12 | 16.97 | <0.001 | 3.81 | <0.001 |
| GS × MR | 12 | 5.39 | <0.001 | 0.83 | 0.63 |
| SWD × GS × MR | 24 | 3.43 | <0.001 | 1.12 | 0.34 |
| Factor | df | B. ischaemum | L. davurica | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BP | RSR | BP | RSR | ||||||
| F | p | F | p | F | p | F | p | ||
| SWD | 2 | 1055.85 | <0.001 | 258.30 | <0.001 | 0.03 | 0.98 | 125.77 | <0.001 |
| GS | 2 | 178.19 | <0.001 | 46.22 | <0.001 | 12.81 | <0.001 | 148.13 | <0.001 |
| MR | 5 | 328.92 | <0.001 | 2.24 | 0.056 | 750.87 | <0.001 | 2.49 | 0.04 |
| SWD × GS | 3 | 10.30 | <0.001 | 7.15 | <0.001 | 2.43 | 0.05 | 27.93 | <0.001 |
| SWD × MR | 10 | 1.83 | 0.06 | 3.90 | <0.001 | 4.80 | <0.001 | 3.12 | 0.002 |
| GS × MR | 10 | 4.12 | <0.001 | 3.90 | <0.001 | 6.17 | <0.001 | 2.53 | 0.01 |
| SWD × GS × MR | 15 | 6.97 | <0.001 | 2.95 | <0.001 | 10.07 | <0.001 | 2.01 | 0.01 |
| Factor | df | RYT | RCI | CB | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B. ischaemum | L. davurica | Total | Root | Shoot | |||||||||
| F | p | F | p | F | p | F | p | F | p | F | p | ||
| SWD | 2 | 0.15 | 0.87 | 32.85 | <0.001 | 21.79 | <0.001 | 39.50 | <0.001 | 23.50 | <0.001 | 27.27 | <0.001 |
| GS | 2 | 0.71 | 0.50 | 15.66 | <0.001 | 44.87 | <0.001 | 38.70 | <0.001 | 30.98 | <0.001 | 24.79 | <0.001 |
| MR | 4 | 1.51 | 0.21 | 1045.38 | <0.001 | 17.44 | <0.001 | 1274.29 | <0.001 | 874.31 | <0.001 | 955.13 | <0.001 |
| SWD × GS | 3 | 9.08 | <0.001 | 38.55 | <0.001 | 6.10 | <0.001 | 36.96 | <0.001 | 17.37 | <0.001 | 40.61 | <0.001 |
| SWD × MR | 8 | 1.12 | 0.36 | 21.46 | <0.001 | 18.68 | <0.001 | 9.17 | <0.001 | 7.55 | <0.001 | 6.50 | <0.001 |
| GS × MR | 8 | 0.68 | 0.71 | 6.61 | <0.001 | 17.19 | <0.001 | 19.10 | <0.001 | 13.74 | <0.001 | 14.90 | <0.001 |
| SWD × GS × MR | 12 | 1.92 | 0.03 | 7.38 | <0.001 | 21.79 | <0.001 | 17.39 | <0.001 | 12.97 | <0.001 | 12.92 | <0.001 |
| Mixture Ratio | No Decreasing | Heading Period | ||||
|---|---|---|---|---|---|---|
| H | M | L | H-M-1 | H-L-1 | M-L-1 | |
| B2L10 | 1.13 ± 0.04 | 0.98 ± 0.03 | 1.17 ± 0.05 | 1.04 ± 0.03 | 1.02 ± 0.02 | 1.15 ± 0.03 |
| B4L8 | 1.05 ± 0.04 | 1.07 ± 0.05 | 1.17 ± 0.01 | 1.09 ± 0.05 | 1.01 ± 0.03 | 1.17 ± 0.05 |
| B6L6 | 1.08 ± 0.02 | 1.03 ± 0.03 | 1.12 ± 0.06 | 1.10 ± 0.09 | 1.05 ± 0.08 | 1.06 ± 0.06 |
| B8L4 | 1.01 ± 0.05 | 1.03 ± 0.10 | 1.15 ± 0.04 | 1.10 ± 0.06 | 1.01 ± 0.07 | 1.13 ± 0.08 |
| B10L2 | 1.04 ± 0.04 | 1.09 ± 0.05 | 1.15 ± 0.03 | 1.07 ± 0.07 | 1.11 ± 0.10 | 1.15 ± 0.03 |
| MixtureRatio | Flowering Period | Mature Period | ||||
| H-M-2 | H-L-2 | M-L-2 | H-M-3 | H-L-3 | M-L-3 | |
| B2L10 | 1.16 ± 0.08 | 1.03 ± 0.02 | 1.02 ± 0.03 | 1.05 ± 0.06 | 1.15 ± 0.06 | 0.95 ± 0.05 |
| B4L8 | 1.13 ± 0.04 | 1.12 ± 0.03 | 1.14 ± 0.02 | 1.04 ± 0.04 | 1.16 ± 0.04 | 0.98 ± 0.08 |
| B6L6 | 1.03 ± 0.01 | 1.02 ± 0.04 | 1.09 ± 0.03 | 1.11 ± 0.04 | 1.13 ± 0.08 | 1.00 ± 0.05 |
| B8L4 | 1.06 ± 0.04 | 1.10 ± 0.07 | 1.08 ± 0.08 | 1.03 ± 0.06 | 1.14 ± 0.04 | 1.01 ± 0.06 |
| B10L2 | 1.06 ± 0.03 | 1.10 ± 0.05 | 1.13 ± 0.05 | 1.06 ± 0.01 | 1.07 ± 0.02 | 1.14 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Niu, F.; Kang, F.; Xu, B.; Deng, X.; Palta, J.A.; Chen, Y. Biomass Allocation and Competitive Ability of a Semiarid Perennial Grass and a Legume in Mixtures under Periodical Soil Water Decreasing Conditions. Agronomy 2022, 12, 2256. https://doi.org/10.3390/agronomy12102256
Xu W, Niu F, Kang F, Xu B, Deng X, Palta JA, Chen Y. Biomass Allocation and Competitive Ability of a Semiarid Perennial Grass and a Legume in Mixtures under Periodical Soil Water Decreasing Conditions. Agronomy. 2022; 12(10):2256. https://doi.org/10.3390/agronomy12102256
Chicago/Turabian StyleXu, Weizhou, Furong Niu, Furen Kang, Bingcheng Xu, Xiping Deng, Jairo A. Palta, and Yinglong Chen. 2022. "Biomass Allocation and Competitive Ability of a Semiarid Perennial Grass and a Legume in Mixtures under Periodical Soil Water Decreasing Conditions" Agronomy 12, no. 10: 2256. https://doi.org/10.3390/agronomy12102256