
Marker-Assisted Pyramiding of Multiple Disease Resistance Genes in Coffee Genotypes (Coffea arabica)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
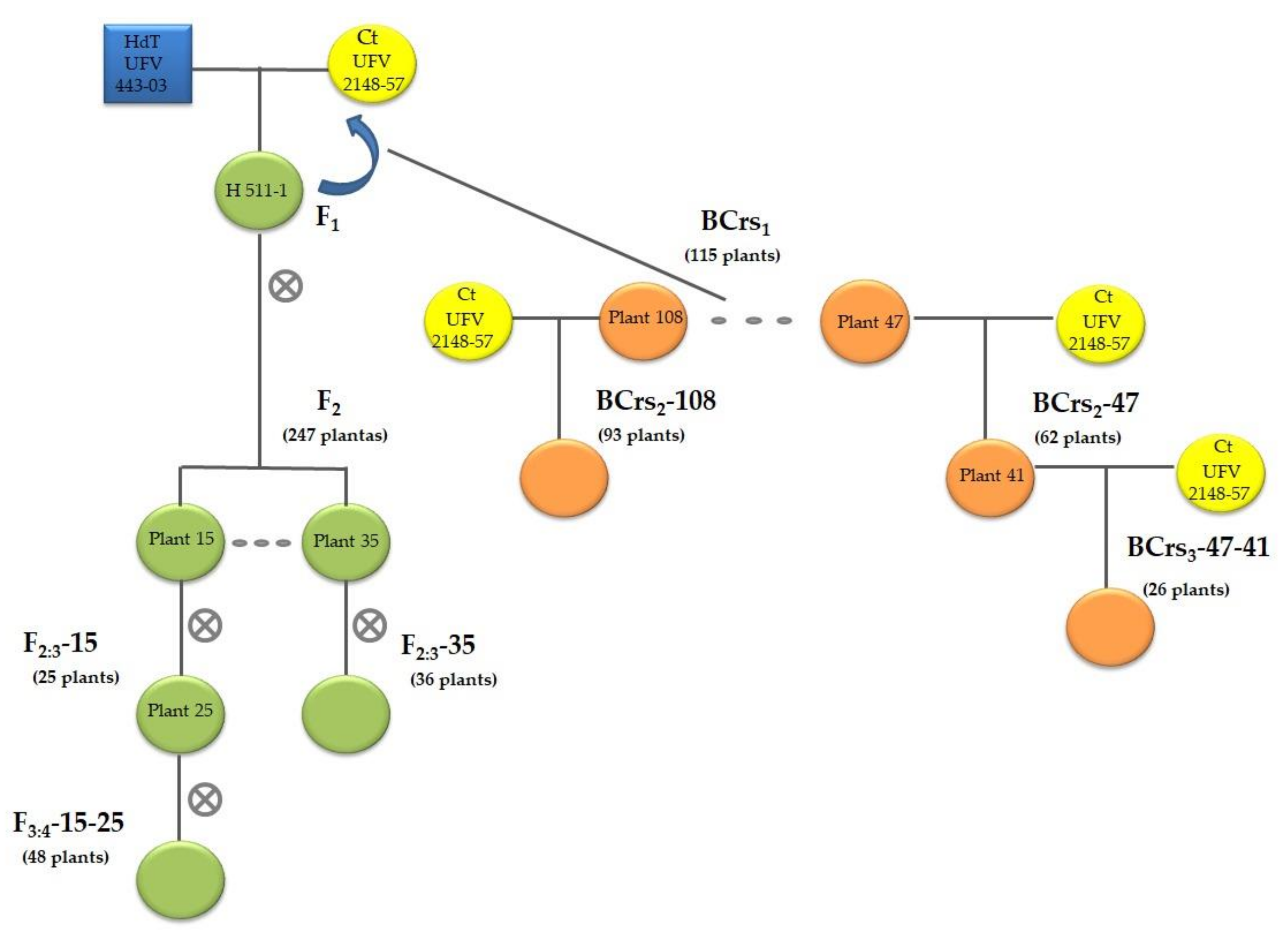
2.1. Plant Material and DNA Extraction
2.2. Disease Evaluation
2.3. SCAR and CAPS Marker Development
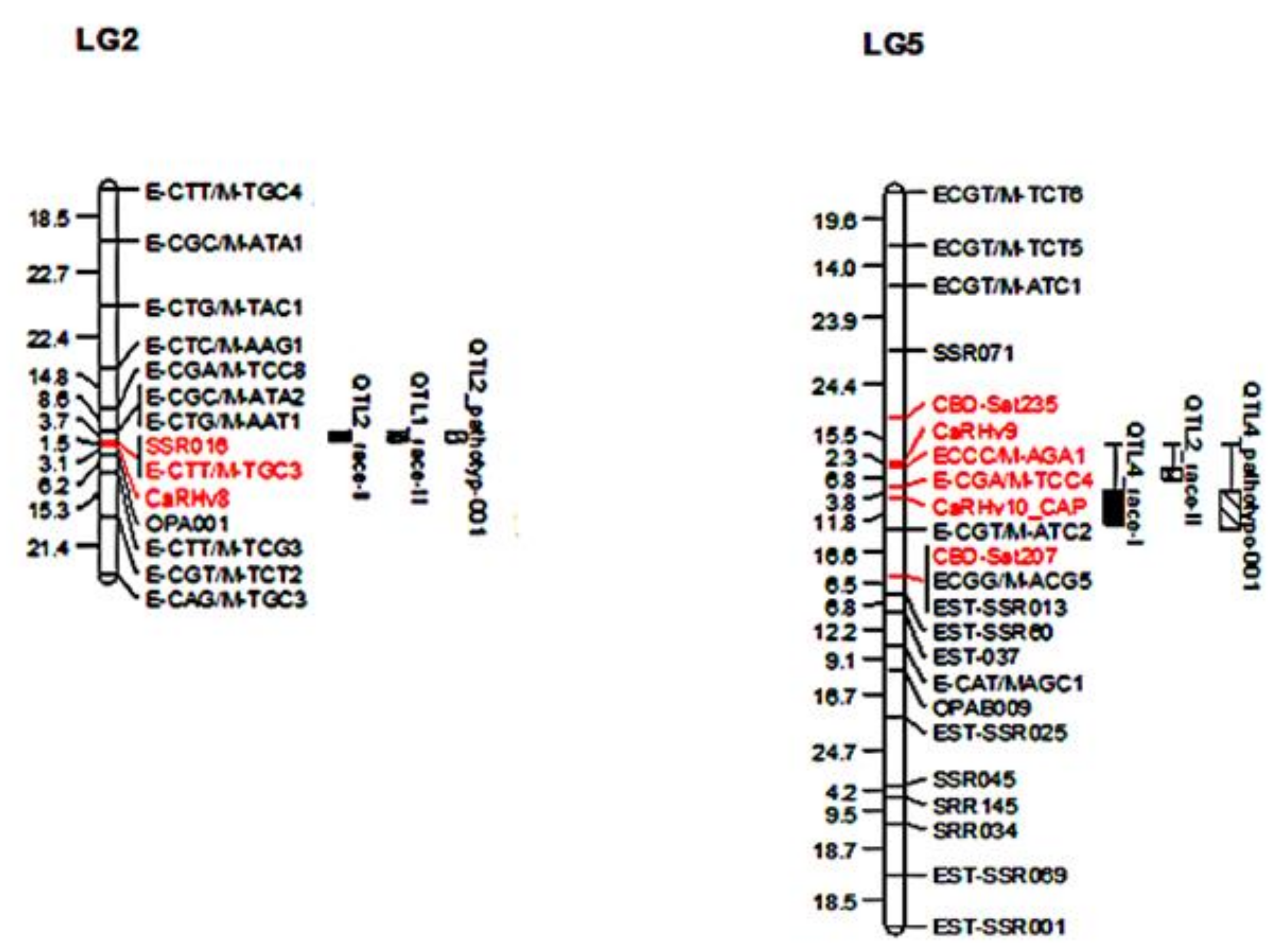
2.4. Linkage Map
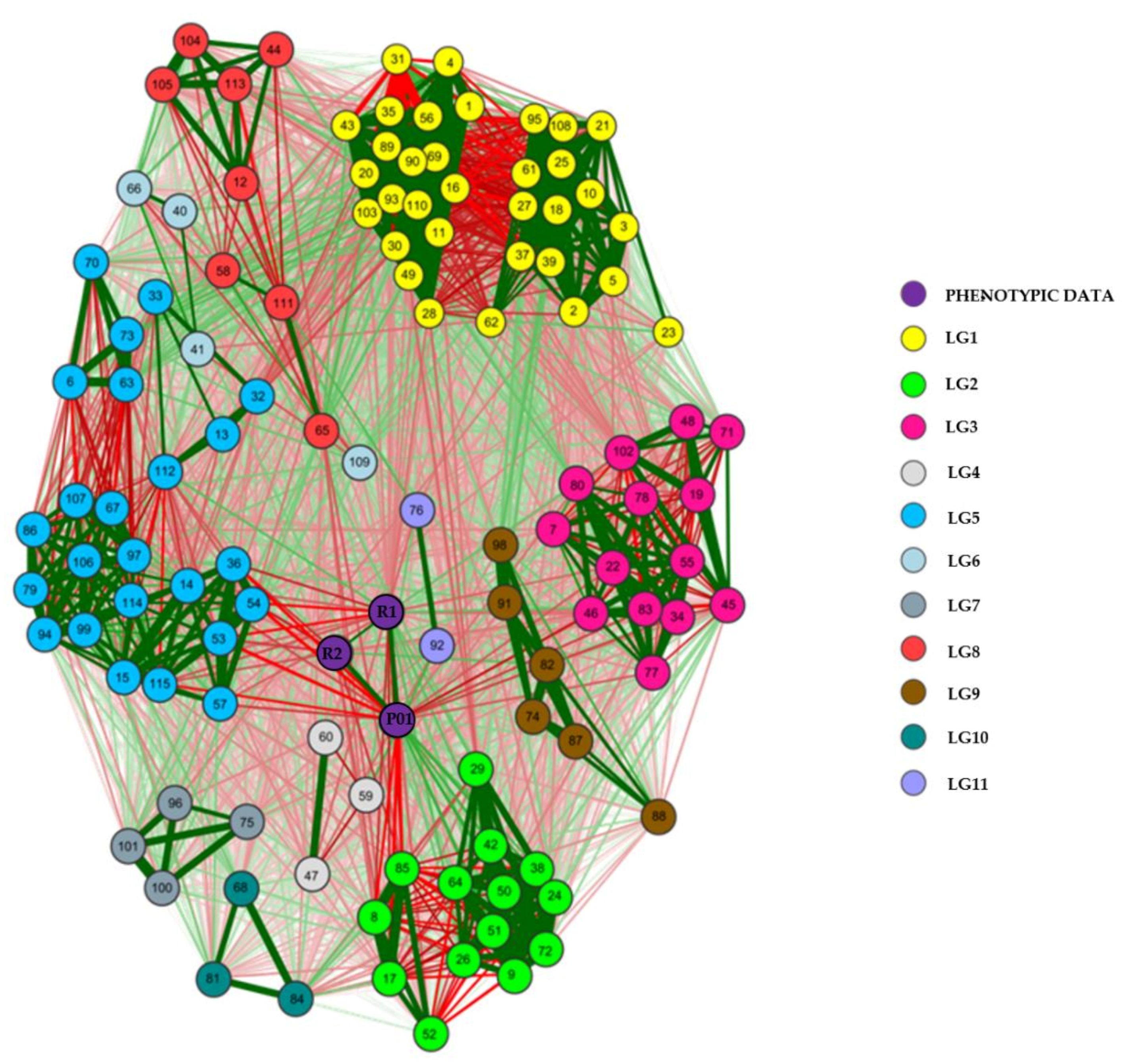
2.5. Molecular Marker-Assisted Selection Efficiency
2.6. Marker-Assisted Selection of Coffee Exhibiting Multiple Disease Resistance
3. Results
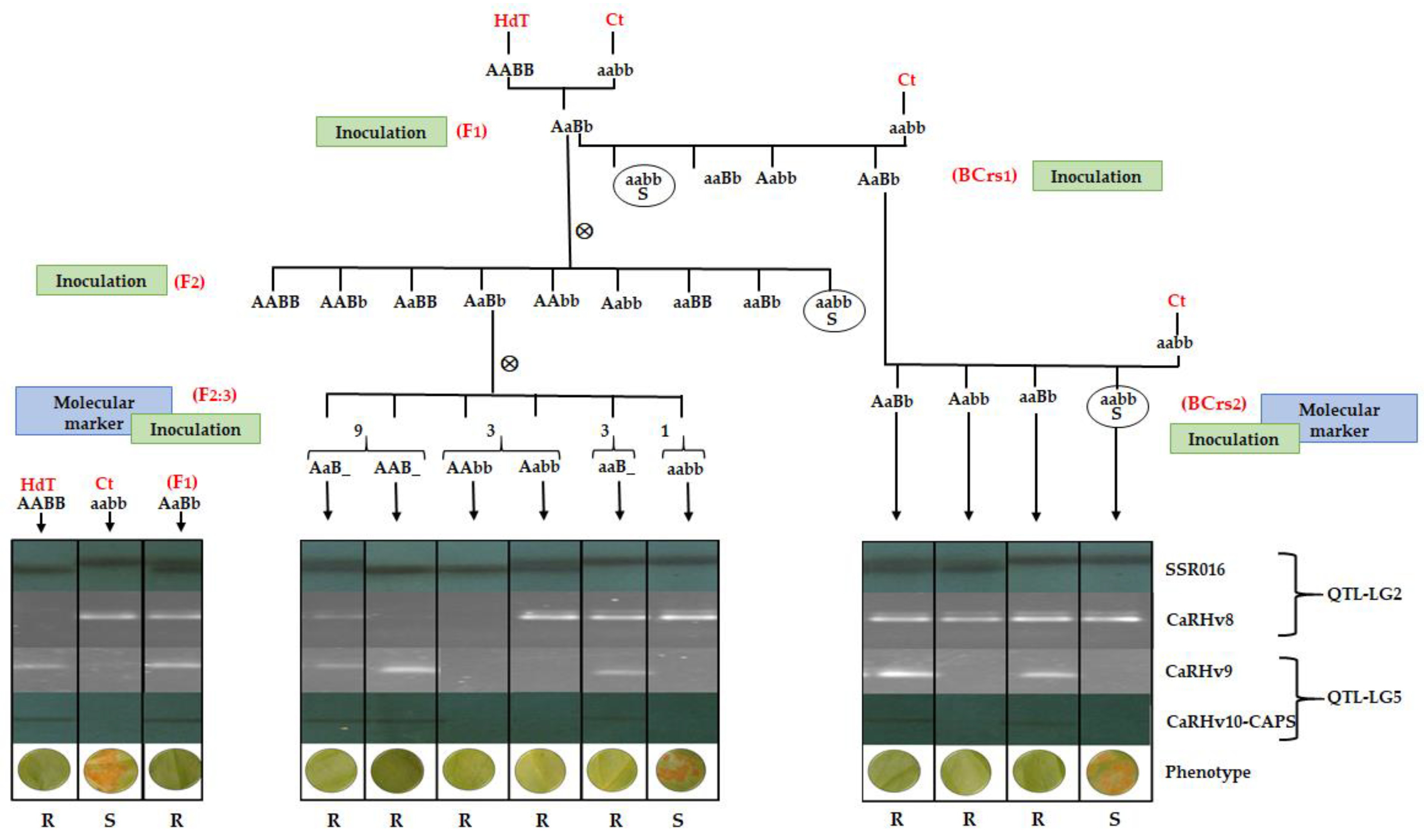
3.1. Molecular Marker Development
3.2. CaRHv8, CaRHv9, and CaRHv10_CAPS Marker Mapping
3.3. Molecular Marker-Assisted Selection Efficiency
3.4. Selection of F3:4 and BCrs3 Plants with Loci A, B, and C of CLR Resistance and Locus D of CBD Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef] [Green Version]
- Setotaw, T.A.; Caixeta, E.T.; Pereira, A.A.; Oliveira, A.C.B.; Cruz, C.D.; Zambolim, E.M.; Zambolim, L.; Sakyiama, N.S. Coefficient of Parentage in L. Cultivars Grown in Brazil. Crop Science 2013, 53, 1237. [Google Scholar] [CrossRef]
- Sousa, T.V.; Caixeta, E.T.; Alkimim, E.R.; Oliveira, A.C.B.; Pereira, A.A.; Sakiyama, N.S.; de Resende Júnior, M.F.; Zambolim, L. Population structure and genetic diversity of coffee progenies derived from Catuaí and Híbrido de Timor revealed by genome-wide SNP marker. Tree Genet. Genomes 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Merot-l'Anthoene, V.; Tournebize, R.; Darracq, O.; Rattina, V.; Lepelley, M.; Bellanger, L.; Tranchant-Dubreuil, C.; Coulée, M.; Pégard, M.; Metairon, S.; et al. Development and evaluation of a genome-wide Coffee 8.5K SNP array and its application for high-density genetic mapping and for investigating the origin of Coffea arabica L. Plant Biotechol. J. 2019, 17, 1418–1430. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, E.; Solano, W.; Gatica-Arias, A.; Chavarría, M.; Araya-Valverde, E. Microsatellite DNA fingerprinting of Coffea sp. germplasm conserved in Costa Rica through singleplex and multiplex PCR. Crop Breed. Appl. Biotechnol. 2020, 20. [Google Scholar] [CrossRef]
- Scalabrin, S.; Toniutti, L.; Gaspero, G.D.; Scaglione, D.; Magris, G.; Vidotto, M.; Pinosio, S.; Cattonaro, F.; Magni, F.; Jurman, I.; et al. A single polyploidization event at the origin of the tetraploid genome of Coffea arabica is responsible for the extremely low genetic variation in wild and cultivated germplasm. Sci. Rep. 2020, 10, 4642. [Google Scholar] [CrossRef] [Green Version]
- Zambolim, L.; Caixeta, E.T. An overview of physiological specialization of coffee leaf rust—New designation of pathotypes. Int. J. Curr. Res. 2021, 13, 15479–15490. [Google Scholar]
- Zambolim, L. Current status and management of coffee leaf rust in Brazil. Trop. Plant Pathol. 2016, 41, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Bettencourt, A.J.; Noronha-Wagner, M. Genetic factors conditioning resistance of Coffea arabica L. to Hemileia vastatrix Berk & Br. Agron. Lusit. 1971, 31, 285–292. [Google Scholar]
- Bettencourt, A.J.; Rodrigues, C.J. Principles and pratice of coffee breeding for resistance to rust and other diseases. Coffee Agron. 1988, 4, 199–234. [Google Scholar]
- Várzea, V.M.P.; Marques, D.V. Population variability of Hemileia vastatrix vs. coffee durable resistance. In Durable Reistance to Coffee Leaf Rust; Zambolim, L., Zambolim, E.M., Várea, V.M.P., Eds.; Universidade Federal de Viçosa: Viçosa, Brazil, 2005; pp. 53–74. [Google Scholar]
- Barka, G.D.; Caixeta, E.T.; Ferreira, S.S.; Zambolim, L. In silico guided structural and functional analysis of genes with potential involvement in resistance to coffee leaf rust: A functional marker based approach. PLoS ONE. 2020, 15, e0222747. [Google Scholar] [CrossRef]
- Almeida, D.P.; Castro, I.S.L.; Mendes, T.A.O.; Alves, D.R.; Barka, G.D.; Barreiros, P.R.R.M.; Zambolim, L.; Sakiyama, N.S.; Caixeta, E.T. Receptor-like kinase (Rlk) as a candidate gene conferring resistance to Hemileia vastatrix in coffee. Sci Agric. 2021, 78, 1–9. [Google Scholar] [CrossRef]
- Barbosa, I.P.; Costa, W.G.; Nascimento, M.; Cruz, C.D.; Oliveira, A.C.B. Recommendation of Coffea arabica genotypes by factor analysis. Euphytica 2019, 215, 178. [Google Scholar] [CrossRef]
- Sousa, T.V.; Caixeta, E.T.; Alkimim, E.R.; Oliveira, A.C.B.; Pereira, A.A.; Sakiyama, N.S.; Zambolim, L.; Resende, M.D.V. Early Selection Enabled by the Implementation of Genomic Selection in Coffea arabica Breeding. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setotaw, T.A.; Caixeta, E.T.; Zambolim, E.M.; Sousa, T.V.; Pereira, A.A.; Oliveira, A.C.B.; Cruz, C.D.; Zambolim, L.; Sakiyama, N.S. Genome Introgression of Híbrido de Timor and Its Potential to Develop High Cup Quality C. arabica Cultivars. J. Agric. Sci. 2020, 12, 64. [Google Scholar] [CrossRef] [Green Version]
- Alkimim, E.R.; Caixeta, E.T.; Sousa, T.V.; Pereira, A.A.; de Oliveira, A.C.B.; Zambolim, L.; Sakiyama, N.S. Marker-assisted selection provides arabica coffee with genes from other Coffea species targeting on multiple resistance to rust and coffee berry disease. Mol. Breed. 2017, 37, 6. [Google Scholar] [CrossRef] [Green Version]
- <named-content content-type="background:white">Lin, M.; Corsi, B.; Ficke, A.; Tan, K.C.; Cockram, J.; Lillemo, M. Genetic mapping using a wheat multi-founder population reveals a locus on chromosome 2A controlling resistance to both leaf and glume blotch caused by the necrotrophic fungal pathogen Parastagonospora nodorum. Theor. Appl. Genet. 2020, 133, 785–808. [Google Scholar]
- Muellner, A.E.; Eshonkulov, B.; Hagenguth, J.; Pachler, B.; Michel, S.; Buerstmayr, M.; Hole, D.; Buerstmayr, H. Genetic mapping of the common and dwarf bunt resistance gene Bt12 descending from the wheat landrace PI119333. Euphytica 2020, 216, 1–15. [Google Scholar] [CrossRef]
- Galiano-Carneiro, A.L.; Kessel, B.; Presterl, T.; Gaikpa, D.S.; Kistner, M.B.; Miedaner, T. Multi-parent QTL mapping reveals stable QTL conferring resistance to Gibberella ear rot in maize. Euphytica 2021, 217, 1–13. [Google Scholar] [CrossRef]
- Prakash, N.S.; Marques, D.V.; Varzea, V.M.P.; Silva, M.C.; Combes, M.C.; Lashermes, P. Introgression molecular analysis of a leaf rust resistance gene from Coffea liberica into C. arabica L. Theor. Appl. Genet. 2004, 109, 1311–1317. [Google Scholar] [CrossRef]
- Mahé, L.; Combes, M.-C.; Várzea, V.M.P.; Guilhaumon, C.; Lashermes, P. Development of sequence characterized DNA markers linked to leaf rust (Hemileia vastatrix) resistance in coffee (Coffea arabica L.). Mol. Breed. 2008, 21, 105–113. [Google Scholar] [CrossRef]
- Diola, V.; de Brito, G.G.; Caixeta, E.T.; Maciel-Zambolim, E.; Sakiyama, N.S.; Loureiro, M.E. High-density genetic mapping for coffee leaf rust resistance. Tree Genet. Genomes 2011, 7, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Romero, G.; Vásquez, L.M.; Lashermes, P.; Herrera, J.C. Identification of a major QTL for adult plant resistance to coffee leaf rust (Hemileia vastatrix) in the natural Timor hybrid (Coffea arabica x C. canephora). Plant Breed. 2014, 133, 121–129. [Google Scholar] [CrossRef]
- Pestana, K.N.; Capucho, A.S.; Caixeta, E.T.; de Almeida, D.P.; Zambolim, E.M.; Cruz, C.D.; Zambolim, L.; Pereira, A.A.; de Oliveira, A.C.B.; Sakiyama, N.S. Inheritance study and linkage mapping of resistance loci to Hemileia vastatrix in Híbrido de Timor UFV 443-03. Tree Genet. Genomes 2015, 11. [Google Scholar] [CrossRef] [Green Version]
- Gichuru, E.K.; Agwanda, C.O.; Combes, M.C.; Mutitu, E.W.; Ngugi, E.C.K.; Bertrand, B.; Lashermes, P. Identification of molecular markers linked to a gene conferring resistance to coffee berry disease (Colletotrichum kahawae) in Coffea arabica. Plant Pathol. 2008, 57, 1117–1124. [Google Scholar] [CrossRef]
- Gimase, J.M.; Thagana, W.M.; Omondi, C.O.; Cheserek, J.J.; Gichimu, B.M.; Gichuru, E.K. Quantitative trait loci (QTL) mapping of resistance to coffee berry disease (Colletotrichum kahawae Waller Bridge) in Coffea arabica L. variety Rume Sudan. Afr. J. Agric. Res. 2020, 16, 1184–1194. [Google Scholar] [CrossRef]
- Silva, R.A.; Zambolim, L.; Castro, I.S.L.; Rodrigues, H.S.; Cruz, C.D.; Caixeta, E.T. The Híbrido de Timor germplasm: Identification of molecular diversity and resistance sources to coffee berry disease and leaf rust. Euphytica 2018, 214, 153. [Google Scholar] [CrossRef]
- Bertrand, B.; Guyot, B.; Anthony, F.; Lashermes, P. Impact of the Coffea canephora gene introgression on the beverage quality of C. arabica. Theor. Appl. Genet. 2003, 107, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; Lopes, M.T.G.; Figueira, A.; Camargo, L.E.A.; Fungaro, M.H.P.; Carneiro, M.S.; Vieira, M.L.C. Marcadores moleculares dominantes (RAPD e AFLP). Biotecnologia Ciência Desenvolvimento 2002, 5, 56–60. [Google Scholar]
- Kumar, S.; Saxena, S. Sequence Characterized Amplified Regions Linked with Apomictic Mode of Reproduction in Four different apomictic Cenchrus Species. Mol. Plant Breed. 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Moncada, M.D.P.; Tovar, E.; Montoya, J.C.; González, A.; Spindel, J.; McCouch, S. A genetic linkage map of coffee (Coffea arabica L.) and QTL for yield, plant height, and bean size. Tree Genet. Genomes 2016, 12, 5. [Google Scholar] [CrossRef]
- Zhang, N.; Xu, B.; Bi, Y.; Lou1, Q.; Chen, J.; Qian, C. Development of a Muskmelon Cultivar with Improved Resistance to Gummy Stem Blight and Desired Agronomic Traits Using Gene Pyramiding. Czech J. Genet. Plant Breed. 2017, 53. [Google Scholar] [CrossRef] [Green Version]
- Diniz, L.E.C.; Sakiyama, N.S.; Lashermes, P.; Caixeta, E.T.; Oliveira, A.C.B.; Zambolim, E.M.; Loureiro, M.E.; Pereira, A.A.; Zambolim, L. Analysis of AFLP markers associated to the Mex-1 resistance locus in Icatu progenies. Crop. Breed. Appl. Biotechnol. 2005, 5, 387–393. [Google Scholar] [CrossRef]
- Zambolim, L.; Chaves, M.C. Efeito de baixas temperaturas e do binomio temperatura-umidade relativa sobre a viabilidade dos uredosporos de Hemileia vastatrix Berk. et Br. e Uromyces phaseoli typica arth. Experientiae 1974, 17, 151–184. [Google Scholar]
- Eskes, A.B.; Toma-Braghini, M. The effect of leaf age on incomplete resistance of coffee to Hemileia vastatrix. Netherlands J. Plant Pathol. 1982, 88, 219–230. [Google Scholar] [CrossRef]
- Tamayo, P.J.; Vale, F.X.R.; Zambolim, L.; Chaves, G.M.; Pereira, A.A. Resistência do Catimor à ferrugem e virulência de raças fisiológicas de Hemileia vastatrix Berk. & Br. Fitopatol. Bras. 1995, 20, 572–576. [Google Scholar]
- Caetano-Anollés, G.; Trigiano, R.N. Recovery of DNA Amplification Products from Silver-Stained polyacrrylamide Gels. Methods in MolecularBiology. In PCR Cloning Protocols-From Molecular Cloning to Genetic Engineering; White, B.A., Ed.; Humana Press: Totowa, NJ, USA, 1996; pp. 111–127. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Cruz, C.D. Programa Genes—Ampliado e integrado aos aplicativos R, Matlab e Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S.; Botstein, D. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 1989, 121, 185–199. [Google Scholar] [CrossRef]
- Rosado, R.D.S.; Rosado, L.D.S.; Borges, L.L.; Bruckner, C.H.; Cruz, C.D.; Dos Santos, C.E.M. Genetic diversity of sour passion fruit revealed by predicted genetic values. Agron. J. 2019, 111, 165–174. [Google Scholar] [CrossRef]
- Fruchterman, T.M.J.; Reingold, E.M. Graph drawing by force-directed placement. Softw. Pract. Exp. 1991, 21, 1129–1164. [Google Scholar] [CrossRef]
- Epskamp, S.; Cramer, A.O.; Waldorp, L.J.; Schmittmann, V.D.; Borsboom, D. qgraph: Network visualizations of relationships in psychometric data. J. Stat. Softw. 2012, 48, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Alzate-Marin, A.L.; Cervigni, G.D.; Moreira, M.A.; Barros, E.G. Seleção assistida por marcadores moleculares visando ao desenvolvimento de plantas resistentes a doenças, com ênfase em feijoeiro e soja. Fitopatol. Bras. 2005, 30, 333–342. [Google Scholar] [CrossRef] [Green Version]
- Alvarenga, S.M.; Caixeta, E.T.; Hufnagel, B.; Thiebaut, F.; Maciel-Zambolim, E.; Zambolim, L.; Sakiyama, N.S. Marcadores moleculares derivados de sequências expressas do genoma café potencialmente envolvidas na resistência à ferrugem. Pesquisa Agropecuária Brasileira 2011, 46, 890–898. [Google Scholar] [CrossRef]
- Combes, M.C.; Andrzejewski, S.; Anthony, F.; Bertrand, B.; Rovell, P.; Graziosi, G.; Sashermes, P. Characterization of microsatellites loci in Coffea arabica and related coffee species. Mol Ecol. 2000, 9, 1171–1193. [Google Scholar] [CrossRef]
- Prakash, N.; Devasia, J.; Das Divya, K.; Manjunatha, B.; Seetharam, H.; Kumar, A.; Jayarama. Breeding for rust resistance in Arabica—Where we are and what next? In Proceedings of the 25th International Conference on Coffee Science (ASIC), Armenia, Colombia, 8–13 September 2014; p. B10. [Google Scholar]
- Silva, D.N.; Várzea, V.; Paulo, O.S.; Batista, D. Population genomic footprints of host adaptation, introgression and recombination in coffee leaf rust. Mol. Plant Pathol. 2018, 19, 1742–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negi, M.S.; Devic, M.; Delseny, M. Identification of AFLP fragments linked to seed coat colour in Brassica juncea and conversion to a SCAR marker for rapid selection. Theor. Appl. Genet. 2000, 101, 146–152. [Google Scholar] [CrossRef]
- Jaber, E.H.A.; Srour, A.Y.; Zambounis, A.G.; Vakalounakis, D.J.; Doulis, A.G. Identification of SCAR markers linked to the Foc gene governing resistance to Fusarium oxysporum f. sp. cucumerinum in cucumber cv. SMR-18. Eur. J. Plant Pathol. 2020, 157, 845–855. [Google Scholar] [CrossRef]
- Wu, P.; Xie, J.; Hu, J.; Qiu, D.; Liu, Z.; Li, J.; Li, M.; Zhang, H.; Yang, L.; Liu, H.; et al. Development of molecular markers linked to powdery mildew resistance gene Pm4b by combining SNP discovery from transcriptome sequencing Data with Bulked Segregant Analysis (BSR-Seq) in Wheat. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Denoeud, F.; Carretero-Paulet, L.; Dereeper, A.; Droc, G.; Guyot, R.; Pietrella, M.; Zheng, C.; Alberti, A.; Anthony, F.; Aprea, G.; et al. The coffee genome provides insight into the convergent evolution of caffeine biosynthesis. Science 2014, 345, 1181–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrier, A.; Berthaud, J. Botanical classification of coffee. In Coffee: Botany, Biochemistry and Production of Beans and Beverage; Clifford, M.N., Wilson, K.C., Eds.; AVI Publishing Company: Westport, CT, USA, 1985; pp. 13–47. [Google Scholar]
- Clarindo, W.R.; Carvalho, C.R. First Coffea arabica karyogram showing that this species is a true allotetraploid. Plant Syst. Evol. 2008, 274, 237. [Google Scholar] [CrossRef]
- Van der Vossen, H.A.M. The cup quality of disease-resistant cultivars of Arabica coffee (Coffea arabica). Expl. Agric. 2009, 45, 323–332. [Google Scholar] [CrossRef]
- Valencia, A.; Morales, A.Y.; Moncada, P.M.I.; Cortina, H.A.; Herrera, J.C. Introgresion of the SH3 gene resistant to rust (Hemileia vastatrix) in improved lines of CASTILLO® variety (Coffea arabica L.). ournal Plant Breed. Crop Sci. 2017, 9, 130–138. [Google Scholar] [CrossRef]
- Prakash, N.S.; Muniswamy, B.; Hanumantha, B.T.; Sreenath, H.L.; Sundaresha; Kumar, D.; Suresh, N.; Santhosh, P.; Soumya, P.R.; Bhat, A.M.; et al. Marker assisted selection and breeding for leaf rust resistance in coffee (Coffea arabica L.) -some recent leads. Indian J. Genet. Plant Breed. 2011, 71, 185–189. [Google Scholar]
- Van der Vossen, H.A.M.; Walyaro, D.J. Additional evidence for oligogenic inheritance of durable host resistance to coffee berry disease (Colletotrichum kahawae) in arabica coffee (Coffea arabica L.). Euphytica 2009, 165, 105–111. [Google Scholar] [CrossRef]
- Horgan, F.G.; Bernal, C.C.; Vu, Q.; Almazan, M.L.P.; Ramal, A.F.; Yasui, H.; Fujita, D. Virulence adaptation in a rice leafhopper: Exposure to ineffective genes compromises pyramided resistance. Crop Prot. 2018, 113, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Pilet-Nayel, M.-L.; Moury, B.; Caffier, V.; Montarry, J.; Kerlan, M.-C.; Fournet, S.; Durel, C.-E.; Delourme, R. Quantitative Resistance to Plant Pathogens in Pyramiding Strategies for Durable Crop Protection. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Srivastava, P.; Mavi, G.S.; Kaur, S.; Kaur, J.; Bala, R.; Singh, T.P.; Sohu, V.S.; Chhuneja, P.; Bains, N.S. Resurrection of wheat cultivar ‘PBW343’using marker assisted gene pyramiding for rust resistance. Front. Plant Sci. 2021, 12, 42. [Google Scholar] [CrossRef]
- Das, G.; Rao, G.J.N.; Varier, M.; Prakash, A.; Prasad, D. Improved Tapaswini having four BB resistance genes pyramided with six genes/QTLs, resistance/tolerance to biotic and abiotic stresses in rice. Sci. Rep. 2018, 8, 2413. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Lu, J.; Zhou, M.; Zheng, S.; Liu, Z.; Zhang, C.; Du, M.; Wang, M.; Li, Y.; Wu, Y.; et al. Developing stripe rust resistant wheat (Triticum aestivum L.) lines with gene pyramiding strategy and marker-assisted selection. Genet. Resour. Crop Evol. 2020, 67, 381–391. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AFLP Marker | ||||
|---|---|---|---|---|
| E-CTG/MAAT1 | E-CTT/MTGC3 | E-CCC/MAGA1 | E-CGA/MTCC4 | |
| LG a | 2 | 2 | 5 | 5 |
| Linkage b | Coupled | Repulsion | Coupled | Coupled |
| Size AFLP c | 670 pb | 600 pb | 220 pb | 800 pb |
| Developed marker | CaRHv7 | CaRHv8 | CaRHv9 | CaRHv10_CAPS |
| Size SCAR d | 108 pb | 272 pb | 151 pb | 340 pb |
| Marker type | SCAR | SCAR | SCAR | CAPS |
| Primer F e | GAACGATTGAGGATGCCAAG | ACCTTCTAGTGTTACCGAGGA | TGATGAAGAAGAGCGCATAGC | GACACATCGTGAACGTGGAG |
| Primer R f | TGTCAAAGTGGTACGGAGGA | TCTTAGCGCCATGAATAGCCA | GTCTAAGACCAGAATCAGATGG | CAGCTGAACAACCGAACTCA |
| Annealing temp (°C) | 65 | 65 | 65 | 65 |
| Genotypic Markers Segregation | Phenotypic Disease Segregation | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breeding Population | Expect Ratio a | SSR16 CaRHv8 Locus A (QTL-LG2) | CaRHv9 CaRHv10_CAPS Locus B (QTL-LG5) | Obs Num Gen b | Expec Num Gen c | Chi-Squared Χ2 (P) | Expect Ratio d R:S | Obs Num e Phe | Expect Num f Phe | Chi-Squared Χ2 (P) | SE g | ||||
| R | S | R | S | ||||||||||||
| HdT (UFV443-03) | __ | AA | AA | B_ | B_ | AABB | AABB | ||||||||
| Ct (UFV 2148-57) | __ | aa | _a | Bb | bb | Aabb | Aabb | ||||||||
| F1 (H 511-1) | __ | Aa | _a | B_ | B_ | AaBb | AaBb | ||||||||
| BCrs2-47 | 1:1:1:1 | Aa | _a | bb | bb | 14 Aabb | 15.,5 Aabb | 1.74 (0.63) | 12R:4S | 47 | 15 | 47 | 15 | 0 (1) | 93.6 |
| Aa | _a | B_ | B_ | 18 AaB_ | 15.5 AaB_ | ||||||||||
| aa | _a | B_ | B_ | 18 aaB_ | 15.5 aaB_ | ||||||||||
| aa | _a | bb | bb | 12 aabb | 15.5 aabb | ||||||||||
| BCrs2-108 | 1:1:1:1 | Aa | _a | bb | bb | 22 Aabb | 23.25Aabb | 0.29 (0.96) | 12R:4S | 59 | 34 | 69 | 24 | 5.62 (0.18) | 90.0 |
| Aa | _a | B_ | B_ | 24 AaB_ | 23.25 AaB_ | ||||||||||
| aa | _a | B_ | B_ | 25 aaB_ | 23.25 aaB_ | ||||||||||
| aa | _a | bb | bb | 22 aabb | 23.25 aabb | ||||||||||
| F2:3 -15 | 9:3:3:1 | AA | AA | B_ | B_ | 7 AAB_ | 5 AAB_ | 4.58 (0.47) | 15R:1S | 25 | 0 | 24 | 1 | 1.04 (0.31) | 100 |
| AA | AA | bb | bb | 4 AAbb | 2 AAbb | ||||||||||
| Aa | _a | B_ | B_ | 7 AaB_ | 9 AaB_ | ||||||||||
| Aa | _a | bb | bb | 2 Aabb | 3 Aabb | ||||||||||
| aa | _a | B_ | B_ | 5 aaB_ | 5 aaB_ | ||||||||||
| aa | _a | bb | bb | Aabb | 1 aabb | ||||||||||
| F2:3-35 | 9:3:3:1 | AA | AA | B_ | B_ | 7 AAB_ | 7 AAB_ | 0.21 (0.99) | 15R:1S | 33 | 3 | 33 | 3 | 0 (1) | 97.3 |
| AA | AA | bb | bb | 2 AAbb | 2 AAbb | ||||||||||
| Aa | _a | B_ | B_ | 15 AaB_ | 14 AaB_ | ||||||||||
| Aa | _a | bb | bb | 4 Aabb | 4 Aabb | ||||||||||
| aa | _a | B_ | B_ | 6 aaB_ | 7 aaB_ | ||||||||||
| aa | _a | bb | bb | 2 aabb | 2 aabb | ||||||||||
| Populations | Locus A | Locus B | Locus C | Locus D | Genotype | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CaRHv8 | SSR16 | CaRHv9 | CaRHv10_CAPS | CARF005 | CBD-Sat207 | CBD-Sat235 | ||||||||||||
| -a | AA | AA | Aa | Aa | B- | bb | B- | Bb | C- | cc | DD | Dd | dd | DD | Dd | dd | ||
| HdT | AABBCCDD | |||||||||||||||||
| Ct | aabbccdd | |||||||||||||||||
| F3:4 -15-25 (48 plants) | 48 (AABBCCDD) | |||||||||||||||||
| BCrs3-47-41 (26 plants) | 6 (AaBbCcDd) | |||||||||||||||||
| 1 (AaBbCcdd) or (AaBbCcDd) | ||||||||||||||||||
| 1 (AaBbCcdd) | ||||||||||||||||||
| 5 (aaBbCcDd) | ||||||||||||||||||
| 1 (AaBbccDd) | ||||||||||||||||||
| 1 (Aabbccdd) | ||||||||||||||||||
| 3 (aaBbCcDd) | ||||||||||||||||||
| 2 (aaBbCcdd) | ||||||||||||||||||
| 1 (aabbCcdd) | ||||||||||||||||||
| 1 (aabbccDd) | ||||||||||||||||||
| 3 (aabbccdd) | ||||||||||||||||||
| 1 (AABBCCDD) | ||||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Almeida, D.P.; Caixeta, E.T.; Moreira, K.F.; de Oliveira, A.C.B.; de Freitas, K.N.P.; Pereira, A.A.; Rosado, R.D.S.; Zambolim, L.; Cruz, C.D. Marker-Assisted Pyramiding of Multiple Disease Resistance Genes in Coffee Genotypes (Coffea arabica). Agronomy 2021, 11, 1763. https://doi.org/10.3390/agronomy11091763
de Almeida DP, Caixeta ET, Moreira KF, de Oliveira ACB, de Freitas KNP, Pereira AA, Rosado RDS, Zambolim L, Cruz CD. Marker-Assisted Pyramiding of Multiple Disease Resistance Genes in Coffee Genotypes (Coffea arabica). Agronomy. 2021; 11(9):1763. https://doi.org/10.3390/agronomy11091763
Chicago/Turabian Stylede Almeida, Dênia Pires, Eveline Teixeira Caixeta, Karoliny Ferreira Moreira, Antonio Carlos Baião de Oliveira, Kátia Nogueira Pestana de Freitas, Antônio Alves Pereira, Renato Domiciano Silva Rosado, Laércio Zambolim, and Cosme Damião Cruz. 2021. "Marker-Assisted Pyramiding of Multiple Disease Resistance Genes in Coffee Genotypes (Coffea arabica)" Agronomy 11, no. 9: 1763. https://doi.org/10.3390/agronomy11091763