
Defense Response to Hemileia vastatrix in Susceptible Grafts onto Resistant Rootstock of Coffea arabica L.

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
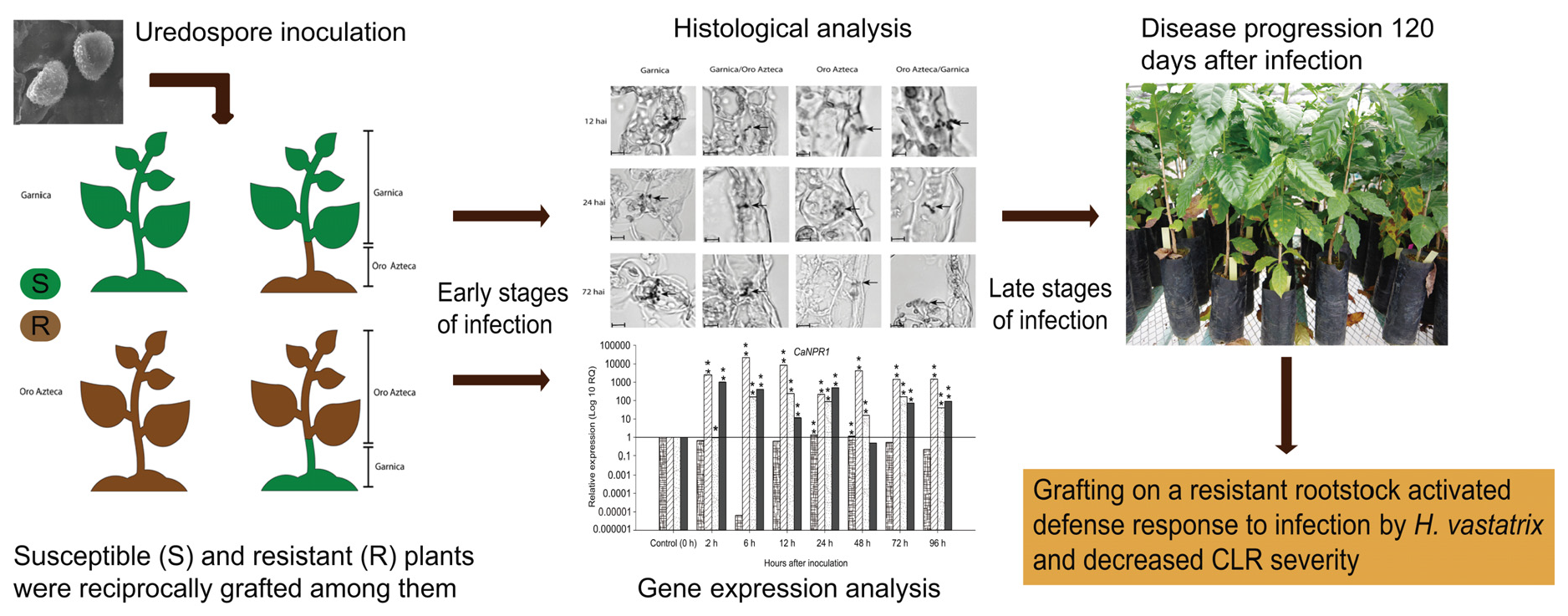
2.1. Experimental Design and Plant Material
2.2. Preparation of the Uredospore Inoculum
2.3. Inoculation and Incubation
2.4. Observation of the Development of Hemileia vastatrix during the First Stages of Infection in the Foliar Tissue
2.5. Selection of Target Genes
2.6. Gene Expression Analysis
2.6.1. RNA Extraction
2.6.2. qRT-PCR: Synthesis of the First cDNA Chain
2.6.3. qRT-PCR Analysis
2.6.4. Quantification of Relative Gene Expression
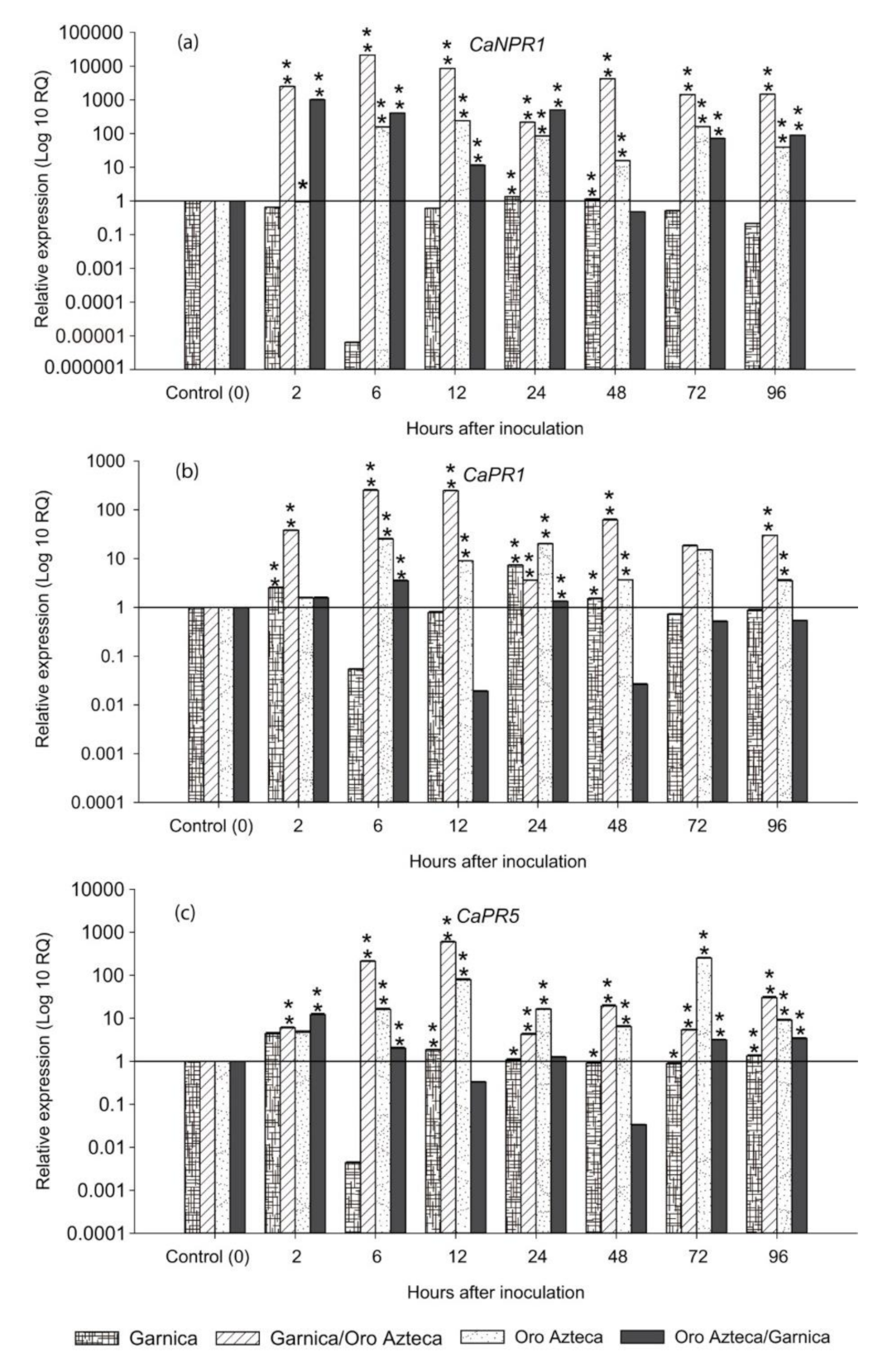
3. Results
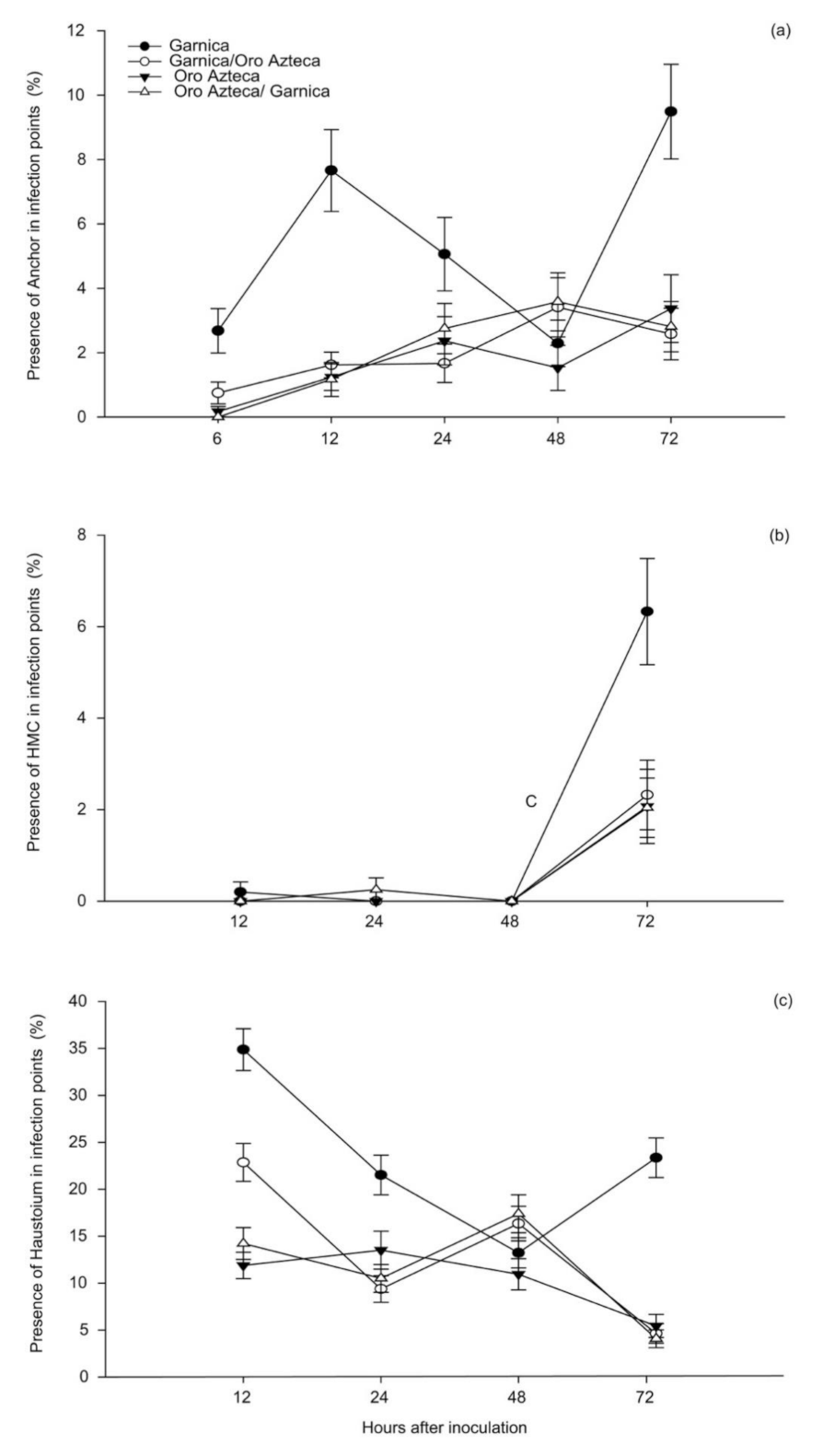
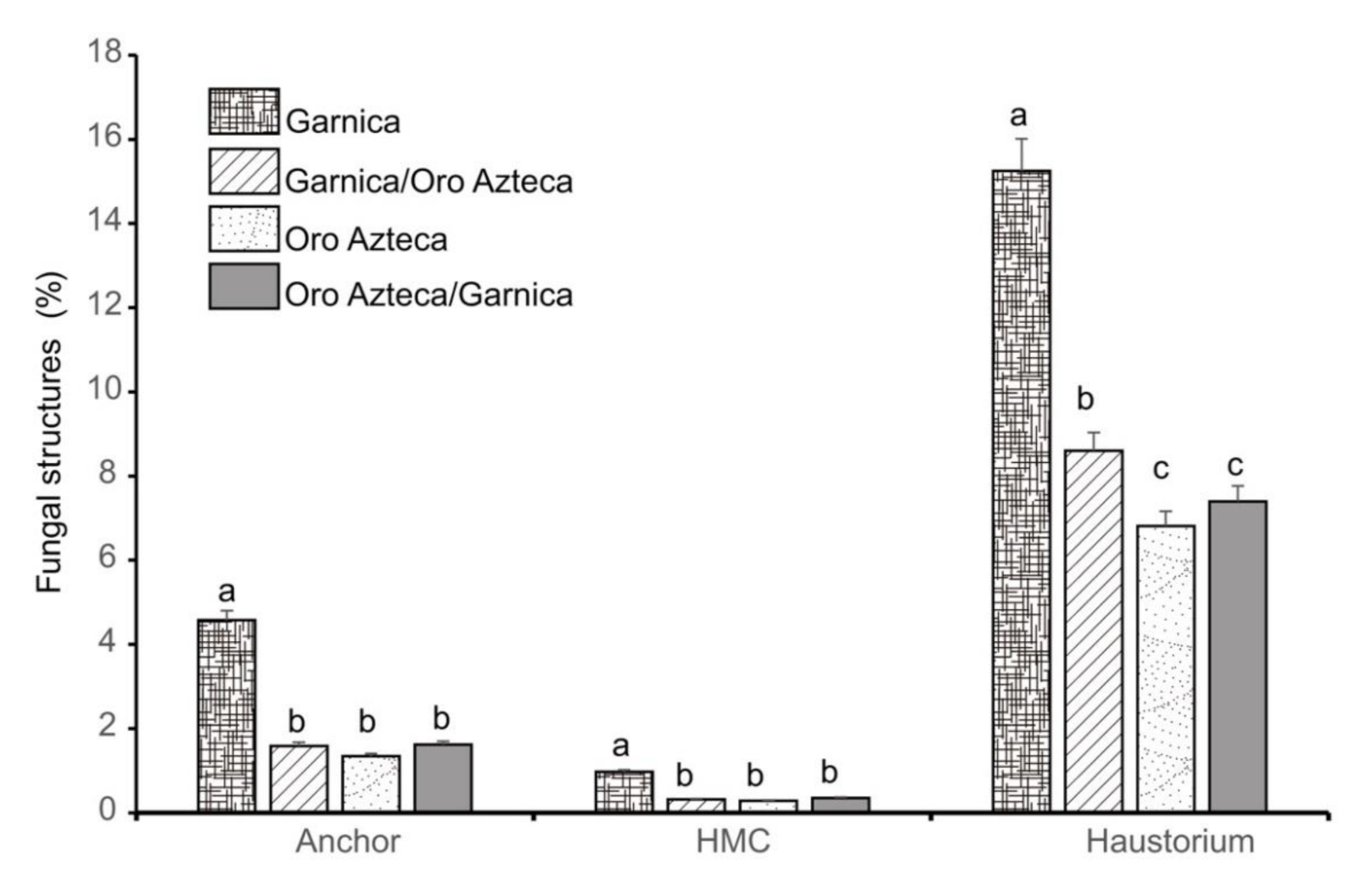
3.1. Development of Hemileia vastatrix in Infected Leaves
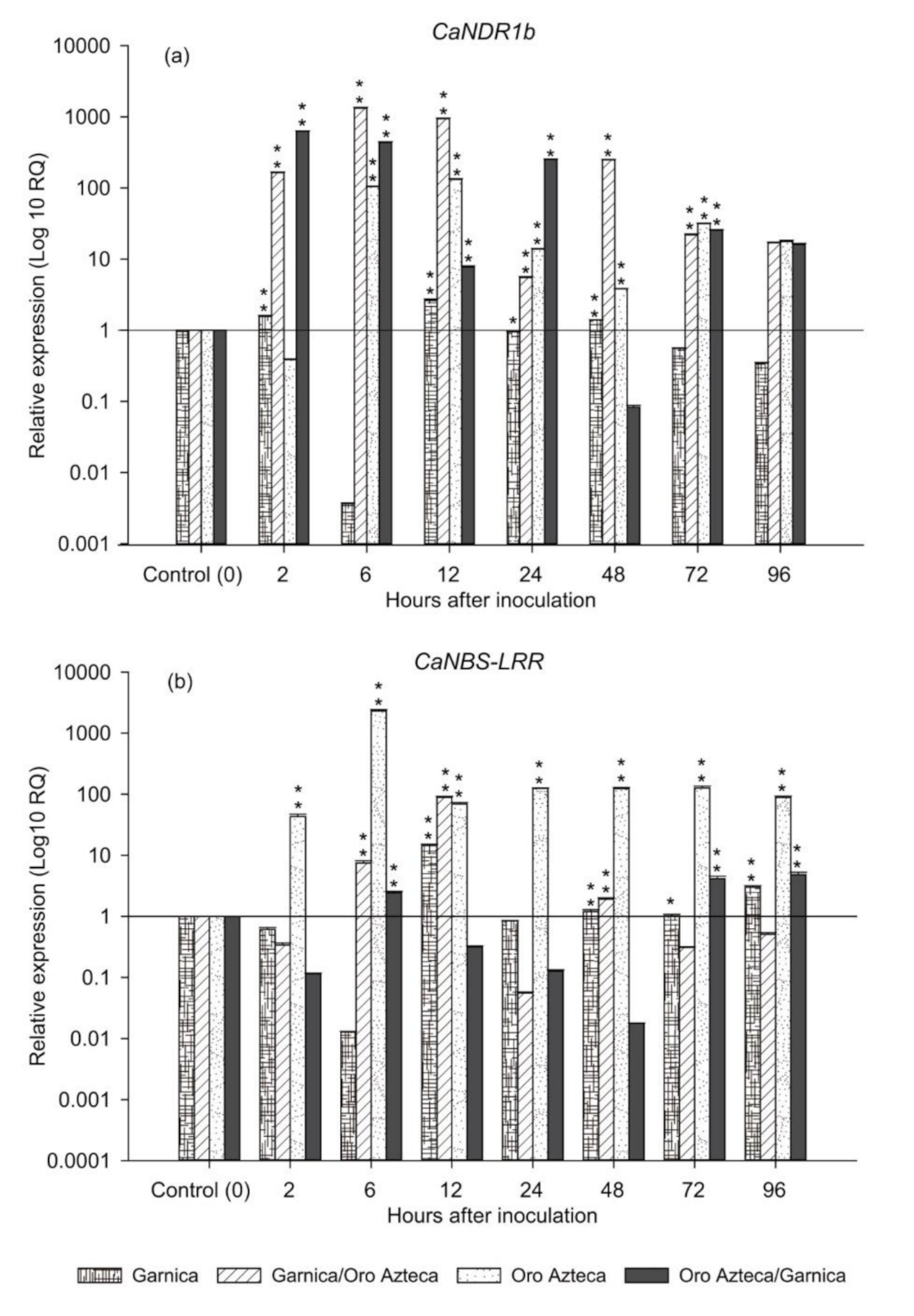
3.2. Expression of CLR Recognition Genes
3.3. Expression of the Salicylic Acid (SA) Pathway-Related Genes
4. Discussion
4.1. Proliferation of Fungal Structures in Ungrafted and Grafted Plants
4.2. Changes in Plant Defense Responses Induced by the Rootstock
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- ICO. Coffee Development Report 2019: Growing for Prosperity Economic Viability as the Catalyst for a Sustainable Coffee Sector; International Coffee Organization: London, UK, 2019; ISBN 978-1527249943. [Google Scholar]
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.J.; Morales, C. The coffee rust crises in Colombia and Central America (2008–2013): Impacts, plausible causes and proposed solutions. Food Secur. 2015, 7, 303–321. [Google Scholar] [CrossRef] [Green Version]
- McCook, S. Coffee is not forever: A global history of the coffee leaf rust. In Ecology and History, 1st ed.; Ohio University Press: Athens, OH, USA, 2019; ISBN 978-0821446843. [Google Scholar]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Zambolim, L. Current status and management of coffee leaf rust in Brazil. Trop. Plant Pathol. 2016, 41, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Berkeley, M.; Broome, C. Hemileia vastatrix . Gard. Chron. 1869, 6, 1157. [Google Scholar]
- Koutouleas, A.; Jørgensen, H.J.L.; Jensen, B.; Lillesø, J.-P.B.; Junge, A.; Ræbild, A. On the hunt for the alternate host of Hemileia vastatrix. Ecol. Evol. 2019, 9, 13619–13631. [Google Scholar] [CrossRef] [Green Version]
- Rayner, R.W. Germination and penetration studies on coffee rust (Hemileia vastatrix B. & Br.). Ann. Appl. Biol. 1961, 49, 497–505. [Google Scholar] [CrossRef]
- Waller, J.M. Coffee rust—Epidemiology and control. Crop Prot. 1982, 1, 385–404. [Google Scholar] [CrossRef]
- Da Costa, G.A.; Lira, J.B.; do Livramento Freitas-Lopes, R.; Lopes, U.P. Tank mix application of copper hydroxide either with cyproconazole or pyraclostrobin fungicides reduced the control of coffee leaf rust. Crop Prot. 2019, 124, 104856. [Google Scholar] [CrossRef]
- De Souza, A.F.; Zambolim, L.; de Jesus Júnior, V.C.; Cecon, P.R. Chemical approaches to manage coffee leaf rust in drip irrigated trees. Australas. Plant Pathol. 2011, 40, 293–300. [Google Scholar] [CrossRef]
- Avelino, J.; Willocquet, L.; Savary, S. Effects of crop management patterns on coffee rust epidemics. Plant Pathol. 2004, 53, 541–547. [Google Scholar] [CrossRef]
- López-Bravo, D.F.; de M. Virginio-Filho, E.; Avelino, J. Shade is conducive to coffee rust as compared to full sun exposure under standardized fruit load conditions. Crop Prot. 2012, 38, 21–29. [Google Scholar] [CrossRef]
- Toniutti, L.; Breitler, J.-C.; Etienne, H.; Campa, C.; Doulbeau, S.; Urban, L.; Lambot, C.; Pinilla, J.-C.H.; Bertrand, B. Influence of environmental conditions and genetic background of arabica coffee (C. arabica L.) on leaf rust (Hemileia vastatrix) pathogenesis. Front. Plant Sci. 2017, 8, 2025. [Google Scholar] [CrossRef] [Green Version]
- Carrión, G.; Rico-Gray, V. Mycoparasites on the coffee rust in Mexico. Fungal Divers. 2002, 11, 49–60. [Google Scholar]
- Haddad, F.; Saraiva, R.M.; Mizubuti, E.S.G.; Romeiro, R.S.; Maffia, L.A. Isolation and selection of Hemileia vastatrix antagonists. Eur. J. Plant Pathol. 2014, 139, 763–772. [Google Scholar] [CrossRef]
- Jackson, D.; Skillman, J.; Vandermeer, J. Indirect biological control of the coffee leaf rust, Hemileia vastatrix, by the entomogenous fungus Lecanicillium lecanii in a complex coffee agroecosystem. Biol. Control 2012, 61, 89–97. [Google Scholar] [CrossRef]
- Daivasikamani, S. Biological control of coffee leaf rust pathogen, Hemileia vastatrix Berkeley and Broome using bacillus subtilis and pseudomonas fluorescens. J. Biopestic. 2009, 2, 94–98. [Google Scholar]
- Haddad, F.; Maffia, L.A.; Mizubuti, E.; Teixeira, H. Biological control of coffee rust by antagonistic bacteria under field conditions in Brazil. Biol. Control 2009, 49, 114–119. [Google Scholar] [CrossRef]
- Bertrand, B.; Guyot, B.; Anthony, F.; Lashermes, P. Impact of the Coffea canephora gene introgression on beverage quality of C. arabica. Theor. Appl. Genet. 2003, 107, 387–394. [Google Scholar] [CrossRef]
- Bertrand, B.; Etienne, H.; Eskes, A. Growth, production, and bean quality of Coffea arabica as affected by interspecific grafting: Consequences for rootstock breeding. HortScience 2001, 36, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Van Der Vossen, H.A.M. The cup quality of disease-resistant cultivars of Arabica coffee (Coffea arabica). Exp. Agric. 2009, 45, 323–332. [Google Scholar] [CrossRef]
- Diniz, I.; Talhinhas, P.; Azinheira, H.G.; Várzea, V.; Medeira, C.; Maia, I.; Petitot, A.-S.; Nicole, M.; Fernandez, D.; do Céu Silva, M. Cellular and molecular analyses of coffee resistance to Hemileia vastatrix and nonhost resistance to Uromyces Vignae in the resistance-donor genotype HDT832/2. Eur. J. Plant Pathol. 2012, 133, 141–157. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Liu, W.; Levi, A. Grafting for disease resistance. HortScience 2008, 43, 1673–1676. [Google Scholar] [CrossRef] [Green Version]
- Louws, F.J.; Rivard, C.L.; Kubota, C. Grafting fruiting vegetables to manage soilborne pathogens, foliar pathogens, arthropods and weeds. Spec. Issue Veg. Grafting 2010, 127, 127–146. [Google Scholar] [CrossRef]
- Guan, W.; Zhao, X.; Hassell, R.; Thies, J. Defense mechanisms involved in disease resistance of grafted vegetables. HortScience 2012, 47, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt, E.E. Plant grafting: New mechanisms, evolutionary implications. Front. Plant Sci. 2014, 5, 727. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Kumar, J.; Kumar, A.; Chaudhury, A.; Singh, S.P. Grafting triggers differential responses between scion and rootstock. PLoS ONE 2015, 10, e0124438. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.J.; Halbrendt, N.; Fazio, G.; Makalowska, I.; Altman, N.; Praul, C.; Maximova, S.N.; Ngugi, H.K.; Crassweller, R.M.; Travis, J.W.; et al. Rootstock-regulated gene expression patterns associated with fire blight resistance in apple. BMC Genom. 2012, 13, 9. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.J.; Rytter, J.; Detwiler, E.A.; Travis, J.W.; McNellis, T.W. Rootstock effects on gene expression patterns in apple tree scions. Plant Mol. Biol. 2003, 53, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Krugner, R.; Ledbetter, C.A. Rootstock effects on almond leaf scorch disease incidence and severity. Plant Dis. 2016, 100, 1617–1621. [Google Scholar] [CrossRef] [PubMed]
- Albert, R.; Künstler, A.; Lantos, F.; Ádám, A.L.; Király, L. Graft-transmissible resistance of cherry pepper (Capsicum annuum Var. Cerasiforme) to powdery mildew (Leveillula taurica) is associated with elevated superoxide accumulation, NADPH oxidase activity and pathogenesis-related gene expression. Acta Physiol. Plant. 2017, 39, 53. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.-T.; Fan, S.-X.; Zhang, X.-C. Effects of rootstocks on the development, disease resistance and quality of Cucumis Sativus L. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 31 July 2008; pp. 161–166. [Google Scholar]
- Bygrave, P.L. Cedrela species are attacked by the tip moth Hypsipyla robusta when grafted on to red cedar Toona ciliata. Aust. For. 1998, 61, 45–47. [Google Scholar] [CrossRef]
- Perez, J.; Eigenbrode, S.D.; Hilje, L.; Tripepi, R.R.; Aguilar, M.E.; Mesen, F. Use of grafting to prevent Hypsipyla grandella (Zeller) (Lepidoptera: Pyralidae) damage to new world meliaceae species. Neotrop. Entomol. 2010, 39, 618–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agut, B.; Gamir, J.; Jaques, J.A.; Flors, V. Systemic resistance in citrus to Tetranychus Urticae induced by conspecifics is transmitted by grafting and mediated by mobile amino acids. J. Exp. Bot. 2016, 67, 5711–5723. [Google Scholar] [CrossRef] [Green Version]
- Stack, S.C.; Sadof, C.S.; Ginzel, M.D. Effects of grafting on host plant resistance in ash (Fraxinus spp.) to emerald ash borer (Agrilus planipennis Fairmaire). Agric. For. Entomol. 2019, 21, 180–189. [Google Scholar] [CrossRef]
- Koepke, T.; Dhingra, A. Rootstock scion somatogenetic interactions in perennial composite plants. Plant Cell Rep. 2013, 32, 1321–1337. [Google Scholar] [CrossRef]
- Albrecht, U.; Tripathi, I.; Kim, H.; Bowman, K.D. Rootstock effects on metabolite composition in leaves and roots of young navel orange (Citrus sinensis L. Osbeck) and pummelo (C. grandis L. Osbeck) trees. Trees 2019, 33, 243–265. [Google Scholar] [CrossRef]
- Kanehira, A.; Yamada, K.; Iwaya, T.; Tsuwamoto, R.; Kasai, A.; Nakazono, M.; Harada, T. Apple phloem cells contain some MRNAs transported over long distances. Tree Genet. Genomes 2010, 6, 635–642. [Google Scholar] [CrossRef]
- Song, G.; Sink, K.C.; Walworth, A.E.; Cook, M.A.; Allison, R.F.; Lang, G.A. Engineering cherry rootstocks with resistance to prunus necrotic ring spot virus through RNAi-mediated silencing. Plant Biotechnol. J. 2013, 11, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Kollwig, G.; Stecyk, E.; Apelt, F.; Dirks, R.; Kragler, F. Graft-transmissible movement of inverted-repeat-induced SiRNA signals into flowers. Plant J. 2014, 80, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Chitarra, W.; Perrone, I.; Avanzato, C.G.; Minio, A.; Boccacci, P.; Santini, D.; Gilardi, G.; Siciliano, I.; Gullino, M.L.; Delledonne, M.; et al. Grapevine grafting: Scion transcript profiling and defense-related metabolites induced by rootstocks. Front. Plant Sci. 2017, 8, 654. [Google Scholar] [CrossRef] [PubMed]
- Cookson, S.J.; Ollat, N. Grafting with rootstocks induces extensive transcriptional re-programming in the shoot apical meristem of grapevine. BMC Plant Biol. 2013, 13, 147. [Google Scholar] [CrossRef]
- Huang, Y.; Si, Y.; Dane, F. Impact of grafting on cold responsive gene expression in satsuma mandarin (Citrus unshiu). Euphytica 2011, 177, 25–32. [Google Scholar] [CrossRef]
- Zhang, Z.-J.; Wang, Y.-M.; Long, L.-K.; Lin, Y.; Pang, J.-S.; Liu, B. Tomato rootstock effects on gene expression patterns in eggplant scions. Russ. J. Plant Physiol. 2008, 55, 93–100. [Google Scholar] [CrossRef]
- Campos, V.P.; Silva, J.R.C. Management of Meloidogyne spp. in coffee plantations. In Plant-Parasitic Nematodes of Coffee; Souza, R.M., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 149–164. ISBN 978-1-4020-8720-2. [Google Scholar]
- Wangai, K.J.; Nzesya, M.J.; Maina, M.W.; Peter, W.M.; Elijah, G.K. Reaction of selected coffee germplasm to root-knot nematodes in Kenya. J. Nat. Sci. Res. 2014, 4, 68–75. [Google Scholar]
- Villain, L.; Salgado, S.M.L.; Trinh, P.Q. Nematode parasites of coffee and cocoa. In Plant Parasitic Nematodes in Subtroptropical and Tropical Agriculture; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CABI Pulishing: Wallingford, UK, 2018; pp. 536–583. [Google Scholar]
- Pham, T.T.; Giang, B.L.; Nguyen, N.H.; Dong Yen, P.N.; Minh Hoang, V.D.; Lien Ha, B.T.; Le, N.T.T. Combination of mycorrhizal symbiosis and root grafting effectively controls nematode in replanted coffee soil. Plants 2020, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Villain, L.; Molina, A.; Sierra, S.; Decazy, B.; Anzueto, F.; Guyot, B.; Perriot, J.-J.; Sarah, J.-L. Evaluation of grafting and nematicide treatments for the management of a root-lesion nematode, Pratylenchus sp. in Coffee arabica L. Plantations in Guatemala. In Proceedings of the Dix-neuvième colloque scientifique international sur le café—Actes, Trieste, Italy, 14–18 May 2001. [Google Scholar]
- González, A.L.; Lopez, A.M.; Gaytán, O.R.T.; Ramos, V.M. Cup quality attributes of catimors as affected by size and shape of coffee bean (Coffea arabica L.). Int. J. Food Prop. 2019, 22, 758–767. [Google Scholar] [CrossRef] [Green Version]
- Rivera Fernández, A.; Villareal Ruíz, C. La variedad garnica de Coffea arabica L. In El Agro Veracruzano 2015; del Angel Pérez, A.L., Ed.; Academia Veracruzana de Ciencias Agrícolas, Pecuarias, Forestales, Acuícolas y Pesqueras, A.C.: Boca del Río, Mexico, 2015; Volume II, pp. 46–57. ISBN 978-607-96939-2-3. [Google Scholar]
- Montagnon, C.; Marraccini, P.; Bertrand, B. Breeding for coffee quality. In Specialty Coffee: Managing Quality; Oberthür, T., Läderach, P., Pohlan, H.A.J., Cock, J., Eds.; International Plant Nutrition Institute: Penang, Malaysia, 2012; pp. 89–118. ISBN 978-983-44503-1-1. [Google Scholar]
- Silva, M.C.; Nicole, M.; Guerra-GuimarÃes, L.; Rodrigues, C.J. Hypersensitive cell death and post-haustorial defence responses arrest the orange rust (Hemileia vastatrix) growth in resistant coffee leaves. Physiol. Mol. Plant Pathol. 2002, 60, 169–183. [Google Scholar] [CrossRef]
- Cohen, L.; Eyal, Z. The histology of processes associated with the infection of resistant and susceptible wheat cultivars with Septoria Tritici. Plant Pathol. 1993, 42, 737–743. [Google Scholar] [CrossRef]
- Pratt, C.; Wetmore, R.H. A paraffin method for refractory plant materials. Stain Technol. 1951, 26, 251–253. [Google Scholar] [CrossRef]
- Ganesh, D.; Petitot, A.-S.; Silva, M.C.; Alary, R.; Lecouls, A.-C.; Fernandez, D. Monitoring of the early molecular resistance responses of coffee (Coffea arabica L.) to the rust fungus (Hemileia vastatrix) using real-time quantitative RT-PCR. Plant Sci. 2006, 170, 1045–1051. [Google Scholar] [CrossRef]
- Ramiro, D.A.; Escoute, J.; Petitot, A.-S.; Nicole, M.; Maluf, M.P.; Fernandez, D. Biphasic haustorial differentiation of coffee rust (Hemileia vastatrix Race II) associated with defence responses in resistant and susceptible coffee cultivars. Plant Pathol. 2009, 58, 944–955. [Google Scholar] [CrossRef]
- Diola, V.; Brito, G.G.; Caixeta, E.T.; Pereira, L.F.P.; Loureiro, M.E. A new set of differentially expressed signaling genes is early expressed in coffee leaf rust race II incompatible interaction. Funct. Integr. Genom. 2013, 13, 379–389. [Google Scholar] [CrossRef]
- Barsalobres-Cavallari, C.F.; Severino, F.E.; Maluf, M.P.; Maia, I.G. Identification of suitable internal control genes for expression studies in Coffea arabica under different experimental conditions. BMC Mol. Biol. 2009, 10, 1. [Google Scholar] [CrossRef] [Green Version]
- Cruz, F.; Kalaoun, S.; Nobile, P.; Colombo, C.; Almeida, J.; Barros, L.M.G.; Romano, E.; Grossi-de-Sá, M.F.; Vaslin, M.; Alves-Ferreira, M. Evaluation of coffee reference genes for relative expression studies by quantitative real-time RT-PCR. Mol. Breed. 2009, 23, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, A.; Loureiro, A.; Batista, D.; Monteiro, F.; Várzea, V.; Pais, M.S.; Gichuru, E.K.; Silva, M.C. Validation of reference genes for normalization of QPCR gene expression data from Coffea spp. Hypocotyls inoculated with Colletotrichum kahawae. BMC Res. Notes 2013, 6, 388. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Tsai, S.M.; Caldas, D.G.G. Validation of reference genes for RT-QPCR normalization in common bean during biotic and abiotic stresses. Plant Cell Rep. 2012, 31, 827–838. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods (San Diego, Calif.) 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Walker, N.J. A technique whose time has come. Science 2002, 296, 557–559. [Google Scholar] [CrossRef]
- Couttolenc-Brenis, E.; Carrión, G.L.; Villain, L.; Ortega-Escalona, F.; Ramírez-Martínez, D.; Mata-Rosas, M.; Méndez-Bravo, A. Prehaustorial local resistance to coffee leaf rust in a mexican cultivar involves expression of Salicylic acid-responsive genes. PeerJ 2020, 8, e8345. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Guimarães, L.L.; Tenente, R.E.R.; Pinheiro, C.C.; Chaves, I.I.; Silva, M.D.C.M.L.M.; Cardoso, F.F.; Planchon, S.S.; De Barros, D.R.D.; Renaut, J.J.; Ricardo, C.P.C. Proteomic analysis of apoplastic fluid of Coffea arabica leaves highlights novel biomarkers for resistance against Hemileia vastatrix. Front. Plant Sci. 2015, 6, 478. [Google Scholar] [CrossRef] [Green Version]
- Martins, E.M.F.; Moraes, W.B.C. Development of Hemileia vastatrix in coffee plants with genetic or induced resistance. J. Phytopathol. 1996, 144, 519–526. [Google Scholar] [CrossRef]
- Kousik, C.S.; Mandal, M.; Hassell, R. Powdery mildew resistant rootstocks that impart tolerance to grafted susceptible watermelon scion seedlings. Plant Dis. 2018, 102, 1290–1298. [Google Scholar] [CrossRef] [Green Version]
- Florez, J.C.; Mofatto, L.S.; do Livramento Freitas-Lopes, R.; Ferreira, S.S.; Zambolim, E.M.; Carazzolle, M.F.; Zambolim, L.; Caixeta, E.T. High throughput transcriptome analysis of coffee reveals prehaustorial resistance in response to Hemileia vastatrix infection. Plant Mol. Biol. 2017, 95, 607–623. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.D.M.; Varanda, C.M.R.; Félix, M.R.F. Induced resistance during the interaction pathogen x plant and the use of resistance inducers. Phytochem. Lett. 2016, 15, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Cacas, J.-L.; Petitot, A.-S.; Bernier, L.; Estevan, J.; Conejero, G.; Mongrand, S.; Fernandez, D. Identification and characterization of the non-race specific disease resistance 1 (NDR1) orthologous protein in coffee. BMC Plant Biol. 2011, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, D.; Santos, P.; Agostini, C.; Bon, M.-C.; Petitot, A.-S.; Silva, M.D.C.; Guerra-Guimarães, L.; I Ribeiro-Barros, A.; Argout, X.; Nicole, M. Coffee (Coffea arabica L.) genes early expressed during infection by the rust fungus (Hemileia vastatrix). Mol. Plant Pathol. 2004, 5, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Yang, Y.; Zhong, C.; Notaguchi, M.; Yu, W. A susceptible scion reduces rootstock tolerance to Ralstonia solanacearum in grafted eggplant. Horticulturae 2019, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Cohn, J.; Sessa, G.; Martin, G.B. Innate immunity in plants. Curr. Opin. Immunol. 2001, 13, 55–62. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyajyan, A.; Devejyan, H.; Haykazyan, V.; Avetisyan, G.; Khanoyan, D. Molecular mechanisms and mediators of the immune response in plants. J. Plant Sci. 2014, 2, 23. [Google Scholar] [CrossRef]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Magnin, N.; Trossat-Magnin, C.; Ollat, N. Heterografting with nonself rootstocks induces genes involved in stress responses at the graft interface when compared with autografted controls. J. Exp. Bot. 2014, 65, 2473–2481. [Google Scholar] [CrossRef] [Green Version]
- Sá, M.; Ferreira, J.P.; Queiroz, V.T.; Vilas-Boas, L.; Silva, M.C.; Almeida, M.H.; Guerra-Guimarães, L.; Bronze, M.R. A liquid chromatography/electrospray ionisation tandem mass spectrometry method for the simultaneous quantification of salicylic, jasmonic and abscisic acids in Coffea arabica leaves. J. Sci. Food Agric. 2014, 94, 529–536. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.A. Systemic acquired resistance and salicylic acid: Past, present, and future. Mol. Plant-Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, D.; Khurana, J.P. Role of pathogenesis-related (PR) proteins in plant defense mechanism. In Molecular Aspects of Plant-Pathogen Interaction; Singh, A., Singh, I.K., Eds.; Springer: Singapore, 2018; pp. 265–281. ISBN 978-9811073717. [Google Scholar]
- Xia, Y. Chapter eleven—Correlation and association analyses in microbiome study integrating multiomics in health and disease. In Progress in Molecular Biology and Translational Science; Sun, J., Ed.; The Microbiome in Health and Disease; Academic Press: Cambridge, MA, USA, 2020; Volume 171, pp. 309–491. [Google Scholar]
- Couttolenc-Brenis, E.; Mata-Rosas, M.; Méndez-Bravo, A.; Villain, L.; Carrion, G. Cambios en la severidad de la infección de roya en planta injertada de café arábica. In Avances en Investigación Agrícola, Pecuaria, Forestal, Acuícola, Pesquería, Desarrollo Rural, Transferencia de Tecnología, Biotecnología, Ambiente, Recursos Naturales y Cambio Climático; Vinay Vadillo, J.C., Esqueda Esquivel, V.A., Tosquy Valle, O.H., Rios Utrera, A., Zerina Lezama, R., Del Angel Perez, A.L., Vázquez Hernández, M.V., Perdomo Montes, C., Eds.; INIFAP, CP, UACH, INAPESCA, UV, TecNM: Medellín, Mexico, 2019; Volume 3, pp. 549–557. ISSN 2594-147X. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Gene Type |
|---|---|---|
| CaNBS-LRR | F5′-CCAAAAACTTTgggTTggTg-3′ R5′-TCCATTgCATTCTCATCTg-3′ | Plant defense involved in pathogen recognition |
| CaNDR1b | F5′-CTTACAgggCggTgTCAAAT-3′ R5′-TACCACTAgCCCAggACAgC-3′ | Plant defense, hypersensitive response |
| CaNPR1 | F5′-gACgCTgCAgTgAAgAAAC-3′ R5′-TgATAgCTTCCCAggCATCT-3′ | Plant defense involved in the salicylic acid pathway |
| CaPR1 | F 5′-CaggAATgCgggCATTATAC-3′ R 5′-CAATCgCATgggTTTgATAA-3′ | Plant defense involved in the synthesis of salicylic acid |
| CaPR5 | F 5′-CtgCCTgAgTTgCAgCAATA-3′ R 5′-TTTCCCTTgTTgATggCTTC-3′ | Plant defense involved in the salicylic acid pathway |
| CaUbiE2 | F5′-CCATTTAAACCCCCAAAggT-3′ R5′-ggTCCAgCTTCgAgCAgTAg-3′ | C. arabica constitutive |
| CaGAPH | F5′- gCAgCACTTCATggTTCTgA-3′ R5′-TTTCCACATTTCAgCCCTTC-3′ | C. arabica constitutive |
| Hai | Pr > F | Garnica | Garnica/Oro Azteca | Oro Azteca | Oro Azteca/Garnica | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| µ | S.E. | µ | S.E. | µ | S.E. | µ | S.E. | |||
| PH | 2 | 0.0910 | 0.81 a | 0.48 | 0 a | 0.00 | 0 a | 0.00 | 0.26 a | 0.26 |
| 6 | <0.0001 | 41.64 a | 2.42 | 18.25 b | 1.55 | 5.78 c | 0.88 | 3.57 c | 0.78 | |
| Anchor | 6 | <0.0001 | 2.68 a | 0.69 | 0.75 b | 0.34 | 0.17 b | 0.16 | 0 b | 0 |
| 12 | <0.0001 | 7.66 a | 1.27 | 1.62 b | 0.4 | 1.24 b | 0.41 | 1.17 b | 0.53 | |
| 24 | 0.0018 | 5.06 a | 1.14 | 1.67 b | 0.6 | 2.37 b | 0.75 | 2.75 ab | 0.78 | |
| 48 | 0.3212 | 2.29 a | 0.72 | 3.41 a | 0.92 | 1.53 a | 0.7 | 3.58 a | 0.9 | |
| 72 | <0.0001 | 9.49 a | 1.47 | 2.58 b | 0.8 | 3.37 b | 1.05 | 2.81 b | 0.78 | |
| HMC | 12 | 0.0940 | 0.20 a | 0.22 | 0 a | 0 | 0 a | 0 | 0 a | 0 |
| 24 | 0.3329 | 0 a | 0 | 0 a | 0 | 0.25 a | 0.26 | 0 a | 0 | |
| 48 | -- | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 72 | 0.0003 | 6.33 a | 1.16 | 2.32 b | 0.76 | 2.07 b | 0.81 | 2.04 b | 0.65 | |
| Haustoria | 12 | <0.0001 | 34.88 a | 2.23 | 22.87 b | 2.02 | 11.88 c | 1.40 | 14.22 c | 1.70 |
| 24 | <0.0001 | 21.52 a | 2.11 | 9.38 b | 1.41 | 13.51 b | 2.02 | 10.50 b | 1.48 | |
| 48 | 0.0780 | 13.23 a | 1.61 | 16.34 a | 1.85 | 10.94 a | 1.65 | 17.39 a | 2.02 | |
| 72 | <0.0001 | 23.36 a | 2.11 | 4.64 b | 1.04 | 5.44 b | 1.24 | 4.08 b | 0.95 | |
| Hai | CaNDRb1 | CaNBS-LRR | ||||
|---|---|---|---|---|---|---|
| χ2 | DF | Pr > χ2 | χ2 | DF | Pr > χ2 | |
| 2 | 12.20 | 3 | 0.0067 | 14.1176 | 3 | 0.0027 |
| 6 | 14.1176 | 3 | 0.0027 | 14.1176 | 3 | 0.0027 |
| 12 | 14.1176 | 3 | 0.0027 | 13.15 | 3 | 0.0043 |
| 24 | 13.15 | 3 | 0.0043 | 13.15 | 3 | 0.0043 |
| 48 | 12.20 | 3 | 0.0067 | 14.1176 | 3 | 0.0027 |
| 72 | 14.1176 | 3 | 0.0027 | 13.15 | 3 | 0.0043 |
| 96 | 10.0792 | 3 | 0.0179 | 14.1176 | 3 | 0.0027 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Couttolenc-Brenis, E.; Carrión, G.; Villain, L.; Ortega-Escalona, F.; Mata-Rosas, M.; Méndez-Bravo, A. Defense Response to Hemileia vastatrix in Susceptible Grafts onto Resistant Rootstock of Coffea arabica L. Agronomy 2021, 11, 1621. https://doi.org/10.3390/agronomy11081621
Couttolenc-Brenis E, Carrión G, Villain L, Ortega-Escalona F, Mata-Rosas M, Méndez-Bravo A. Defense Response to Hemileia vastatrix in Susceptible Grafts onto Resistant Rootstock of Coffea arabica L. Agronomy. 2021; 11(8):1621. https://doi.org/10.3390/agronomy11081621
Chicago/Turabian StyleCouttolenc-Brenis, Edgar, Gloria Carrión, Luc Villain, Fernando Ortega-Escalona, Martín Mata-Rosas, and Alfonso Méndez-Bravo. 2021. "Defense Response to Hemileia vastatrix in Susceptible Grafts onto Resistant Rootstock of Coffea arabica L." Agronomy 11, no. 8: 1621. https://doi.org/10.3390/agronomy11081621