
LED Light Pre-Treatment Improves Pre-Basic Seed Potato (Solanum tuberosum L. cv. Golden King) Production in the Aeroponic System


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plantlet Production and Growth Conditions
2.2. Plant Factory and Light-Emitting Diode (LED) Settings
2.3. Aeroponic System
2.4. Measurement of Plant Growth Characteristics and Seed Tuber Yield
2.5. Analysis of Photosynthetic Pigments of Potato Plants
2.6. Determination of Total Soluble Carbohydrate (TSC) and Total Soluble Sugar (TSS) Content
2.7. Determination of Tuber Yield
2.8. Statistical Analysis
3. Results
3.1. Effect of LED Light on Plant Morphological Characteristics
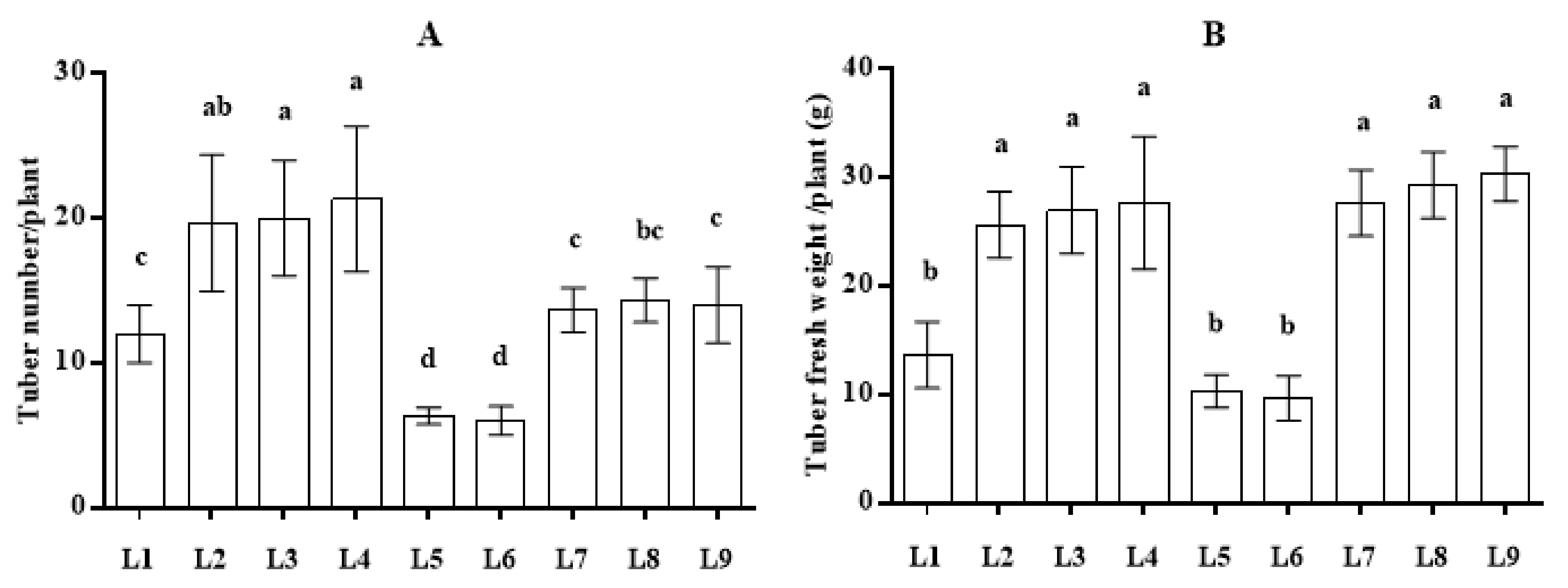
3.2. Effect of LED Light on Tuber Yield and Grading
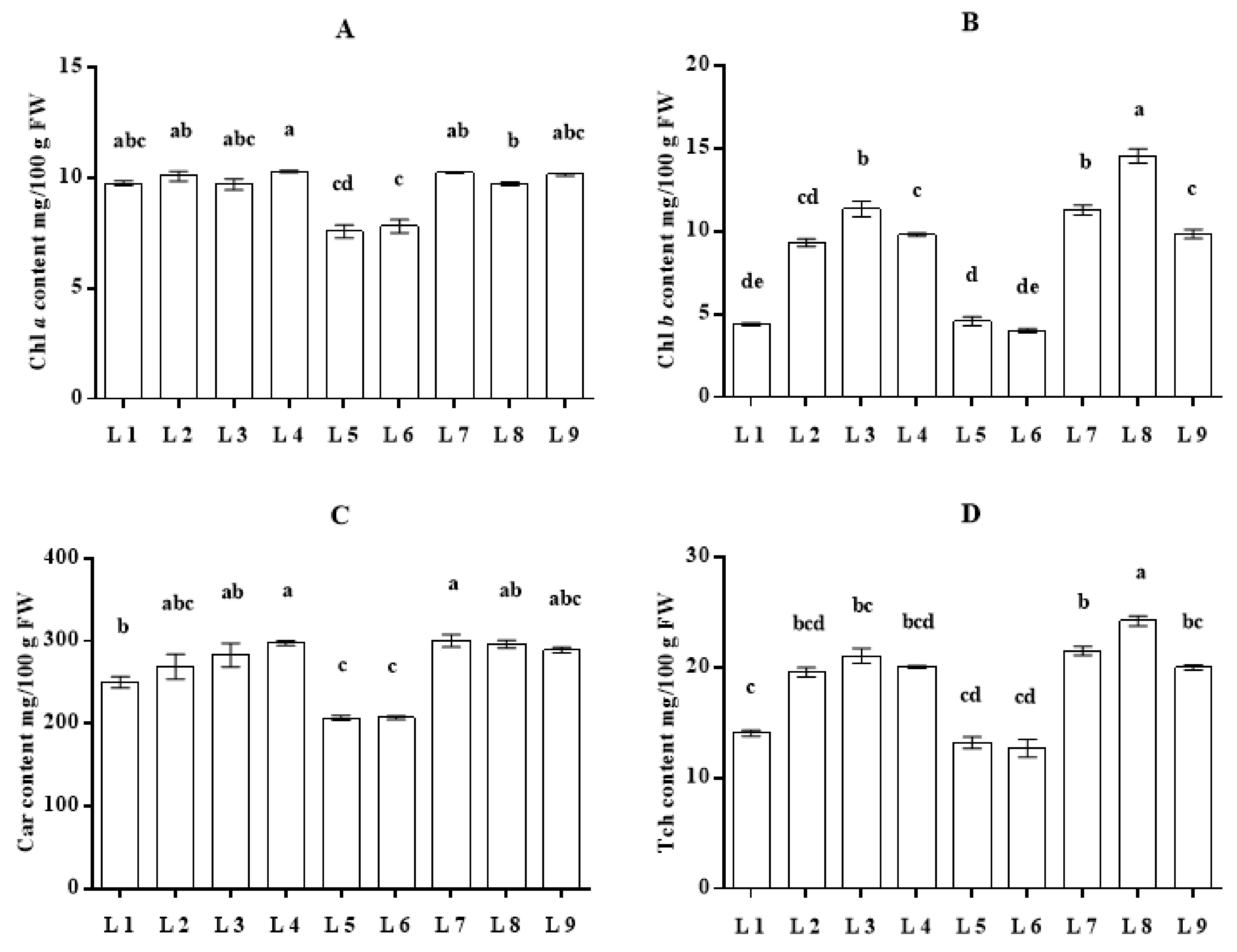
3.3. Effect of LED Light on Photosynthetic Pigments
3.4. Effect of LED Light on Total Soluble Carbohydrates (TSC) and Total Soluble Sugar Content (TSS)
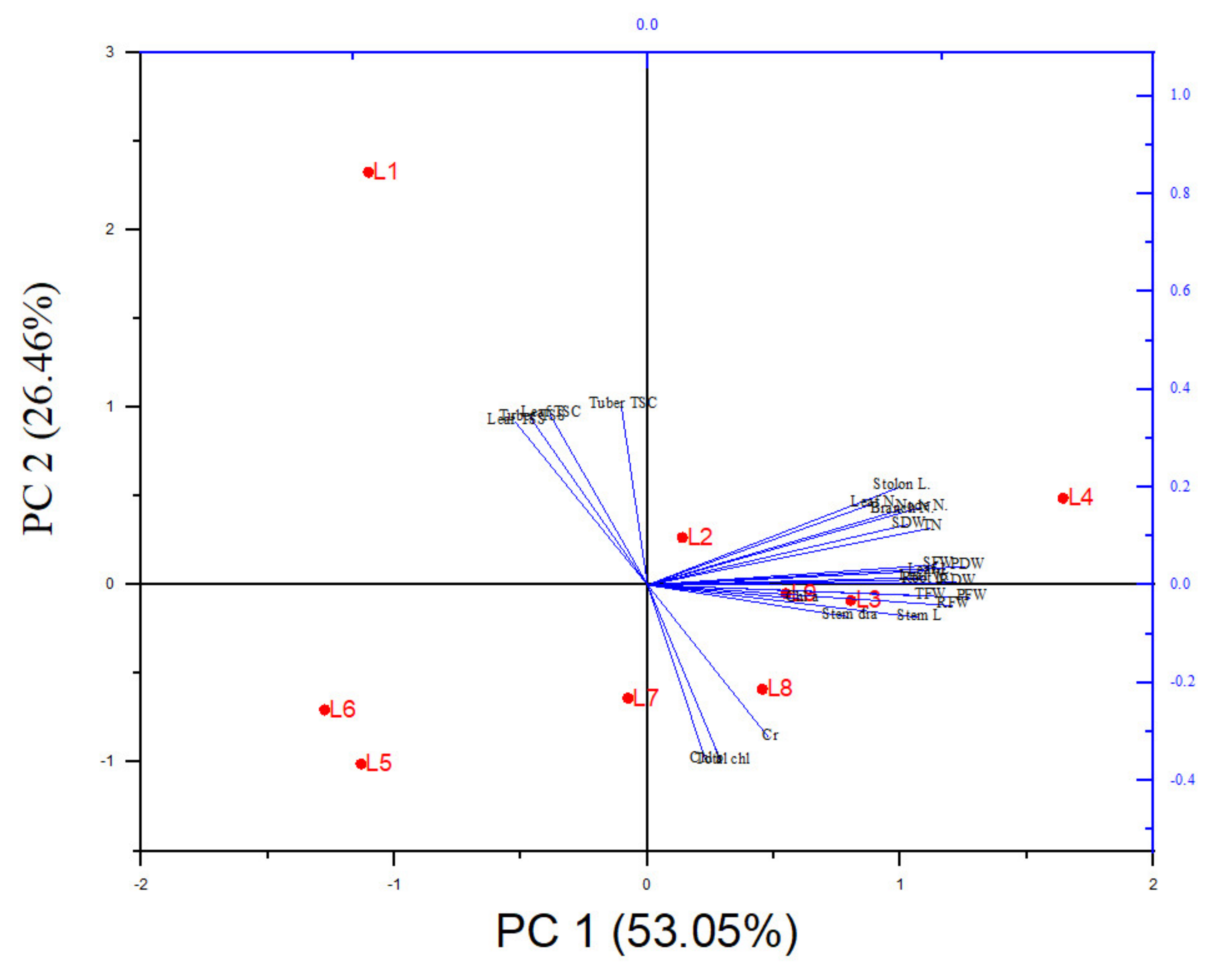
3.5. The PCA Analysis Revealed the Connection between Treatments and Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halterman, D.; Guenthner, J.; Collinge, S.; Butler, N.; Douches, D. Biotech potatoes in the 21st century: 20 years since the first biotech potato. Am. J. Potato Res. 2016, 93, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Islam, J.; Choi, S.P.; Azad, O.K.; Kim, J.W.; Lim, Y.-S. Evaluation of Tuber Yield and Marketable Quality of Newly Developed Thirty-Two Potato Varieties Grown in Three Different Ecological Zones in South Korea. Agriculture 2020, 10, 327. [Google Scholar] [CrossRef]
- Rolston, M.P.; Gomez, J.S. Methods for pre-basic seed production of herbage cultivars. N. Z. J. Exp. Agric. 1986, 14, 165–171. [Google Scholar] [CrossRef]
- Buckseth, T.; Sharma, A.K.; Pandey, K.K.; Singh, B.P.; Muthuraj, R. Methods of pre-basic seed potato production with special reference to aeroponics—A review. Sci. Hortic. 2016, 204, 79–87. [Google Scholar] [CrossRef]
- Navarrete Ortega, J.I. Evaluación de Dos Métodos de Micropropagacion para la Producción de Semilla Categoría Pre Básica de dos Variedades de Papa (Solanum tuberosum) Bajo Condiciones de Invernadero. Ph.D. Thesis, Central University of Ecuador, Quito, Ecuador, 2004. [Google Scholar]
- Lommen, W.J.M. The canon of potato science: 27. Hydroponics. Potato Res. 2007, 50, 315. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.H.; Azad, M.O.K.; Islam, M.J.; Rana, M.S.; Li, K.; Lim, Y.S. Production of Potato (Solanum tuberosum L.) Seed Tuber under Artificial LED Light Irradiation in Plant Factory. Plants 2021, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Folta, K.M. Breeding new varieties for controlled environments. Plant Biol. 2019, 21, 6–12. [Google Scholar] [CrossRef]
- Islam, M.J.; Hassan, M.K.; Sarker, S.R.; Rahman, A.B.; Fakir, M.S.A. Light and temperature effects on sprout yield and its proximate composition and vitamin C content in lignosus and mung beans. J. Bangladesh Agric. Univ. 2017, 15, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.; Rahman, M.H.; Cheong, E.J.; Lim, J.-D.; Lim, Y.-S. Cannabinoids Accumulation in Hemp (Cannabis sativa L.) Plants under LED Light Spectra and Their Discrete Role as a Stress Marker. Biology 2021, 10, 710. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Matthes, M.C.; Napier, J.A.; Pickett, J.A. Stressful “memories” of plants: Evidence and possible mechanisms. Plant Sci. 2007, 173, 603–608. [Google Scholar] [CrossRef]
- Galis, I.; Gaquerel, E.; Pandey, S.P.; Baldwin, I.T. Molecular mechanisms underlying plant memory in JA-mediated defence responses. Plant Cell Environ. 2009, 32, 617–627. [Google Scholar] [CrossRef]
- Ding, Y.; Fromm, M.; Avramova, Z. Multiple exposures to drought’train’transcriptional responses in Arabidopsis. Nat. Commun. 2012, 3, 1–9. [Google Scholar] [CrossRef]
- Skirycz, A.; Inzé, D. More from less: Plant growth under limited water. Curr. Opin. Biotechnol. 2010, 21, 197–203. [Google Scholar] [CrossRef]
- Terashima, I.; Fujita, T.; Inoue, T.; Chow, W.S.; Oguchi, R. Green light drives leaf photosynthesis more efficiently than red light in strong white light: Revisiting the enigmatic question of why leaves are green. Plant Cell Physiol. 2009, 50, 684–697. [Google Scholar] [CrossRef] [Green Version]
- Hanyu, H.; Shoji, K. Acceleration of growth in spinach by short-term exposure to red and blue light at the beginning and at the end of the daily dark period. In Proceedings of the IV International ISHS Symposium on Artificial Lighting, Quebec, QC, Canada, 7–9 November 2000; Volume 580, pp. 145–150. [Google Scholar]
- Smith, H. Light quality, photoperception, and plant strategy. Annu. Rev. Plant Physiol. 1982, 33, 481–518. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Lefsrud, M.G.; Kopsell, D.E.; Curran-Celentano, J. Irradiance Levels Affect Growth Parameters and Carotenoid Pigments in Kale and Spinach Grown in a Controlled Environment. Physiol. Plant. 2006, 127, 624–631. [Google Scholar]
- Azad, M.O.K.; Kjaer, K.H.; Adnan, M.; Naznin, M.T.; Lim, J.D.; Sung, I.J.; Park, C.H.; Lim, Y.S. The Evaluation of Growth Performance, Photosynthetic Capacity, and Primary and Secondary Metabolite Content of Leaf Lettuce Grown under Limited Irradiation of Blue and Red LED Light in an Urban Plant Factory. Agriculture 2020, 10, 28. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Hahn, E.-J.; Heo, J.-W.; Paek, K.-Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Zhang, T.; Maruhnich, S.A.; Folta, K.M. Green light induces shade avoidance symptoms. Plant Physiol. 2011, 157, 1528–1536. [Google Scholar] [CrossRef] [Green Version]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Gao, C.; El-Sawah, A.M.; Ali, D.F.I.; Hamoud, Y.A.; Shaghaleh, H.; Sheteiwy, M.S. The integration of bio and organic fertilizers improve plant growth, grain yield, quality and metabolism of hybrid maize (Zea mays L.). Agronomy 2020, 10, 319. [Google Scholar] [CrossRef] [Green Version]
- Siyami, R.; Mirshekari, B.; Farahvash, F.; Rashidi, V.; Tarinejad, A. The effect of physical priming of seed on traits and yield of corn (Zea mays L.) under water deficit conditions in Iran. Appl. Ecol. Environ. Res. 2018, 16, 617–627. [Google Scholar] [CrossRef]
- Khan, A.; Khalil, S.K.; Khan, A.Z.; Marwat, K.B.; Afzal, A. The role of seed priming in semi-arid area for mung bean phenology and yield. Pakistan J. Bot. 2008, 40, 2471–2480. [Google Scholar]
- Hu, L.; Yu, J.; Liao, W.; Zhang, G.; Xie, J.; Lv, J.; Xiao, X.; Yang, B.; Zhou, R.; Bu, R. Moderate ammonium: Nitrate alleviates low light intensity stress in mini Chinese cabbage seedling by regulating root architecture and photosynthesis. Sci. Hortic. 2015, 186, 143–153. [Google Scholar] [CrossRef]
- Gao, J.; Shi, J.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Grain yield and root characteristics of summer maize (Zea mays L.) under shade stress conditions. J. Agron. Crop Sci. 2017, 203, 562–573. [Google Scholar] [CrossRef]
- Silva, T.D.; Batista, D.S.; Fortini, E.A.; de Castro, K.M.; Felipe, S.H.S.; Fernandes, A.M.; de Jesus Sousa, R.M.; Chagas, K.; da Silva, J.V.S.; de Freitas Correia, L.N. Blue and red light affects morphogenesis and 20-hydroxyecdisone content of in vitro Pfaffia glomerata accessions. J. Photochem. Photobiol. B Biol. 2020, 203, 111761. [Google Scholar] [CrossRef]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.K.; Rahman, M.H.; Rana, M.S.; Kang, C.-W.; Lim, J.-D.; Lim, Y.-S. Comparative Growth, Photosynthetic Pigments, and Osmolytes Analysis of Hemp (Cannabis sativa L.) Seedlings under an Aeroponics System with Different LED Light Sources. Horticulturae 2021, 7, 239. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Islam, M.J.; Kim, J.W.; Begum, M.K.; Sohel, M.A.T.; Lim, Y.-S. Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants 2020, 9, 1511. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Nhut, D.T.; Don, N.T.; Tanaka, M. Light-emitting diodes as an effective lighting source for in vitro banana culture. In Protocols for Micropropagation of Woody Trees and Fruits; Springer: Berlin, Germany, 2007; pp. 527–541. [Google Scholar]
- Li, H.; Tang, C.; Xu, Z. The effects of different light qualities on rapeseed (Brassica napus L.) plantlet growth and morphogenesis in vitro. Sci. Hortic. 2013, 150, 117–124. [Google Scholar] [CrossRef]
- Han, T.; Vaganov, V.; Cao, S.; Li, Q.; Ling, L.; Cheng, X.; Peng, L.; Zhang, C.; Yakovlev, A.N.; Zhong, Y. Improving “color rendering” of LED lighting for the growth of lettuce. Sci. Rep. 2017, 7, 45944. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, K.; Gong, X.; Wang, H.; Gao, Y.; Wang, X.; Zeng, Z.; Hu, Y. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 2020, 19, 108–119. [Google Scholar] [CrossRef]
- Fan, X.; Zang, J.; Xu, Z.; Guo, S.; Jiao, X.; Liu, X.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Verdaguer, D.; Jansen, M.A.K.; Llorens, L.; Morales, L.O.; Neugart, S. UV-A radiation effects on higher plants: Exploring the known unknown. Plant Sci. 2017, 255, 72–81. [Google Scholar] [CrossRef]
- Cooley, N.M.; Higgins, J.T.; Holmes, M.G.; Attridge, T.H. Ecotypic differences in responses of Arabidopsis thaliana L. to elevated polychromatic UV-A and UV-B+ A radiation in the natural environment: A positive correlation between UV-B+ A inhibition and growth rate. J. Photochem. Photobiol. B Biol. 2001, 60, 143–150. [Google Scholar] [CrossRef]
- Qian, M.; Rosenqvist, E.; Flygare, A.-M.; Kalbina, I.; Teng, Y.; Jansen, M.A.K.; Strid, Å. UV-A light induces a robust and dwarfed phenotype in cucumber plants (Cucumis sativus L.) without affecting fruit yield. Sci. Hortic. 2020, 263, 109110. [Google Scholar] [CrossRef]
- Rodríguez-Calcerrada, J.; Pardos, J.A.; Gil, L.; Reich, P.B.; Aranda, I. Light response in seedlings of a temperate (Quercus petraea) and a sub-Mediterranean species (Quercus pyrenaica): Contrasting ecological strategies as potential keys to regeneration performance in mixed marginal populations. Plant Ecol. 2008, 195, 273–285. [Google Scholar] [CrossRef]
- Folta, K.M.; Childers, K.S. Light as a growth regulator: Controlling plant biology with narrow-bandwidth solid-state lighting systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef] [Green Version]
- Monostori, I.; Heilmann, M.; Kocsy, G.; Rakszegi, M.; Ahres, M.; Altenbach, S.B.; Szalai, G.; Pál, M.; Toldi, D.; Simon-Sarkadi, L. LED lighting–modification of growth, metabolism, yield and flour composition in wheat by spectral quality and intensity. Front. Plant Sci. 2018, 9, 605. [Google Scholar] [CrossRef]
- Assmann, S.M. Signal transduction in guard cells. Annu. Rev. Cell Biol. 1993, 9, 345–375. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-H.; Goins, G.D.; Wheeler, R.M.; Sager, J.C. Stomatal conductance of lettuce grown under or exposed to different light qualities. Ann. Bot. 2004, 94, 691–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seabrook, J.E.A. Light effects on the growth and morphogenesis of potato (Solanum tuberosum) in vitro: A review. Am. J. Potato Res. 2005, 82, 353–367. [Google Scholar] [CrossRef]
- Eviatar-Ribak, T.; Shalit-Kaneh, A.; Chappell-Maor, L.; Amsellem, Z.; Eshed, Y.; Lifschitz, E. A cytokinin-activating enzyme promotes tuber formation in tomato. Curr. Biol. 2013, 23, 1057–1064. [Google Scholar] [CrossRef] [Green Version]
- Aksenova, N.P.; Konstantinova, T.N.; Chailakhian, M.K. Morphogenetic effect of blue and red-light under illumination of the overground and underground potato plant organs in vitro. Dokl. Akad. Nauk SSSR 1989, 305, 508–512. [Google Scholar]
- Legris, M.; Ince, Y.Ç.; Fankhauser, C. Molecular mechanisms underlying phytochrome-controlled morphogenesis in plants. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Batutis, E.J.; Ewing, E.E. Far-red reversal of red light effect during long-night induction of potato (Solanum tuberosum L.) tuberization. Plant Physiol. 1982, 69, 672–674. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, L.S. Transduction of blue-light signals. Plant Physiol. 1993, 102, 333. [Google Scholar] [CrossRef] [PubMed]
- Fixen, K.R.; Thomas, S.C.; Tong, C.B.S. Blue light inhibition of tuberization in a day-neutral potato. J. Plant Growth Regul. 2012, 31, 342–350. [Google Scholar] [CrossRef]
- Roumeliotis, E.; Visser, R.G.F.; Bachem, C.W.B. A crosstalk of auxin and GA during tuber development. Plant Signal. Behav. 2012, 7, 1360–1363. [Google Scholar] [CrossRef] [Green Version]
- Lovell, P.H.; Booth, A. Effects of gibberellic acid on growth, tuber formation and carbohydrate distribution in Solanum tuberosum. New Phytol. 1967, 66, 525–537. [Google Scholar] [CrossRef]
- Wang, H.B.; Wang, S.; Wang, X.D.; Shi, X.B.; Wang, B.L.; Zheng, X.C.; Wang, Z.Q.; Liu, F.Z. Effects of light quality on leaf senescence and endogenous hormones content in grapevine under protected cultivation. Chin. J. Appl. Ecol. 2017, 28, 3535. [Google Scholar]
- Su, N.; Wu, Q.; Cui, J. Effects of monochromatic LED light quality on endogenous hormone content in leaves and hypocotyl of Cucumis sativus L. Chin. J. Appl. Environ. Biol. 2014, 20, 45–49. [Google Scholar]
- Kara, A.N.; Kotov, A.A.; Bukhov, N.G. Specific distribution of gibberellins, cytokinins, indole-3-acetic acid, and abscisic acid in radish plants closely correlates with photomorphogenetic responses to blue or red light. J. Plant Physiol. 1997, 151, 51–59. [Google Scholar] [CrossRef]
- Hu, Y. Effects of red and blue LEDs on in vitro growth and microtuberization of potato single-node cuttings. Front. Agr. Sci. Eng. 2018, 5, 197–205. [Google Scholar]
- Asghari-Zakaria, R.; Maleki-Zanjani, B.; Sedghi, E. Effect of in vitro chitosan application on growth and minituber yield of Solanum tuberosum L. Plant Soil Env. 2009, 55, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, C.P.; Pundir, R.K.; Pathak, A.; Joshi, N.; Bagri, D.S. Irradiation Studies of LED Light Spectra on the Growth and Development of Potato (Solanum tuberosum L.). Plant Sci. Today 2020, 7, 406–416. [Google Scholar] [CrossRef]
- Lin, K.-H.; Huang, M.-Y.; Huang, W.-D.; Hsu, M.-H.; Yang, Z.-W.; Yang, C.-M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Turnbull, T.L.; Barlow, A.M.; Adams, M.A. Photosynthetic benefits of ultraviolet-A to Pimelea ligustrina, a woody shrub of sub-alpine Australia. Oecologia 2013, 173, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of different light sources on the growth of non-heading Chinese cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Cao, S.; Shi, L.; Chen, W.; Su, X.; Yang, Z. Blue light irradiation affects anthocyanin content and enzyme activities involved in postharvest strawberry fruit. J. Agric. Food Chem. 2014, 62, 4778–4783. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, J.R.O.; Cordenunsi, B.R.; Lajolo, F.M.; Alcocer, M.J.C. Banana sucrose-phosphate synthase gene expression during fruit ripening. Planta 1997, 203, 283–288. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spectrum Combinations | Ratio (%) | Intensity (µmol m−2 s−1) | Code Name |
|---|---|---|---|
| Natural Light * | L1 | ||
| R:B | 80:20 ** | 300 | L2 |
| R:B:G | 70:20:10 | L3 | |
| R:B:FR | 70:20:10 | L4 | |
| R:B:G:FR | 60:20:10:10 | L5 | |
| R:B:G:FR:UV | 50:20:10:10:10 | L6 | |
| R:B:FR:UV | 60:20:10:10 | L7 | |
| R:B:W:FR: | 50:20:20:10 | L8 | |
| R:B:W:FR:UV | 40:20:20:10:10 | L9 |
| Chemical Name | Vegetative Growth Period (Transplantation to 40th Day) | Tuber Bulking Period (41th Days to Harvesting Day) | ||
|---|---|---|---|---|
| A Tank (50 L) | B Tank (50 L) | A Tank (50 L) | B Tank (50 L) | |
| Ca(NO3)2. 4H2O | 1.5 kg | 7.66 kg | ||
| KNO3 | 3.79 kg | 3.79 kg | 3.54 kg | 3.54 kg |
| (NH4)2HPO4 | 1.6 kg | 1.52 kg | ||
| MgSO4 | 4.3 kg | 3.68 kg | ||
| K2SO4 | 1.3 kg | |||
| Fe-EDTA | 460 g | 460 g | 30.8 g | |
| MnSO4 | 30.8g | |||
| H3BO3 | 57.2 g | 57.2 g | ||
| ZnSO4 | 3.6 g | 3.6 g | ||
| CuSO4 | 1.3 g | 1.3 g | ||
| (NH4)6Mo7O24.4H2O | 0.4 g | 0.4 g | ||
| Treatments | Stem Length (cm) | Stem Diameter (mm) | PFW (g) | PDW (g) | Branch Number | Node Number | Leaf Number | Leaf Length (cm) | Leaf Width (cm) |
|---|---|---|---|---|---|---|---|---|---|
| L1 | 32 ± 3.26 d | 4.5 ± 0.43 cd | 13.1 ± 3.08 d | 1.36 ± 0.37 cde | 4 ± 0.81 b | 21 ± 2.44 bc | 24.61 ± 3.29 b | 15 ± 1.63 cd | 10.3 ± 1.12 d |
| L2 | 37.66 ± 4.18 cd | 6.75 ± 0.95 a | 38.93 ± 4.5 bc | 2.8 ± 0.43 b | 3 ± 0.82 bcd | 19 ± 2.94 bcd | 17.21 ± 1.69 de | 19 ± 1.41 ab | 12.6 ± 1.2 bc |
| L3 | 46.33 ± 3.09 b | 7.02 ± 0.43 a | 51.33 ± 11.08 b | 3.6 ± 1.08 ab | 4.33 ± 0.47 b | 24.33 ± 4.78 b | 23.3 ± 4.18 bc | 22 ± 0.81 a | 15.5 ± 0.4 a |
| L4 | 56.67 ± 7.4 a | 5.33 ± 0.24 bc | 67 ± 14.44 a | 4.4 ± 1.55 a | 7.33 ± 0.94 a | 36.33 ± 2.49 a | 46 ± 3.25 a | 19 ± 0.8 ab | 13.16 ± 0.62 bc |
| L5 | 32 ± 4.32 d | 5.34 ± 0.38 bc | 16.46 ± 4.92 d | 0.96 ± 0.33 de | 2.33 ± 0.47 cd | 14.32 ± 1.69 e | 14.66 ± 3.08 e | 13.3 ± 2.86 d | 9.66 ± 1.19 de |
| L6 | 40.37 ± 2.49 bc | 4 ± 0.21 d | 12.86 ± 1.92 d | 0.56 ± 0.04 e | 1.66 ± 0.47 d | 14.23 ± 1.66 e | 14.61 ± 2.62 e | 12.66 ± 0.47 d | 8 ± 0.81 e |
| L7 | 46.33 ± 3.68 b | 5.06 ± 0.73 c | 37.33 ± 4.98 c | 2.26 ± 0.44 bcd | 2.33 ± 0.41 cd | 16 ± 3.26 cde | 19.32 ± 2.49 cde | 17 ± 0.82 bc | 11.6 ± 1.24 cd |
| L8 | 44 ± 2.94 bc | 6.61 ± 0.35 a | 46.66 ± 3.39 bc | 3.1 ± 0.37 ab | 3.66 ± 0.33 bc | 21.23 ± 2.49 bc | 19 ± 0.81 cde | 19 ± 1.4 ab | 13.83 ± 0.62 abc |
| L9 | 42 ± 2.44 bc | 6.21 ± 0.36 ab | 45.33 ± 6.54 bc | 2.7 ± 0.32 bc | 6 ± 0.83 a | 23.31 ± 3.09 b | 19.66 ± 1.24 cde | 19.6 ± 1.22 ab | 14.33 ± 1.21 ab |
| LSD(0.05) | 8.57 | 0.98 | 13.23 | 1.35 | 1.49 | 5.96 | 5.18 | 3.15 | 2.24 |
| Treatments | Root Length (cm) | RFW (g) | RDW (g) | Stolon Length (cm) | SFW (g) | SDW (g) |
|---|---|---|---|---|---|---|
| L1 | 26.66 ± 2.49 cd | 3.04 ± 1.16 e | 0.2 ± 0.04 cd | 34.66 ± 4.49 bc | 2.1 ± 0.35 d | 0.1 ± 0.01 de |
| L2 | 28.33 ± 2.35 bcd | 5.16 ± 0.89 bcde | 0.37 ± 0.05 bcd | 38 ± 4.32 ab | 2.7 ± 0.5 bcd | 0.17 ± 0.03 bc |
| L3 | 34 ± 2.16 ab | 6.8 ± 1.29 abcd | 0.54 ± 0.14 ab | 34.32 ± 2.62 bc | 2.93 ± 0.61 bcd | 0.21 ± 0.05 ab |
| L4 | 37.21 ± 6.54 a | 10.1 ± 3.47 a | 0.62 ± 0.29 a | 43.61 ± 7.58 a | 4.7 ± 0.86 a | 0.24 ± 0.04 a |
| L5 | 23.61 ± 3.39 d | 3.5 ± 1.44 cde | 0.13 ± 0.06 d | 24 ± 5.09 d | 2.2 ± 0.68 cd | 0.05 ± 0.01 ef |
| L6 | 30 ± 1.63 bcd | 3.3 ± 1.52 de | 0.12 ± 0.08 cd | 27 ± 3.55 cd | 1.03 ± 0.49 e | 0.02 ± 0.01 f |
| L7 | 29 ± 2.16 bcd | 7.1 ± 1.75 abc | 0.29 ± 0.11 cd | 29.3 ± 4.18 bcd | 3.2 ± 0.48 bc | 0.05 ± 0.02 ef |
| L8 | 32 ± 1.63 abc | 7 ± 1.43 abc | 0.54 ± 0.05 ab | 35.66 ± 3.29 abc | 2.76 ± 0.4 bcd | 0.13 ± 0.02 cd |
| L9 | 35 ± 4.08 ab | 8.33 ± 1.58 ab | 0.42 ± 0.06 abc | 33.12 ± 1.24 bc | 3.33 ± 0.75 b | 0.07 ± 0.01 def |
| LSD(0.05) | 7.02 | 3.6 | 0.24 | 8.97 | 1.03 | 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.H.; Islam, M.J.; Azad, M.O.K.; Rana, M.S.; Ryu, B.R.; Lim, Y.-S. LED Light Pre-Treatment Improves Pre-Basic Seed Potato (Solanum tuberosum L. cv. Golden King) Production in the Aeroponic System. Agronomy 2021, 11, 1627. https://doi.org/10.3390/agronomy11081627
Rahman MH, Islam MJ, Azad MOK, Rana MS, Ryu BR, Lim Y-S. LED Light Pre-Treatment Improves Pre-Basic Seed Potato (Solanum tuberosum L. cv. Golden King) Production in the Aeroponic System. Agronomy. 2021; 11(8):1627. https://doi.org/10.3390/agronomy11081627
Chicago/Turabian StyleRahman, Md. Hafizur, Md. Jahirul Islam, Md. Obyedul Kalam Azad, Md. Soyel Rana, Byeong Ryeol Ryu, and Young-Seok Lim. 2021. "LED Light Pre-Treatment Improves Pre-Basic Seed Potato (Solanum tuberosum L. cv. Golden King) Production in the Aeroponic System" Agronomy 11, no. 8: 1627. https://doi.org/10.3390/agronomy11081627