
Tillage Intensity Influences Insect-Pest and Predator Dynamics of Wheat Crop Grown under Different Conservation Agriculture Practices in Rice-Wheat Cropping System of Indo-Gangetic Plain

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Tillage Operations
2.2. Weather Conditions
2.3. Insect Sampling
2.4. Normalized Difference Vegetative Index (NDVI)
2.5. Canopy Temperature
2.6. Statistical Analysis
3. Results
3.1. Foliar Aphid, R. maidis
3.2. Root Aphid, R. rufiabdominalis
3.3. Termites, O. obesus and M. obesi
3.4. Pink Stem Borer, S. inferens
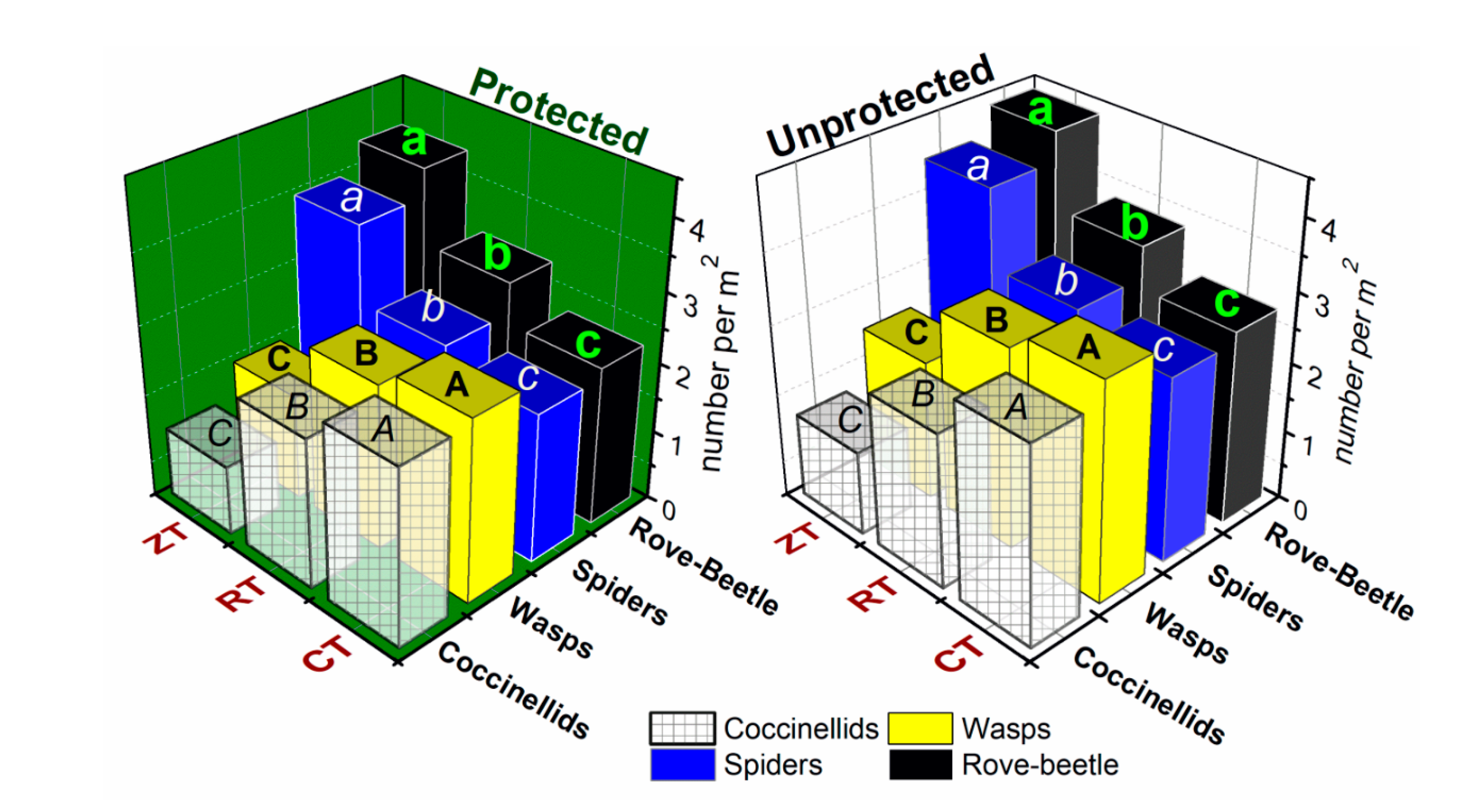
3.5. Natural Enemies
3.6. Relationship between Normalized Difference Vegetative Index (NDVI) and Insect-Pest Abundance and Damage
3.7. Relationship between Canopy Temperature and Insect-Pest Abundance and Damage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R. Soils and world food security. Soil Tillage Res. 2009, 102, 1–4. [Google Scholar] [CrossRef]
- Bhardwaj, A.K.; Jasrotia, P.; Hamilton, S.K.; Robertson, G.P. Ecological management of intensively cropped agro-ecosystems improves soil quality with sustained productivity. Agric. Ecosyst. Environ. 2011, 140, 419–429. [Google Scholar] [CrossRef]
- Henneron, L.; Bernard, L.; Hedde, M.; Pelosi, C.; Villenave, C.; Chenu, C.; Bertrand, M.; Girardin, C.; Blanchart, E. Fourteen years of evidence for positive effects of conservation agriculture and organic farming on soil life. Agron. Sustain. Dev. 2015, 35, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, A.K.; Rajwar, D.; Mandal, U.K.; Ahamad, S.; Kaphaliya, B.; Minhas, P.S.; Prabhakar, M.; Banyal, R.; Singh, R.; Chaudhari, S.K.; et al. Impact of carbon inputs on soil carbon fractionation, sequestration and biological responses under major nutrient management practices for rice-wheat cropping systems. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Erenstein, O.; Sayre, K.D.; Wall, P.; Hellin, J.; Dixon, J. Conservation agriculture in maize- and wheat-based systems in the (sub) tropics: Lessons from adaptation initiatives in South Asia, Mexico, and Southern Africa. J. Sustain. Agric. 2012, 36, 180–206. [Google Scholar] [CrossRef]
- Busari, M.A.; Kukal, S.S.; Kaur, A.; Bhatt, R.; Dulazi, A.A. Conservation tillage impacts on soil, crop and the environment. Int. Soil Water Conserv. Res. 2015, 3, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Six, J.; Ogle, S.M.; Jay Breidt, F.; Conant, R.T.; Mosier, A.R.; Paustian, K. The potential to mitigate global warming with no-tillage management is only realized when practised in the long term. Glob.Chang. Biol. 2004, 10, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Ruan, L.; Philip Robertson, G. Initial nitrous oxide, carbon dioxide, and methane costs of converting conservation reserve program grassland to row crops under no-till vs. Conventional tillage. Glob. Chang. Biol. 2013, 19, 2478–2489. [Google Scholar] [CrossRef] [Green Version]
- Dhaliwal, G.S.; Jindal, V.; Dhawan, A.K. Insect pest problems and crop losses: Changing trends. Indian J. Ecol. 2010, 37, 1–7. [Google Scholar]
- Singh, B.; Kular, J.S.; Ram, H.; Mahal, M.S. Relative abundance and damage of some insect pests of wheat under different tillage practices in rice–wheat cropping in India. Crop Prot. 2014, 61, 16–22. [Google Scholar] [CrossRef]
- Katare, S.; Jasrotia, P.; Patil, S.D.; Reza, M.W.; Saharan, M.S. Influence of sowing time and weather factors on seasonal dynamics of aphids in three wheat growing zones of India. J. Agrometeorol. 2018, 20, 134–138. [Google Scholar]
- Joshi, A.K.; Chand, R.; Arun, B. Wheat improvement in eastern and warmer regions of ndia: Conventional and non-conventional approaches. In A Compendium of Training Program (26–30 December, 2003); NATP, Indian Council of Agricultural Research—Banaras Hindu University: Varanasi, India, 2004. [Google Scholar]
- Ram, H.; Singh, B.; Sharma, I.; Bimbraw, A.S.; Mavi, G.S. Potentials of Resource Conservation Technology and Incidence of Pink Stem Borer (Sesamia inferens) in Various Varieties of Wheat (Triticum aestivum L.). In Proceedings of the 3rd International Group Meeting on “Wheat Productivity Enhancement under Changing Climate”; University of Agricultural Sciences: Dharwad, India, 2011; p. 149. [Google Scholar]
- Singh, B. Incidence of the pink noctuid stem borer, Sesamia inferens (Walker), on wheat under two tillage conditions and three sowing dates in north-western plains of India. J. Entomol. 2012, 9, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Jaipal, S.; Malik, R.K.; Yadav, A.; Gupta, R. IPM issues in zero-tillage system in rice-wheat cropping sequence. Tech. Bull. 2005, 8, 32. [Google Scholar]
- Jasrotia, P.; Katare, S. Compatibility of insecticides with propiconazole against foliar aphid Rhopalosiphum maidis (Fitch) and yellow rust in wheat. Indian J. Entomol. 2018, 80, 1304–1309. [Google Scholar] [CrossRef]
- Macfadyen, S.; Kriticos, D.J. Modelling the geographical range of a species with variable life-history. PLoS ONE 2012, 7, e40313. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, M.R.; Daniel, T.C.; Schweizer, E.E.; Allmaras, R.R. Conservation tillage. Science 1985, 230, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Kladivko, E.J. Tillage systems and soil biology. Soil Tillage Res. 2001, 61, 61–76. [Google Scholar] [CrossRef]
- Burton, R.L.; Krenzer Jr, E.G. Reduction of greenbug (Homoptera: Aphididae) populations by surface residues in wheat tillage studies. J. Econ. Entomol. 1985, 78, 390–394. [Google Scholar] [CrossRef]
- Hesler, L.S.; Berg, R.K. Tillage impact cereal-aphid (Homoptera: Aphididae) infestation in spring small grains. J. Econ. Entomol. 2003, 96, 1792–1797. [Google Scholar] [CrossRef]
- Clement, S.; Elberson, L.; Youssef, N.; Young, F.; Evens, M. Cereal aphid and natural enemies populations in cereal production systems in Eastern Washington. J. Kans. Entomol. Soc. 2004, 77, 165–173. [Google Scholar] [CrossRef]
- Dong, Z.; Hou, R.; Ouyang, Z.; Zhang, R. Tritrophic interaction influenced by warming and tillage: A field study on winter wheat, aphids and parasitoids. Agric. Ecosyst. Environ. 2013, 181, 144–148. [Google Scholar] [CrossRef]
- Kooyman, C.H.R.; Onck, R.F.M. The Interactions between Termite Activity, Agricultural Practices and Soil Characteristics in Kisii District, Kenya (No. 87-3); Agricultural University: Wageningen, The Netherlands, 1987. [Google Scholar]
- Rathore, L.; Sharma, P.K. Field efficacy of insecticides and biopesticides against aphid complex in wheat. J. Entomol. Res. 2016, 40, 77–80. [Google Scholar] [CrossRef]
- Gadhiya, V.C.; Borad, P.K. Effect of insecticidal seed treatment on reduction of termite damage and increase in wheat yield. Pestic. Res. J. 2013, 25, 87–89. [Google Scholar]
- Rivers, A.; Barbercheck, M.; Govaerts, B.; Verhulst, N. Conservation agriculture affects arthropod community composition in a rainfed maize–wheat system in central Mexico. Appl. Soil Ecol. 2016, 100, 81–90. [Google Scholar] [CrossRef]
- Shearin, A.F.; Reberg-Horton, S.C.; Gallandt, E.R. Direct effects of tillage on the activity density of ground beetle (Coleoptera: Carabidae) weed seed predators. Environ. Entomol. 2014, 36, 1140–1146. [Google Scholar] [CrossRef]
- Rivers, A.; Mullen, C.; Wallace, J.; Barbercheck, M. Cover crop-based reduced tillage system influences Carabidae (Coleoptera) activity, diversity and trophic group during transition to organic production. Renew. Agric. Food Syst. 2017, 32, 538. [Google Scholar] [CrossRef]
- Thorbek, P.; Bilde, T. Reduced numbers of generalist arthropod predators after crop management. J. Appl. Ecol. 2004, 41, 526–538. [Google Scholar] [CrossRef]
- Tamburini, G.; De Simone, S.; Sigura, M.; Boscutti, F.; Marini, L. Conservation tillage mitigates the negative effect of landscape simplification on biological control. J. Appl. Ecol. 2016, 53, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Rice, M.E.; Wilde, G.E. Aphid predators associated with conventional-and conservation-tillage winter wheat. J. Kans. Entomol. Soc. 1991, 64, 245–250. [Google Scholar]
- Wilson-Rummenie, A.C.; Radford, B.J.; Robertson, L.N.; Simpson, G.B.; Bell, K.L. Reduced tillage increases population density of soil macrofauna in a semiarid environment in central Queensland. Environ. Entomol. 1999, 28, 163–172. [Google Scholar] [CrossRef]
- Andersen, A. Long-term experiments with reduced tillage in spring cereals. II. Effects on pests and beneficial insects. Crop Prot. 2003, 22, 147–152. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Okland, B. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Saudreau, M.; Pincebourde, S.; Dassot, M.; Adam, B.; Loxdale, H.D.; Biron, D.G. On the canopy structure manipulation to buffer climate change effects on insect herbivore development. Trees 2013, 27, 239–248. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks after Sowing (WAS) | Dates | Standard Meteorological Weeks | 2016–2017 | 2017–2018 | ||||
|---|---|---|---|---|---|---|---|---|
| Average Temperature (°C) | Average Relative Humidity (%) | Total Rainfall (mm/day) | Average Temperature (°C) | Average Relative Humidity (%) | Total Rainfall mm/day | |||
| 1 | 19 Nov–25 Nov | 47 | 19.1 | 59.5 | 0.0 | 16.0 | 57.8 | 0.0 |
| 2 | 26 Nov–02 Dec | 48 | 19.6 | 62.9 | 0.0 | 15.9 | 61.4 | 0.0 |
| 3 | 03 Dec–09 Dec | 49 | 15.4 | 80.9 | 0.0 | 15.9 | 60.9 | 0.0 |
| 4 | 10 Dec–16 Dec | 50 | 15.6 | 81.4 | 0.0 | 14.2 | 82.4 | 5.2 |
| 5 | 17 Dec–23 Dec | 51 | 14.4 | 72.4 | 0.0 | 14.4 | 74.6 | 0.0 |
| 6 | 24 Dec–31 Dec | 52 | 14.1 | 77.1 | 0.0 | 13.5 | 80.7 | 0.0 |
| 7 | 01 Jan–07 Jan | 1 | 15.3 | 80.9 | 8.0 | 9.6 | 89.8 | 0.0 |
| 8 | 08 Jan–14 Jan | 2 | 9.8 | 80.4 | 19.0 | 12.1 | 70.5 | 0.0 |
| 9 | 15 Jan–21 Jan | 3 | 11.2 | 81.0 | 3.6 | 13.3 | 74.4 | 0.0 |
| 10 | 22 Jan–28 Jan 30 | 4 | 15.1 | 84.1 | 55.2 | 12.2 | 86.1 | 34.2 |
| 11 | 29 Jan–04 Feb | 5 | 14.2 | 80.4 | 0.0 | 14.9 | 72.0 | 0.0 |
| 12 | 05 Feb–11 Feb | 6 | 14.0 | 75.2 | 0.0 | 13.2 | 67.0 | 0.0 |
| 13 | 12 Feb–18 Feb | 7 | 16.1 | 72.9 | 0.0 | 15.0 | 75.9 | 29.0 |
| 14 | 19 Feb–25 Feb | 8 | 17.5 | 67.4 | 0.0 | 17.6 | 75.4 | 0.0 |
| 15 | 26 Feb–04 Mar | 9 | 17.8 | 62.3 | 0.3 | 19.9 | 72.4 | 0.0 |
| 16 | 05 Mar–11 Mar | 10 | 17.5 | 63.9 | 7.5 | 19.3 | 66.9 | 0.0 |
| 17 | 12 Mar–18 Mar | 11 | 15.5 | 64.6 | 0.0 | 21.2 | 62.4 | 0.0 |
| 18 | 19 Mar–25 Mar | 12 | 22.0 | 60.9 | 0.0 | 21.6 | 62.9 | 0.0 |
| Source | Foliar Aphid | Root Aphid | Termites | Pink Stem Borer | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | df | F | p | |
| Tillage | 2 | 3.85 | 0.04 | 2 | 3.28 | 0.04 | 2 | 9.09 | 0.04 | 2 | 20.11 | 0.0002 |
| Insecticide protection | 1 | 5.13 | 0.03 | 1 | 18.77 | 0.001 | 1 | 7.24 | 0.014 | 1 | 40.36 | 0.0001 |
| Time of observation | 4 | 13.36 | 0.000 | 2 | 1.25 | 0.329 | 3 | 1.32 | 0.032 | 4 | 1.65 | 0.211 |
| Tillage × insecticide protection | 2 | 0.09 | 0.902 | 2 | 1.21 | 0.03 | 2 | 1.27 | 0.302 | 2 | 0.28 | 0.04 |
| Tillage × time of observation | 8 | 0.33 | 0.940 | 4 | 0.05 | 0.993 | 6 | 0.09 | 0.510 | 8 | 0.16 | 0.999 |
| Tillage × insecticide protection × Time of observation | 8 | 0.12 | 0.027 | 4 | 0.13 | 0.968 | 6 | 0.13 | 0.989 | 8 | 0.10 | 0.825 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasrotia, P.; Bhardwaj, A.K.; Katare, S.; Yadav, J.; Kashyap, P.L.; Kumar, S.; Singh, G.P. Tillage Intensity Influences Insect-Pest and Predator Dynamics of Wheat Crop Grown under Different Conservation Agriculture Practices in Rice-Wheat Cropping System of Indo-Gangetic Plain. Agronomy 2021, 11, 1087. https://doi.org/10.3390/agronomy11061087
Jasrotia P, Bhardwaj AK, Katare S, Yadav J, Kashyap PL, Kumar S, Singh GP. Tillage Intensity Influences Insect-Pest and Predator Dynamics of Wheat Crop Grown under Different Conservation Agriculture Practices in Rice-Wheat Cropping System of Indo-Gangetic Plain. Agronomy. 2021; 11(6):1087. https://doi.org/10.3390/agronomy11061087
Chicago/Turabian StyleJasrotia, Poonam, Ajay Kumar Bhardwaj, Subhash Katare, Jayant Yadav, Prem Lal Kashyap, Sudheer Kumar, and Gyanendra Pratap Singh. 2021. "Tillage Intensity Influences Insect-Pest and Predator Dynamics of Wheat Crop Grown under Different Conservation Agriculture Practices in Rice-Wheat Cropping System of Indo-Gangetic Plain" Agronomy 11, no. 6: 1087. https://doi.org/10.3390/agronomy11061087