
Dynamic of Morphological and Physiological Parameters and Variation of Soil Characteristics during Miscanthus × giganteus Cultivation in the Diesel-Contaminated Land

 , , , and
, , , and
Abstract
:1. Introduction
- (a)
- Impact of amending soil by biochars to the soil parameters, specifically: pH, NO3, NH4, P2O5, and K during M. × giganteus vegetation versus changes of parameters in soil without crops;
- (b)
- Impact of varied diesel concentrations in the soil and its biochar amending to M. × giganteus development verifying by plant’s morphological and physiological parameters;
- (c)
- Ensuring M. × giganteus adaptive potential while the crop was cultivated in the diesel-contaminated soils, including with biochars via determination of NPQt and SPAD values.
2. Materials and Methods
2.1. Design of the Pot Experiment
- (a)
- Experiment without amendments (marked as Ctr);
- (b)
- Experiment with biochar 1 (marked as B1);
- (c)
- Experiment with biochar 2 (marked as B2).
2.2. Soil Artificial Contamination and Amending
2.3. Analysis of Soil Parameters
2.4. Measuring of Plant’s Morphological and Physiological Parameters
- (a)
- Changes of plants’ bioparameters: height, the number of stems; the number of leaves per one stem; leaves width and length, from which the leaves surface area (LSA) was calculated using the following equation:
- (b)
- Changes in plant state during vegetation by measuring chlorophyll fluorescence [49,50]. The measurement was made on intact, fully expanded leaves using the MultispeQ v1.0 device [51] linked to the PhotosynQ platform (http://www.photosynq.com/technology, accessed on 17 February 2021).
2.5. Statistical Analysis
3. Results and Discussion
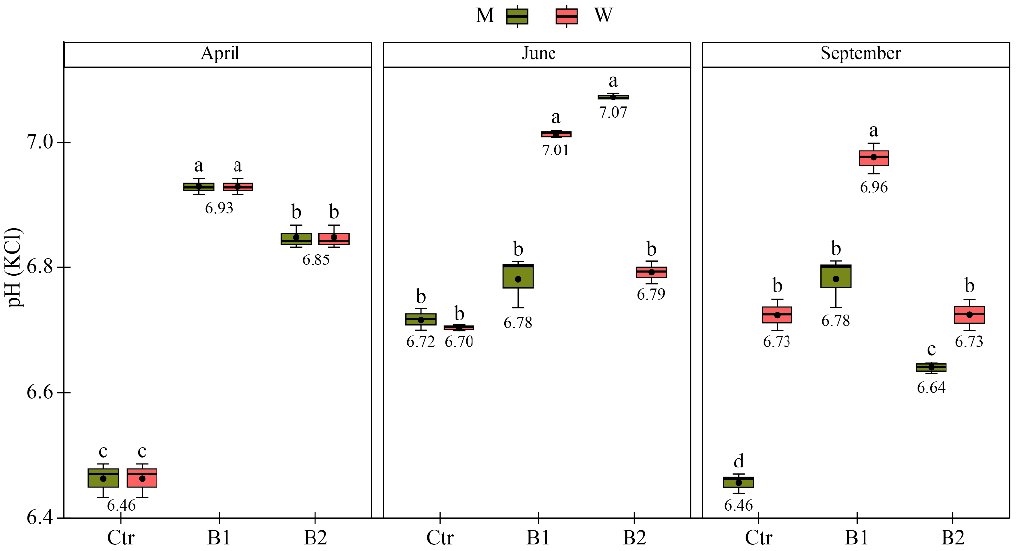
3.1. Impact of M. × giganteus Growing to the Parameters of Uncontaminated Soil
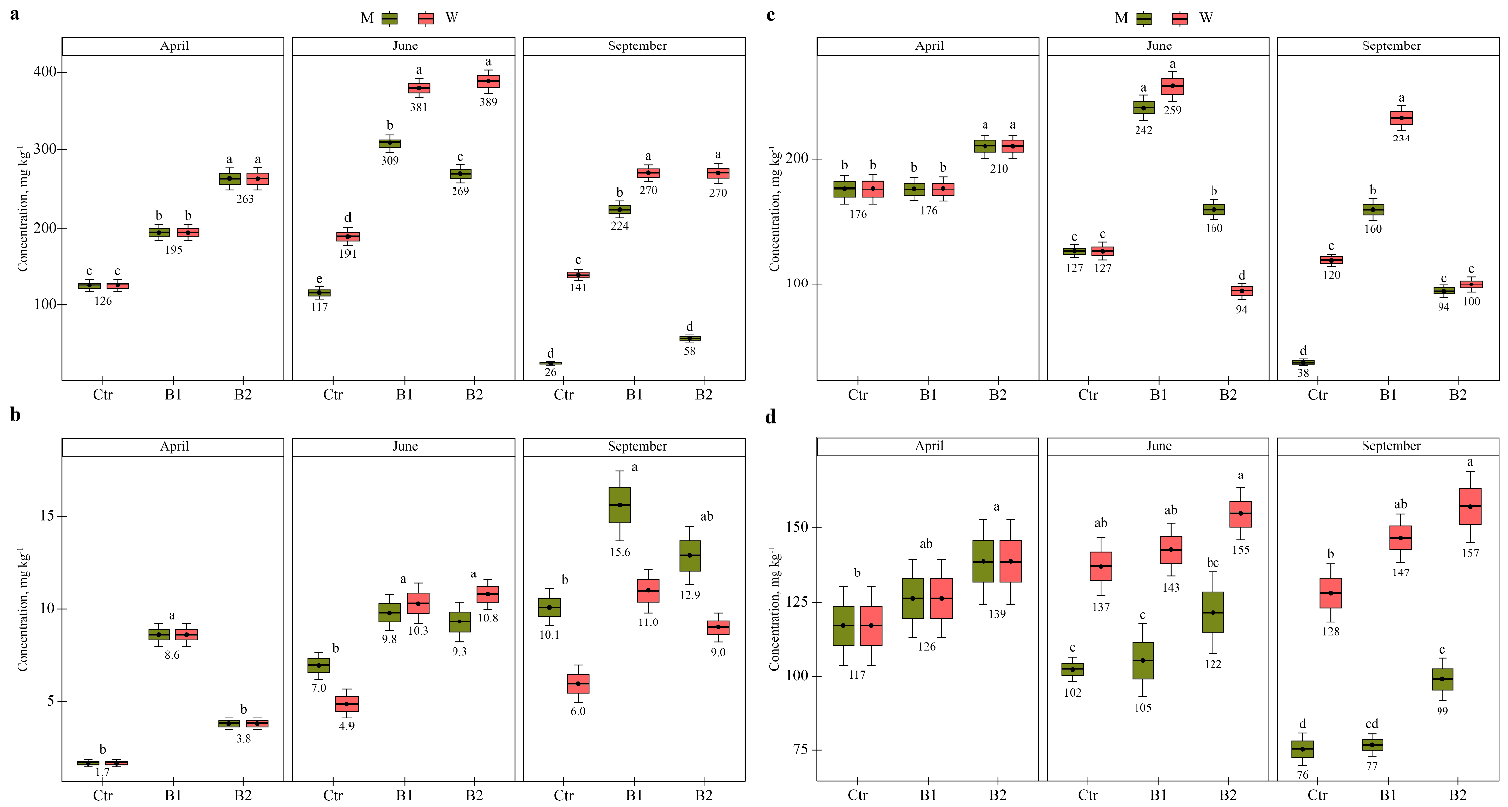
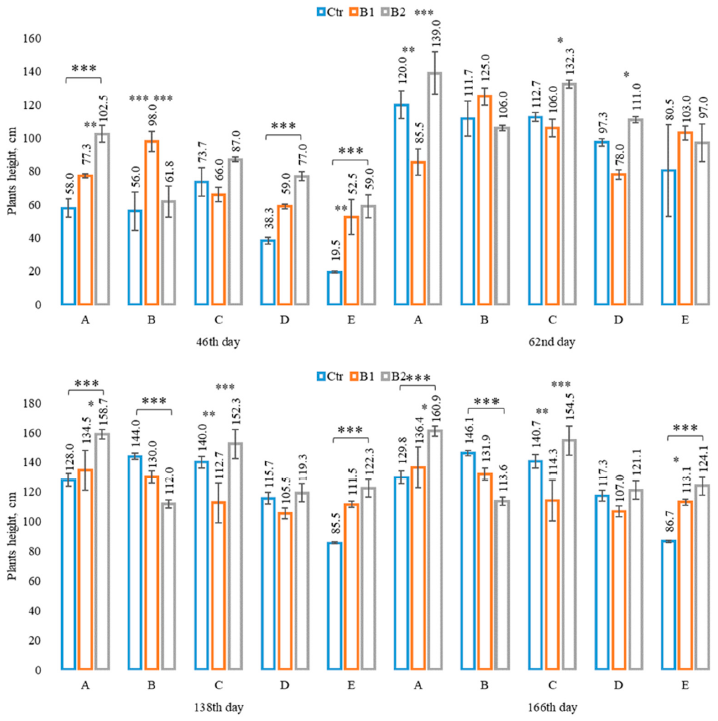
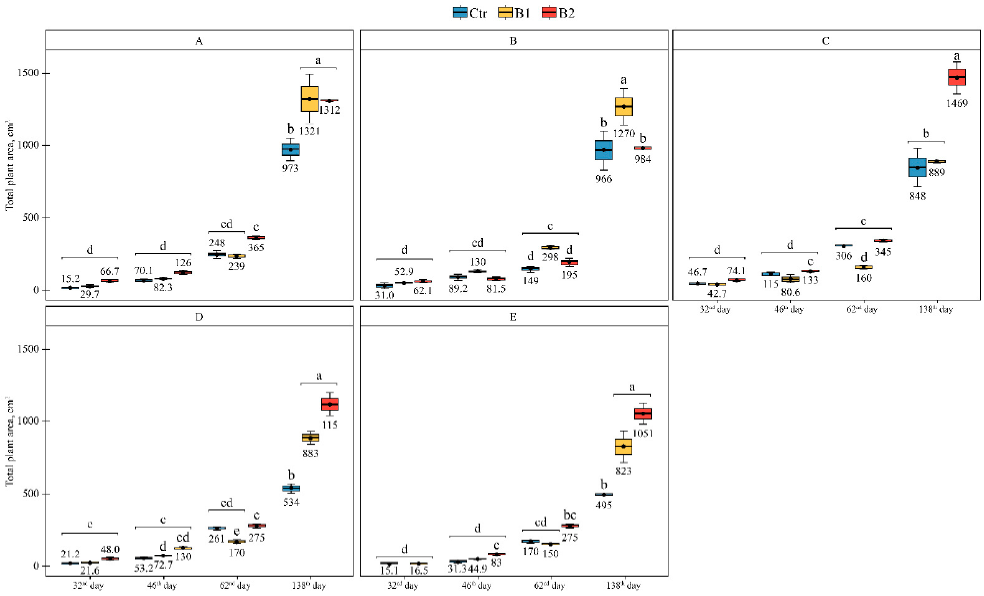
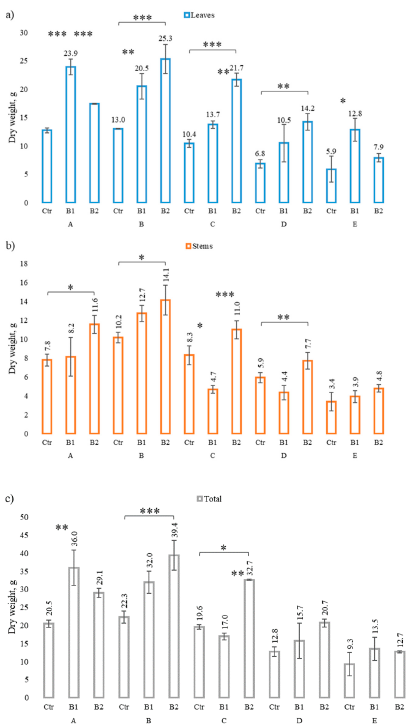
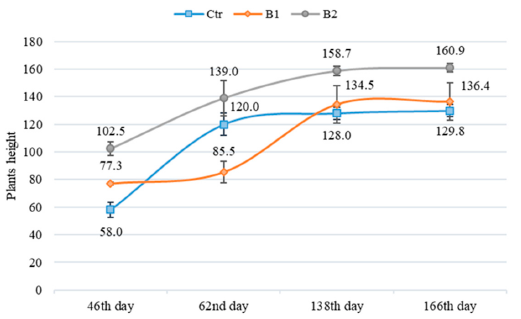
3.2. Impact of Soil Contamination by Diesel to M. × giganteus Bioparameters
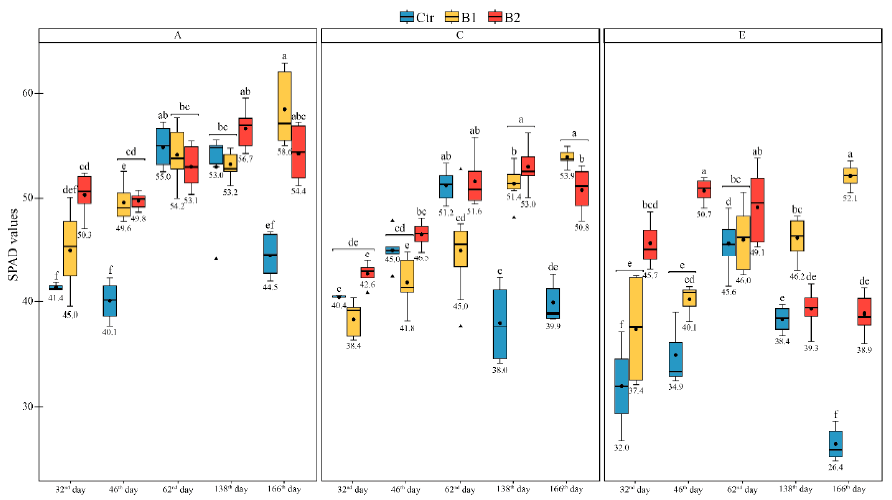
3.3. Impact of Soil Amendment by Biochar and Contamination by Diesel to the Plant State
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Eugenio, N.; McLaughlin, M.; Pennock, D. Soil Pollution: A Hidden Reality; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Okere, U.; Semple, K. Biodegradation of PAHs in ‘Pristine’Soils from Different Climatic Regions. J. Bioremed. Biodegrad. 2012, S1, 6. [Google Scholar] [CrossRef] [Green Version]
- Korchagina, L. Features of plants growing in upland bogs in the conditions of oil pollution in the middle Ob’s region. News Nizhnevartovsk State Univ. Math. Nat. Sci. 2015, 1, 8. [Google Scholar]
- Roy, S.; Labelle, S.; Mehta, P.; Mihoc, A.; Fortin, N.; Masson, C.; Leblanc, R.; Châteauneuf, G.; Sura, C.; Gallipeau, C.; et al. Phytoremediation of Heavy Metal and PAH-Contaminated Brownfield Sites. Plant Soil 2005, 272, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Podan, I.; Dzura, N. Impact of Oil Contamination and Humates to the Development of Miscanthus Plants. Ecol. Sci. 2019, 182–186. [Google Scholar] [CrossRef]
- Técher, D.; Martinez-Chois, C.; Laval-Gilly, P.; Henry, S.; Bennasroune, A.; D’Innocenzo, M.; Falla, J. Assessment of Miscanthus × giganteus for Rhizoremediation of Long Term PAH Contaminated Soils. Appl. Soil Ecol. 2012, 62, 42–49. [Google Scholar] [CrossRef]
- Gerhardt, K.E.; Huang, X.-D.; Glick, B.R.; Greenberg, B.M. Phytoremediation and Rhizoremediation of Organic Soil Contaminants: Potential and Challenges. Plant Sci. 2009, 176, 20–30. [Google Scholar] [CrossRef]
- Susarla, S.; Medina, V.F.; McCutcheon, S.C. Phytoremediation: An Ecological Solution to Organic Chemical Contamination. Ecol. Eng. 2002, 18, 647–658. [Google Scholar] [CrossRef]
- Zhang, C.; Feng, Y.; Liu, Y.; Chang, H.; Li, Z.; Xue, J. Uptake and Translocation of Organic Pollutants in Plants: A Review. J. Integr. Agric. 2017, 16, 1659–1668. [Google Scholar] [CrossRef] [Green Version]
- Brosse, N.; Dufour, A.; Meng, X.; Sun, Q.; Ragauskas, A. Miscanthus: A Fast-Growing Crop for Biofuels and Chemicals Production. Biofuels Bioprod. Biorefining 2012, 6, 580–598. [Google Scholar] [CrossRef]
- McCalmont, J.P.; Hastings, A.F.S.J.; McNamara, N.P.; Richter, G.M.; Robson, P.; Donnison, I.S.; Clifton-Brown, J.C. Environmental Costs and Benefits of Growing Miscanthus for Bioenergy in the UK. GCB Bioenergy 2015, 9, 489–507. [Google Scholar] [CrossRef]
- Nsanganwimana, F.; Pourrut, B.; Mench, M.; Douay, F. Suitability of Miscanthus Species for Managing Inorganic and Organic Contaminated Land and Restoring Ecosystem Services. A Review. J. Environ. Manag. 2014, 143, 123–134. [Google Scholar] [CrossRef]
- Wanat, N.; Austruy, A.; Joussein, E.; Soubrand, M.; Hitmi, A.; Gauthier-Moussard, C.; Lenain, J.-F.; Vernay, P.; Munch, J.C.; Pichon, M. Potentials of Miscanthus × giganteus Grown on Highly Contaminated Technosols. J. Geochem. Explor. 2013, 126–127, 78–84. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.; Stefanovska, T.; Lewis, E.E.; Erickson, L.E.; Davis, L.C. Miscanthus as a Productive Biofuel Crop for Phytoremediation. Crit. Rev. Plant Sci. 2014, 33, 1–19. [Google Scholar] [CrossRef]
- Andrejić, G.; Šinžar-Sekulić, J.; Prica, M.; Dželetović, Ž.; Rakić, T. Phytoremediation Potential and Physiological Response of Miscanthus × giganteus Cultivated on Fertilized and Non-Fertilized Flotation Tailings. Environ. Sci. Pollut. Res. 2019, 26, 34658–34669. [Google Scholar] [CrossRef]
- Cosentino, S.L.; Scordia, D.; Testa, G.; Monti, A.; Alexopoulou, E.; Christou, M. 1—The Importance of Perennial Grasses as a Feedstock for Bioenergy and Bioproducts. In Perennial Grasses for Bioenergy and Bioproducts; Alexopoulou, E., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1–33. ISBN 978-0-12-812900-5. [Google Scholar]
- Lewandowski, I.; Clifton-Brown, J.; Kiesel, A.; Hastings, A.; Iqbal, Y. 2—Miscanthus. In Perennial Grasses for Bioenergy and Bioproducts; Alexopoulou, E., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 35–59. ISBN 978-0-12-812900-5. [Google Scholar]
- Roik, M.; Sinchenko, V.; Purkin, V.; Kvak, V.; Humentik, M. (Eds.) Miscanthus in Ukraine; FOP Yamchinskiy Press: Kyiv, Ukraine, 2019; ISBN 978-617-7804-11-5. [Google Scholar]
- Pidlisnyuk, V.V.; Shapoval, P.; Zgorelec, Z.; Stefanovska, T.; Zhukov, O. Multiyear Phytoremediation and Dynamic of Foliar Metal(Loid)s Concentration during Application of Miscanthus × giganteus Greef et Deu to Polluted Soil from Bakar, Croatia. Environ. Sci. Pollut. Res. 2020. [Google Scholar] [CrossRef]
- Nebeská, D.; Trögl, J.; Pidlisnyuk, V.; Popelka, J.; Veronesi Dáňová, P.; Usťak, S.; Honzík, R. Effect of Growing Miscanthus × giganteus on Soil Microbial Communities in Post-Military Soil. Sustainability 2018, 10, 4021. [Google Scholar] [CrossRef] [Green Version]
- Técher, D.; Laval-Gilly, P.; Henry, S.; Bennasroune, A.; Formanek, P.; Martinez-Chois, C.; D’Innocenzo, M.; Muanda, F.; Dicko, A.; Rejšek, K.; et al. Contribution of Miscanthus × giganteus Root Exudates to the Biostimulation of PAH Degradation: An in Vitro Study. Sci. Total Environ. 2011, 409, 4489–4495. [Google Scholar] [CrossRef]
- Nebeská, D.; Auer Malinská, H.; Erol, A.; Pidlisnyuk, V.; Kuráň, P.; Medžová, A.; Smaha, M.; Trögl, J. Stress Response of Miscanthus Plants and Soil Microbial Communities: A Case Study in Metals and Hydrocarbons Contaminated Soils. Appl. Sci. 2021, 11, 1866. [Google Scholar] [CrossRef]
- Ciarkowska, K.; Gambus, F.; Antonkiewicz, J.; Koliopoulos, T. Polycyclic Aromatic Hydrocarbon and Heavy Metal Contents in the Urban Soils in Southern Poland. Chemosphere 2019, 229, 214–226. [Google Scholar] [CrossRef]
- Ptaszek, N.; Pacwa-Płociniczak, M.; Noszczyńska, M.; Płociniczak, T. Comparative Study on Multiway Enhanced Bio- and Phytoremediation of Aged Petroleum-Contaminated Soil. Agronomy 2020, 10, 947. [Google Scholar] [CrossRef]
- Antonkiewicz, J.; Kołodziej, B.; Bielińska, E.J.; Popławska, A. The Possibility of Using Sewage Sludge for Energy Crop Cultivation Exemplified by Reed Canary Grass and Giant Miscanthus. Soil Sci. Annu. 2019, 70, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Kharytonov, M.; Pidlisnyuk, V.; Stefanovska, T.; Babenko, M.; Martynova, N.; Rula, I. The Estimation of Miscanthus × giganteus’ Adaptive Potential for Cultivation on the Mining and Post-Mining Lands in Ukraine. Environ. Sci. Pollut. Res. 2019, 26, 2974–2986. [Google Scholar] [CrossRef] [PubMed]
- Alasmary, Z.; Todd, T.; Hettiarachchi, G.M.; Stefanovska, T.; Pidlisnyuk, V.; Roozeboom, K.; Erickson, L.; Davis, L.; Zhukov, O. Effect of Soil Treatments and Amendments on the Nematode Community under Miscanthus Growing in a Lead Contaminated Military Site. Agronomy 2020, 10, 1727. [Google Scholar] [CrossRef]
- Mamirova, A.; Pidlisnyuk, V.; Amirbekov, A.; Ševců, A.; Nurzhanova, A. Phytoremediation Potential of Miscanthus sinensis And. in Organochlorine Pesticides Contaminated Soil Amended by Tween 20 and Activated Carbon. Environ. Sci. Pollut. Res. 2021, 28, 16092–16106. [Google Scholar] [CrossRef]
- Pidlisnyuk, V.V.; Erickson, L.; Stefanovska, T.; Popelka, J.; Hettiarachchi, G.; Davis, L.; Trögl, J. Potential Phytomanagement of Military Polluted Sites and Biomass Production Using Biofuel Crop Miscanthus × giganteus. Environ. Pollut. 2019, 249, 330–337. [Google Scholar] [CrossRef]
- Crane-Droesch, A.; Abiven, S.; Jeffery, S.; Torn, M.S. Heterogeneous Global Crop Yield Response to Biochar: A Meta-Regression Analysis. Environ. Res. Lett. 2013, 8, 044049. [Google Scholar] [CrossRef]
- Kloss, S.; Zehetner, F.; Wimmer, B.; Buecker, J.; Rempt, F.; Soja, G. Biochar Application to Temperate Soils: Effects on Soil Fertility and Crop Growth under Greenhouse Conditions. J. Plant Nutr. Soil Sci. 2014, 177, 3–15. [Google Scholar] [CrossRef]
- Saletnik, B.; Zagula, G.; Bajcar, M.; Czernicka, M.; Puchalski, C. Biochar and Biomass Ash as a Soil Ameliorant: The Effect on Selected Soil Properties and Yield of Giant Miscanthus (Miscanthus × giganteus). Energies 2018, 11, 2535. [Google Scholar] [CrossRef] [Green Version]
- FAO. World Reference Base for Soil Resources 2014: International Soil Classification Systems for Naming Soils and Creating Legends for Soil Maps (Update 2015); World Soil Resources: Rome, Italy, 2014. [Google Scholar]
- DSTU 4287:2004. Soil Quality. Sampling; DP “UkrNDNC”: Kyiv, Ukraine, 2005; p. 9. [Google Scholar]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy Metals in Agricultural Soils of the European Union with Implications for Food Safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Dstu Iso 11464:2007. Soil Quality. Pretreatment of Samples for Physico-Chemical Analyses; DP “UkrNDNC”: Kyiv, Ukraine, 2009; p. 12. [Google Scholar]
- Kvak, V. Optimization of Elements of Technology for Production Miscanthus for Energy Fuel in the Western Part of Ukrainian Forest-Step. Ph.D. Thesis, Institute of Bioenergy Crops and Sugar Beets, Kyiv, Ukraine, 2014. [Google Scholar]
- DSTU 7688:2015. Diesel Fuel Euro; DP “UkrNDNC”: Kyiv, Ukraine, 2015; p. 16. [Google Scholar]
- GOST 9965-76. Petroleum for Oil-Processing Plants. Specifications; Standardinform: Moscow, Russia, 1977; p. 82. [Google Scholar]
- Wyszkowska, J.; Kucharski, J.; Waldowska, E. The Influence of Diesel Oil Contamination on Soil Microorganisms and Oat Growth. Rostl. Vyrob. 2002, 48, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Houben, D.; Evrard, L.; Sonnet, P. Beneficial Effects of Biochar Application to Contaminated Soils on the Bioavailability of Cd, Pb and Zn and the Biomass Production of Rapeseed (Brassica Napus L.). Biomass Bioenergy 2013, 57, 196–204. [Google Scholar] [CrossRef]
- MA. Announcement of the Ministry of Agriculture Related Requests to the Fertilizers; Ministry of Agriculture: Prague, Czech Republic, 2000; Volume N474/2000. [Google Scholar]
- DSTU EN ISO/IEC 17065. Issued by the System for Environmental Certification and Ecolabelling; DP “UkrNDNC”: Kyiv, Ukraine, 2016; Volume Certificate 10156. [Google Scholar]
- DSTU 4289:2004. Soil Quality. Methods for Determination of Organic Matter; DP “UkrNDNC”: Kyiv, Ukraine, 2005; p. 18. [Google Scholar]
- DSTU 4725:2007. Soil Quality. Potassium, Ammonium, Nitrate and Chloride Ion Activity Determination by Potentiometeric Method; DP “UkrNDNC”: Kyiv, Ukraine, 2008; p. 22. [Google Scholar]
- DSTU 4115:2002. Soils. Determination of Mobile Phosphorus and Potassium Compounds by the Modified Chirikov’s Method; DP “UkrNDNC”: Kyiv, Ukraine, 2003; p. 12. [Google Scholar]
- DSTU 7861:2015. Soil Quality. Determination of Exchanges Calcium, Magnesium, Sodium and Potassium in Soil According to Shollenberger in NSC ISSAR Named after Sokolovsky Modification; DP “UkrNDNC”: Kyiv, Ukraine, 2016; p. 12. [Google Scholar]
- DSTU ISO 10390:2001. Soil Quality. Determination of pH.; DP “UkrNDNC”: Kyiv, Ukraine, 2002; p. 14. [Google Scholar]
- Krause, G.H.; Weis, E. Chlorophyll Fluorescence as a Tool in Plant Physiology. Photosynth. Res. 1984, 5, 139–157. [Google Scholar] [CrossRef]
- Malinská, H.; Pidlisnyuk, V.; Nebeská, D.; Erol, A.; Medžová, A.; Trögl, J. Physiological Response of Miscanthus × giganteus to Plant Growth Regulators in Nutritionally Poor Soil. Plants 2020, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A Tool for Large-Scale Plant Phenotyping Connected to the Open PhotosynQ Network. R. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef] [Green Version]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The Relationship between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence. Biochim. Biophys. Acta (BBA) 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Hunt, E.R.; Daughtry, C.S.T. Chlorophyll Meter Calibrations for Chlorophyll Content Using Measured and Simulated Leaf Transmittances. Agron. J. 2014, 106, 931–939. [Google Scholar] [CrossRef] [Green Version]
- Tietz, S.; Hall, C.C.; Cruz, J.A.; Kramer, D.M. NPQ(T): A Chlorophyll Fluorescence Parameter for Rapid Estimation and Imaging of Non-Photochemical Quenching of Excitons in Photosystem-II-Associated Antenna Complexes. Plant Cell Environ. 2017, 40, 1243–1255. [Google Scholar] [CrossRef]
- Monti, A.; Zatta, A. Root Distribution and Soil Moisture Retrieval in Perennial and Annual Energy Crops in Northern Italy. Agric. Ecosyst. Environ. 2009, 132, 252–259. [Google Scholar] [CrossRef]
- Ben Fradj, N.; Rozakis, S.; Borzęcka, M.; Matyka, M. Miscanthus in the European Bio-Economy: A Network Analysis. Ind. Crop. Prod. 2020, 148, 112281. [Google Scholar] [CrossRef]
- Wagner, A.; Kaupenjohann, M. Biochar Addition Enhanced Growth of Dactylis Glomerata L. and Immobilized Zn and Cd but Mobilized Cu and Pb on a Former Sewage Field Soil. Eur. J. Soil Sci. 2015, 66, 505–515. [Google Scholar] [CrossRef]
- Luo, Y.; Lin, Q.; Durenkamp, M.; Dungait, A.; Brookes, P. Soil Priming Effects Following Substrates Addition to Biochar-Treated Soils after 431 Days of Pre-Incubation. Biol. Fertil. Soils 2017, 53, 315–326. [Google Scholar] [CrossRef]
- Tikhonenko, D.; Gorin, M.; Laktionov, M.; Kanivec, V.; Medvedev, V.; Baluk, S.; Buligin, S.; Truskaveckii, R.; Kanash, O.; Degtyar’ov, V.; et al. Soil Science: Textbook; Tikhonenko, D., Ed.; Higher Education: Kyiv, Ukraine, 2005; ISBN 966-8081-37-4. [Google Scholar]
- Gospodarenko, G. Agrochemistry: Textbook; Agrarian education: Kyiv, Ukraine, 2013; ISBN 978-966-669-317-7. [Google Scholar]
- Cadoux, S.; Riche, A.B.; Yates, N.E.; Machet, J.-M. Nutrient Requirements of Miscanthus × giganteus: Conclusions from a Review of Published Studies. Biomass Bioenergy 2012, 38, 14–22. [Google Scholar] [CrossRef]
- Babadzanova, O.; Sukach, Y.; Sukach, P. Phytotoxicity of soils contaminated by diesel. Sci. Bull. Natl. For. Tech. Univ. Ukr. 2017, 27, 92–94. [Google Scholar]
- Alkorta, I.; Garbisu, C. Phytoremediation of Organic Contaminants in Soils. Bioresour. Technol. 2001, 79, 273–276. [Google Scholar] [CrossRef]
- Shukry, W.; Al-Hawas, G.; Al-Moaik, R.; El-Bendary, M. Effect of Petroleum Crude Oil on Mineral Nutrient Elements and Soil Properties of Jojoba Plant (Simmondsia chinensis). Acta Bot. Hung. 2013, 55, 117–133. [Google Scholar] [CrossRef]
- Rahbar, F.G.; Kiarostami, K.; Shirdam, R. Effects of Petroleum Hydrocarbons on Growth, Photosynthetic Pigments and Carbohydrate Levels of Sunflower. J. Food Agric. Environ. 2012, 10, 773–776. [Google Scholar]
- Saeed, M.; Ilyas, N. Toxicity of Soil Hydrocarbon Pollution in Field Crops and Its Remediation. In Agronomic Crops: Volume 3: Stress Responses and Tolerance; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020; pp. 603–611. ISBN 9789811500251. [Google Scholar]
- Khan, S.; Afzal, M.; Iqbal, S.; Khan, Q.M. Plant–Bacteria Partnerships for the Remediation of Hydrocarbon Contaminated Soils. Chemosphere 2013, 90, 1317–1332. [Google Scholar] [CrossRef]
- Merkl, N.; Schultze-Kraft, R.; Infante, C. Phytoremediation in the Tropics—The Effect of Crude Oil on the Growth of Tropical Plants. Bioremediation J. 2004, 8, 177–184. [Google Scholar] [CrossRef]
- Kvak, V.; Hanzhenko, O.; Humentyk, M.Y.; Tsvihun, H. Relationship between leaf area, aboveground mass and morphometric indices of Miscanthus. Sci. Pap. Inst. Bioenergy Crop. Sugar Beets 2018, 26, 75–83. [Google Scholar] [CrossRef]
- Kvak, V.; Hanzhenko, O.; Zykov, P.; Khivrych, O. Method for determination of leaf area in different sorts of Miscanthus by calculation. Adv. Agrotechnol. 2017, 5, 7. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Gómez, S.; Petenello, M.C.; Feldman, S.R. Growth, Nutrient Status, and Photosynthetic Response to Diesel-Contaminated Soil of a Cordgrass, Spartina argentinensis. Mar. Pollut. Bull. 2014, 79, 34–38. [Google Scholar] [CrossRef]
- Baruah, P.; Saikia, R.R.; Baruah, P.P.; Deka, S. Effect of Crude Oil Contamination on the Chlorophyll Content and Morpho-Anatomy of Cyperus brevifolius (Rottb.) Hassk. Environ. Sci. Pollut. Res. 2014, 21, 12530–12538. [Google Scholar] [CrossRef]
- Loggini, B.; Scartazza, A.; Brugnoli, E.; Navari-Izzo, F. Antioxidative Defense System, Pigment Composition, and Photosynthetic Efficiency in Two Wheat Cultivars Subjected to Drought. Plant Physiol. 1999, 119, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Bielczynski, L.W.; Łącki, M.K.; Hoefnagels, I.; Gambin, A.; Croce, R. Leaf and Plant Age Affects Photosynthetic Performance and Photoprotective Capacity. Plant Physiol. 2017, 175, 1634–1648. [Google Scholar] [CrossRef]
- Kandel, B.P. Spad Value Varies with Age and Leaf of Maize Plant and Its Relationship with Grain Yield. BMC Res. Notes 2020, 13, 475. [Google Scholar] [CrossRef]
- Han, G.; Cui, B.X.; Zhang, X.X.; Li, K.R. The Effects of Petroleum-Contaminated Soil on Photosynthesis of Amorpha fruticosa Seedlings. Int. J. Environ. Sci. Technol. 2016, 13, 2383–2392. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of QA Redox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209. [Google Scholar] [CrossRef]
- Netto, A.T.; Campostrini, E.; de Oliveira, J.G.; Bressan-Smith, R.E. Photosynthetic Pigments, Nitrogen, Chlorophyll a Fluorescence and SPAD-502 Readings in Coffee Leaves. Sci. Hortic. 2005, 104, 199–209. [Google Scholar] [CrossRef]
- Maier, R.M.; Gentry, T.J. Chapter 17—Microorganisms and Organic Pollutants. In Environmental Microbiology, 3rd ed.; Pepper, I.L., Gerba, C.P., Gentry, T.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 377–413. ISBN 978-0-12-394626-3. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agrochemical Parameters | Unit | Mean ± SD |
|---|---|---|
| pH (KCl) | 6.46 ± 0.03 | |
| Organic matter | % | 1.13 ± 0.04 |
| NO3 | mg kg−1 | 126 ± 9 |
| NH4 | mg kg−1 | 1.7 ± 0.2 |
| P2O5 | mg kg−1 | 176 ± 12 |
| Exchangeable K | mg kg−1 | 116.9 ± 13.2 |
| TEs | MPC, mg kg−1 | TEs Concentration in the Soil, mg kg−1 | |
|---|---|---|---|
| EC [35] | Ukraine [34] | ||
| Mn | NA | 1500 | 628.3 ± 44.4 |
| Fe | NA | 22,073 ± 129.6 | |
| Cu | 100 | 55 | 17.0 ± 7.5 |
| Zn | 200 | 300 | 49.4 ± 5.7 |
| Pb | 60 | 30 | 25.1 ± 3.5 |
| TEs | B1, mg kg−1 | B2, mg kg−1 |
|---|---|---|
| Mg | 13,139.6 ± 1622.4 | 14,224.3 ± 2122.1 |
| Al | 38,762.9 ± 767.3 | 22,223.8 ± 977.8 |
| Si | 141,728.1 ± 726.8 | 272,941.6 ± 1692.0 |
| P | 106,458.1 ± 919.8 | 46,767.8 ± 1002.0 |
| S | 15,711.4 ± 116.1 | 7510.8 ± 133.4 |
| K | 10,677.8 ± 1552.9 | 60,664.0 ± 5726.2 |
| Ca | 94,085.4 ± 1143.1 | 71,290.6 ± 3621.5 |
| Ti | 4811.5 ± 359.5 | 4906.0 ± 927.5 |
| Cr | 453.0 ± 94.5 | - |
| Mn | 896.5 ± 72.4 | 821.4 ± 208.3 |
| Fe | 123,923.4 ± 452.1 | 35,621.2 ± 411.8 |
| Ni | 137.4 ± 28.1 | 78.9 ± 48.9 |
| Cu | 627.2 ± 24.2 | 152.1 ± 36.0 |
| Zn | 6847.5 ± 54.9 | 727.2 ± 36.4 |
| Sr | 485.0 ± 6.3 | 554.2 ± 12.9 |
| Zr | 260.6 ± 5.7 | 662.3 ± 14.8 |
| Sn | 57.4 ± 19.6 | - |
| Pb | 100.0 ± 8.4 | 81.9 ± 15.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pidlisnyuk, V.; Herts, A.; Khomenchuk, V.; Mamirova, A.; Kononchuk, O.; Ust’ak, S. Dynamic of Morphological and Physiological Parameters and Variation of Soil Characteristics during Miscanthus × giganteus Cultivation in the Diesel-Contaminated Land. Agronomy 2021, 11, 798. https://doi.org/10.3390/agronomy11040798
Pidlisnyuk V, Herts A, Khomenchuk V, Mamirova A, Kononchuk O, Ust’ak S. Dynamic of Morphological and Physiological Parameters and Variation of Soil Characteristics during Miscanthus × giganteus Cultivation in the Diesel-Contaminated Land. Agronomy. 2021; 11(4):798. https://doi.org/10.3390/agronomy11040798
Chicago/Turabian StylePidlisnyuk, Valentina, Andriy Herts, Volodymyr Khomenchuk, Aigerim Mamirova, Oleksandr Kononchuk, and Sergey Ust’ak. 2021. "Dynamic of Morphological and Physiological Parameters and Variation of Soil Characteristics during Miscanthus × giganteus Cultivation in the Diesel-Contaminated Land" Agronomy 11, no. 4: 798. https://doi.org/10.3390/agronomy11040798