
Is Domestication Speciation? The Implications of a Messy Domestication Model in the Holocene


Department of Anthropology, Penn Museum, University of Pennsylvania, Philadelphia, PA 19104, USA
Agronomy 2021, 11(4), 784; https://doi.org/10.3390/agronomy11040784
Submission received: 11 March 2021
/
Revised: 9 April 2021
/
Accepted: 14 April 2021
/
Published: 16 April 2021
(This article belongs to the Special Issue Histories of Crops, between Niche Construction, Domestication and Diversification)
{kind=link}
{kind=link}
Abstract
:Domestication is one of the fundamental process that has shaped our world in the last 12,000 years. Changes in the morphology, genetics, and behavior of plants and animals have redefined our interactions with our environments and ourselves. However, while great strides have been made towards understanding the mechanics, timing, and localities of domestication, a fundamental question remains at the heart of archaeological and scientific modelling of this process—how does domestication fit into a framework of evolution and natural selection? At the core of this is the ontological problem of what is a species? In this paper, the complicated concepts and constructs underlying ‘species’ and how this can be applied to the process of domestication are explored. The case studies of soybean and proto-indica rice are used to illustrate that our choice of ‘species’ definitions carries with it ramifications for our interpretations, and that care needs to be made when handling this challenging classificatory system.
1. Introduction: What Is Domestication?
The intricacies of how humans, animals, and plants became so entangled in their day-to-day interactions has been a question that long puzzled archaeologists and evolutionary biologists alike. Darwin devoted not only sections of his more famous Origins manuscript [1], but an entire two volumes to the investigation of The Variation of Animals and Plants Under Domestication [2]. From an archaeological standpoint, domestication became entangled with ideas of agricultural revolutions (see for example [3], and the search for the origins/centers/hearths of domestication events (following on from Vavilov’s [4] seminal work). In more recent years, definitions of ‘domestication’ have moved away from thinking about it as an event towards recognition of the temporality and complexity of the processes involved.
Fuller and Hildebrand [5] suggest domestication should be seen as “genetic and morphological changes on the part of the plant [and animal] population in response to selective pressures imposed by cultivation” (see also Langlie et al. [6], who adopt this definition). This definition of domestication therefore emphasizes changes in the domesticate caused by/responding to human action. A similar definition is made by Larson et al. [7] (p. 6140), who note that domestication “can generally be considered as a selective process for adaptation to human agro-economic niches and, at some point in the process, human preferences”. They go on to stress that these responses lead the plant/animal to “express traits that were favorable to human use, harvesting and edibility”. Again, human action and plant/animal change in response are emphasized in this definition, along with the notion of the long-term temporality of domestication processes. Similar definitions of the human–plant, human–animal interaction leading to change can be found in Lee et al. [8], Zeder et al. [9], Gepts et al. [10], and Zohary et al. [11]; though see Ingold [12].
An important aspect that all recent definitions of domestication highlight is the protracted temporality of the process of domestication. Larson et al. [7] comment on the length of many pre-domestication cultivation periods, and how domestication traits may not have appeared all at the same time in different floral and faunal taxa exploited. Data from archaeological sites in works by Hillman and Davis [13], Willcox and Strouder [14] and Yang et al. [15] illustrate this. Fuller [16] notes that domesticates “may evolve over thousands of generations, as desirable traits are selected for in the agricultural environment and become fixed within the crop genome” (cited in Meyer et al. [17]). In citing this, Meyer et al. [17] also note that some domesticates have been altered from their wild form over shorter periods, providing kiwi and cranberry as examples, but also comment that these would still be in the hundreds of years’ time span for domestication to occur, placing them, as Fuller and Stevens [18] note, outside the lifespan of the individual (plant, animal, and human). It can also be noted that polyploidy and genome duplication can lead to domestication, often more rapidly than the processes envisioned in the definitions and models of Fuller [16], Larson et al. [7], and Meyer et al. [17]. This is seen particularly in long-generation perennials, where self-incompatibility leads to higher rates of interspecific hybridization [19].
The question then becomes how can the changes associated with the various forms of domestication, the majority of which occurred in the distant past, be identified? In modern plants and animals and in archaeological remains, domesticates are usually phenotypically distinctive, and as such, these distinct distinguishing traits are used to delineate wild from domestic taxa. This “domestication syndrome” [20,21] has provided the mainstay of much archaeological work looking not only for domesticates, but also for the process of domestication—where do we see them being used, and what do they look like and when (see for example [6,7,17,20,21])?
Domestication syndrome traits are those that, as Larson et al. [7] put it, “were favorable for human use, harvesting and edibility”. Examples include loss of seed dispersal mechanisms without human intervention (seen in cereals and pulses [11,22]), reduction of physical and chemical defenses (loss of awns in cereals [23]; loss of toxicity in almonds [24]; reduction in horns in cattle [25]), endocrine changes (seen in guinea pigs [26]); facial neoteny (though there is some debate about this [27]), and reduced astringency and seedlessness (in tree fruits [18]). These are but a few of the domestication syndrome traits, and not all cereals will have the same traits, nor all pulses the same traits, etc., and parallel lines of evolution can and do exist. Both similarity and variation within and between plant/animal ‘groups’ is seen.
Additionally, this has led to two related problems—the first, as Purugganan and Fuller [28] note, has been a debate within evolutionary biology and history of science of how do we fit domestication into a framework of evolution and natural selection? Additionally, the second issue is tied to this. As Lee et al. [8] comment, “the differences between domesticated plants [and by extension animals] and their wild relatives are not always clear”. They make this point specifically in relation to archaeological specimens and preservation concerns, but it echoes broader debates within biology about the relationship between domesticates, wild taxa, and the problem of speciation in the evolutionary framework, for example within hominin anthropology [29,30,31], and to discussions about how to apply typologies and classificatory schema within the broad spans of time that archaeology addresses, e.g., [32,33,34].
2. How Does ‘Domestication’ Relate to Evolutionary Models?
Contemporaries of Darwin questioned his use of the artificial selection analogy for natural selection. Wallace [35] in a volume containing Darwin’s work critiqued the analogy. Concern over the different pathways for species divergence and differentiation that natural and artificial selection take has dogged the literature based on the use of analogy. Darwin’s [1,35] discussions of natural selection drew heavily on his observations taken from plant and animal breeders, and his own experiments, but how this analogy formed and was used throughout his work has become more of a philosophical debate than a scientific one (see for example [36].)
Darwin [2] describes three forms of change building on the variation in nature—natural selection, methodical selection, and unconscious selection (though in later works he adds in sexual selection explicitly as a concept). The latter two—methodical and unconscious selection—are traditionally grouped under artificial selection. Gregory [37] suggest that while these all differ based on the reason why change occurs, there is still a fundamental mechanistic similarity of process within them that has been refined since the rediscovery of the laws of Mendelian inheritance [38].
Gregory [37] argues that despite concerns relating to the intent of Darwin in his use of analogy, there remains useful lessons in artificial selection analogies. As well as laying out suggestions relating to evolutionary studies, Gregory [37] highlights in particular how selection, both natural and artificial, leads to profound changes. Finches, dogs, and maize are all highlighted as examples, and the morphological, behavioral, and genetic differences between various forms (word chosen carefully here) are noted.
Additionally, this is where the second point outlined above—the question of the clarity of difference between wild and domesticated plants and animals—comes into play. Morphologically, behaviorally, and genetically varieties of finches, dog, and maize are easy to distinguish when comparing a start and end point. When looking at a domestication process, however, this becomes more complex.
Meyer et al. [17] highlight this in trying to draw up definitions. They layout a term for a thing that is ‘domesticated’: “morphologically and genetically distinct from their wild ancestors as a result of artificial selection, or are not known to occur outside of cultivation”. They outline ‘undomesticated’ things: “uncultivated plants that continue to be wild-harvested with no conscious artificial selection pressures and no discernable morphological and/or genetic differentiations that could be used to distinguish them as a domesticate”. Aside from concerns relating to the consciousness of all domestication processes implied in this definition, it sets two boundaries—a wild end of the spectrum and a domesticated end. However, as our definitions of domestication above have highlighted, this is a prolonged (hundreds to thousands of years) process on a continuum of human interaction with plants and animals. Meyer et al. [17] suggest ‘semi-domesticated’ as a term for the bit in the middle, and define plants (and by extension animals) in this period as “under cultivation and subject to conscious artificial selection pressures. Although named cultivars may exist, these are not yet morphologically or genetically distinct from their wild counterparts”. Again, however, this implies that when change occurs, it is sudden: a long period of pre-domestication cultivation of things that are morphologically and genetically undomesticated then suddenly transform into phenotypically domesticated things. There is an additional irony to the term ‘semi-domesticated’: it implies not only a start and end point of domestication, but also that the process is goal orientated. “Semi” emphasizes that underlying many archaeological and genetic definitions of domestication is a suggestion that domestication is separate from ‘natural’ processes, from evolution and evolutionary theory, because humans are involved. However, domestication is still a part of evolution, and other form of the changes that form evolution, and like evolution, domestication must be seen as continual, non-linear, and non-discrete, and subject to the passage of time, without the benefit of hindsight by/about the human and non-human actors involved (perhaps reminiscent of the definitions in Ingold [12]).
However, this leaves us with a challenge: how are we to incorporate an evolutionary model of domestication within discussions change between one ‘thing’ (the wild taxa) at the start of the process and the ‘thing’ (domesticated taxa) it becomes? What are we to do with the ‘things’ between undomesticated and domesticated? Additionally, are the taxa at the start and end truly different: does domestication lead to ‘speciation’?
3. Does ‘Domestication’ Lead to Speciation?
The role of speciation as process in domestication has oft been overlooked, despite the debates surrounding artificial and natural selection analogies. Domestication scholars require some form of naming classification system to describe their data. It is not necessarily the place of archaeobotanists or zooarchaeologists to question agreed on conventions of taxonomy or naming systems. However, we need to recognize that the decisions to define an organism, including domesticated organisms, as different genera, species, and subspecies is complex and not inherent to the organism itself. Instead, it is affected by our own subjective decisions about how to create or apply classificatory systems. How we define ‘species’ has ramifications to our modelling, particularly when thinking about change over time. Discussed in the archaeological domestication literature is the idea that a plant or animal undergoes a process that leads to it changing from wild species x to become domesticated species y, without much consideration of what these species mean.
‘Species’ is a much debated construct within evolutionary and biological literature, with early discussions in Darwin and Wallace [1,2,35] expanded on by Mayr [39,40], Simpson [41], and Templeton [42], for example. More recent debates have expanded on these to think about the underlying ontological constructs of ‘species’. Far from the simple high-school notion of reproductive isolation [43], scholars such as Wilkins [44] and Mayden [45] have identified as many as 27 different definitions of ‘species’ in use by biologists, with likely many more underlying variants. De Queiroz [46,47] and Hey [48] have termed this the ‘species problem’—that there are too many competing definitions, and no underlying agreement on what a species is and how and why species are being defined. This complexity of ‘species’, and thus of speciation within the evolutionary biology, plays into the broader intricacies of a model of archaeological domestication—if we cannot define what a species is, how can we understand the process by which humans have been involved in creating one (even if that process was unconscious, mutualistic, or commensal)? How can we discuss the evolutionary processes without consideration of evolutionary classificatory debates?
An example of how the ontological tangles of ‘species’ in evolutionary theory can muddy the waters of archaeological discussion is beautifully outlined in the terminological convolutions of Lee et al. [8]. The strictures of using a reproductive isolation model of species leads them to complications when trying to understand how wild and domesticated soybean are related. As Lee et al. [8] comment “domesticated plants are usually phenotypically distinctive and, because they are also cultivated, taxonomists often consider them to be separate species” from their wild counterparts. However, while niche difference [49] and some limited physical differences [8] are apparent between wild and domesticated soybean, at a fundamental level, these can still cross breed and produce fertile F1 offspring [20]. This leads Lee et al. [8] to the fundamental problem that domesticated soybean and its wild relative should be classified as subspecies of one another if we follow a reproductive isolation model to define species [20], despite the apparent ecological and phenotypic differences between these two taxa.
In order to find a way around these naming and definition challenges, they adopt a similar classificatory system to that seen in Harlan and de Wet [50], a system that works from the ‘known’, the domesticated and supposed ‘final’ form of a plant, backwards to the unknown/less well known, the wild progenitors. While this may be logical in many respects—as archaeologists we are always looking from the present backwards with hindsight—it fails to take in the evolutionary trajectories of plants, the evolutionary nature of domestication as branching, non-linear, and complex. It leads to wild subspecies ancestors being defined on the basis that they lack domesticated traits, places a burden on us to find a start and end point to domestication, and creates an artificially smooth path between the wild and domesticated. Such an approach also results in clunky trinomials, a problem that Harlan [20,51] did try to address, but continued to be of concern throughout this system. For Lee et al. [8], this resulted in the slightly complex situation whereby the domesticated soybean Glycine max and its wild relatives G. soja are instead named as G. max subsp. max and G. max subsp. soja, respectively. Wild soybean, the ancestor, is relegated to the realm of a subspecies of the domesticate it eventually becomes, and a linear path between the two is presumed. Additionally, this added further complications, as the archaeological record is not always clear and identifications can often only be made to genus level rather than species, let alone to subspecies [8], and Lee et al. [8] noted that their samples were not perfectly preserved to reach such levels of identifications.
To try to handle this species/subspecies-based chaos, and the ensuing wild/domesticated discussion, Lee et al. [8] decide to avoid the taxonomic debate as much as they can, and state “Throughout this paper we use the term ‘‘soybean’’ to refer to G. max without specifying the subspecies because of the ambiguity in the archaeological record. We do not wish to bias the terminology to favor either wild or domesticated soybean” [8].
This is an excellent example of the ontological challenges posed by the way ‘species’ is used in archaeological domestication studies: the terms we use, and the schemes we settle on, have real ramifications for our data, narratives, and interpretations.
The problem arises from our attempts to use species as both a unit of identification and delineation of processes. In taxonomic terms, we might argue it is simply there to help classify the world. However, species takes on a more active role in domestication discussions, as behind the labels are concepts, mechanisms of change, and notions of the relationship between the ‘species’, its ancestors, and the surrounding world in which it is found.
We are trying to define a concept that has many definitions and ontological discussions already behind it [44]. Species as based on simply reproductive isolation fails to take into account ecology or phylogeny or seed dispersal mechanisms (for example), but using these alternative systems as bases for a definition also fails to take other aspects such as phenotypic expression and morphology into account as well. Hey [48] suggests that at the heart of the problem are two completing ideas: species as taxa and species as evolutionary groups. On the one hand we have something that is a category, discrete in itself, and on the other something that is somewhat discrete but capable of changing [48]. One is a classification scheme, the other an explanatory model. This creates underlying problems of communication across biology. Add in additional nuances of how we define a species and the differences in these approaches increases.
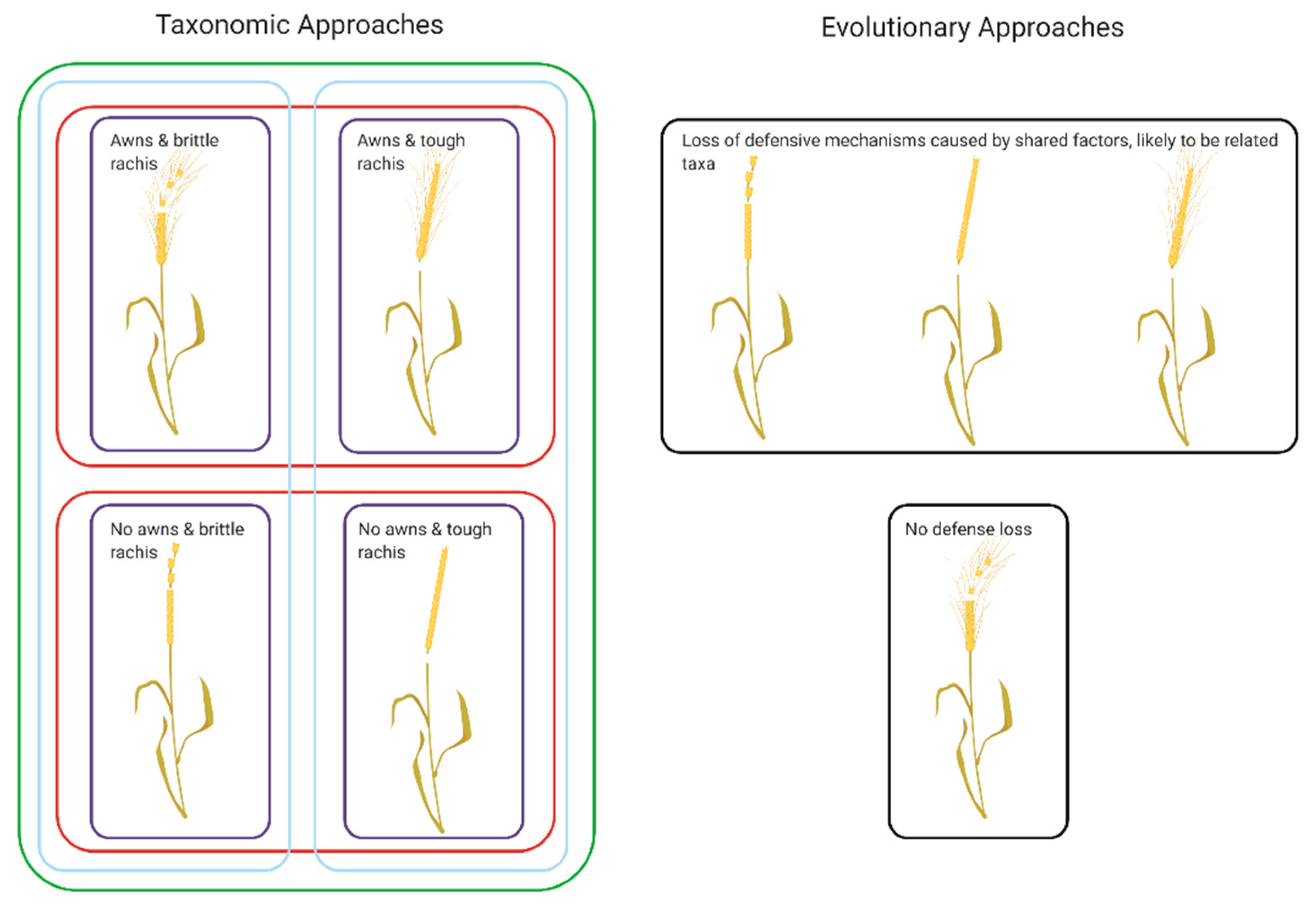
Within the taxonomic definitions, categorization is created through perception—we might say this cereal looks different from this cereal because of the lack of awns, the tough rachis, etc. Under an evolutionary model, however, these changes in awns presence and rachis toughness may not be useful because they may or may not be ‘distinct’ at a genetic level or linked through relatedness [48]. Finally, Hey [48] argues, species taxa and evolutionary groups are formed for different motivations—we create taxa for convenience (separating our hypothetical cereal into different types for our own visual use for example), while evolutionary groups are created for their relationships to one another (our hypothetical cereal evolved from this one while this cereal is a hybrid of a completely different set of cereals within the same genera and maybe some completely different ones) regardless of how we perceive them to look (Figure 1).
If we look to wheat we can see this playing out [11,17,52,53]. Through the study of cytogenetic and molecular affinities, it has been recognized that one of the key steps in wheat domestication was rapid domestication through polyploidy leading to speciation and genetic incompatibility [11,53]. As a result of this, we see in Zohary et al. [11] a comparison of taxonomies based primarily on morphological differences (species as taxa groups approach as defined by Hey [48]) and taxonomies based on genetic phylogeny (species as evolutionary groups approach following Hey [48]). Hey [48] would argue that both could be equally valid, providing that the underlying ontological frameworks are explicitly explored and understood. Most archaeobotanists these days tend to follow the phylogenetic taxonomy, reasoning this must be the most up-to-date schema of ‘relatedness’. It might be argued then that the reasoning behind choosing which taxonomic classification to follow is not commonly expanded on in the archaeobotanical domestication literature, nor the implications of choosing a definition of different ‘species’ explored within domestication studies of wheat.
Turning back to Lee et al. [8] the struggle between how to apply classificatory naming schema and the proposed evolutionary relatedness of the studied taxa caused difficulties that had the potential to derail the domestication process modelling. Rather than relying on a discrete model of biological incompatibility and a naming system that then led them to utilize a linear model of change from one form to another, Lee et al. [8] could instead have gone down the route favored by Wilkins [44] and outlined any one of the many definitions of species available to them to explain their model in a more flexible fashion. Perhaps for example Ecospecies or maybe simply stating that they were using Taxonomic species could suffice to sidestep the reproductive isolation concerns? One pushes the debate down an evolutionary route, the other down a taxa route, but both remain valid (though Hey [48] might take issue with the latter). Choosing either would have resulted in a very different paper. This was sidestepped by a careful exploration of the paradigm underlying their decisions, and preservation concerns for the remains would make deciding on either route very difficult. The authors’ decision to focus on the data instead provided a thorough study of the changes between wild and domesticated soybean over time by doing so, but as a result missed some of the key questions of both taxonomy and evolutionary change during the domestication process.
This also highlights that we need to consider how our attempts to understand species often take on a methodology and identification tone—trying to explain how we have identified something as a species rather than what was actually going on in the past. Wilkins [44] argues that “the tests we use to find out whether two organisms are the same species or not may in fact be quite incidental to the causes of their being species”. This can be seen in the debates across archaeology regarding the timing of genetic change and morphological expression, pinpointing the moment when something ‘becomes domesticated’ based on one dataset or another. In the case of the soybean, the authors deployed a multi-stranded approach to ask the question “How does soybean seed size inform us about the domestication of this crop?” [8], but at times let the human and non-human actor angle slip from the story, for example stating that human interactions with the plant “seems to be the result of an accumulation of minor changes at quantitative trait loci (QTL)” [8]. While this provides a what and how domestication occurred (gene change), it does not provide a why, nor the story of the human–plant interaction behind it.
Therefore, do we junk the idea of species in our attempts to model domestication, assume that it is too complex and burdened by underlying ideas to use, or go down the route of Hey [48] in accepting that a species is nothing more than an idea in the mind of the collector or accessioner? (Additionally, this needs to be carefully accompanied by a definition much in the way domestication, agriculture, and cultivation are—‘warning this paper contains species which are defined as…’). To do so seems fraught with complexity, but could help avoid the sticky situation that the case study below outlines. On the other hand, as also outlined below, nuanced definitions of ‘species’ in the framework of a domestication story can illuminate the complexity of domestication, and show that a rigid definition of ‘species’ leaves no room for human and plant agency, ecological niche restrictions, and the role of time in the process of domestication.
4. Domestication and Proto-Indica
Modern domesticated rice, Oryza sativa, has a complex history, as it is the product of repeated instances of hybridization. Rice has been chosen here not because it is special, but simply because the proto-indica hypothesis in many ways is illustrative of the domestication-species conundrum. Similar problems are encountered in other taxa stories. As noted in the history of Chenopodium sp. by Langlie [54], there has been a tendency to “present modern specimens as a sort of idealized morphological type that all archaeological specimens are evolving towards or should conform to”. This has led to linear models in which there is a start point, species a, and an end point, species b. However, how then do we handle situations when “archaeological seeds do not conform to these proportions […] are they fully domesticated” or something entirely different [54]? Rice archaeobotanists, geneticists, and modelers it seems are far from the only scientists to struggle with the idea of how to handle a fluid and dynamic process like domestication and how to classify individual ‘species’ finds within this spectrum.
Asian rice has a well-recognized genetic subdivision into indica-type rice, japonica-type, and aus-type rice [55]. Archaeological and genetic evidence suggested that japonica rice cultivation began in the Yangtze valley, China c.9000BP with the pre-domestication cultivation of wild O. rufipogon [56,57]. This led to domestication with several key genes becoming fixed in the population [57], but this was not the only domestication process [56,58], and rather hybridization between japonica and South Asian rice was necessary for the domestication of O. sativa.
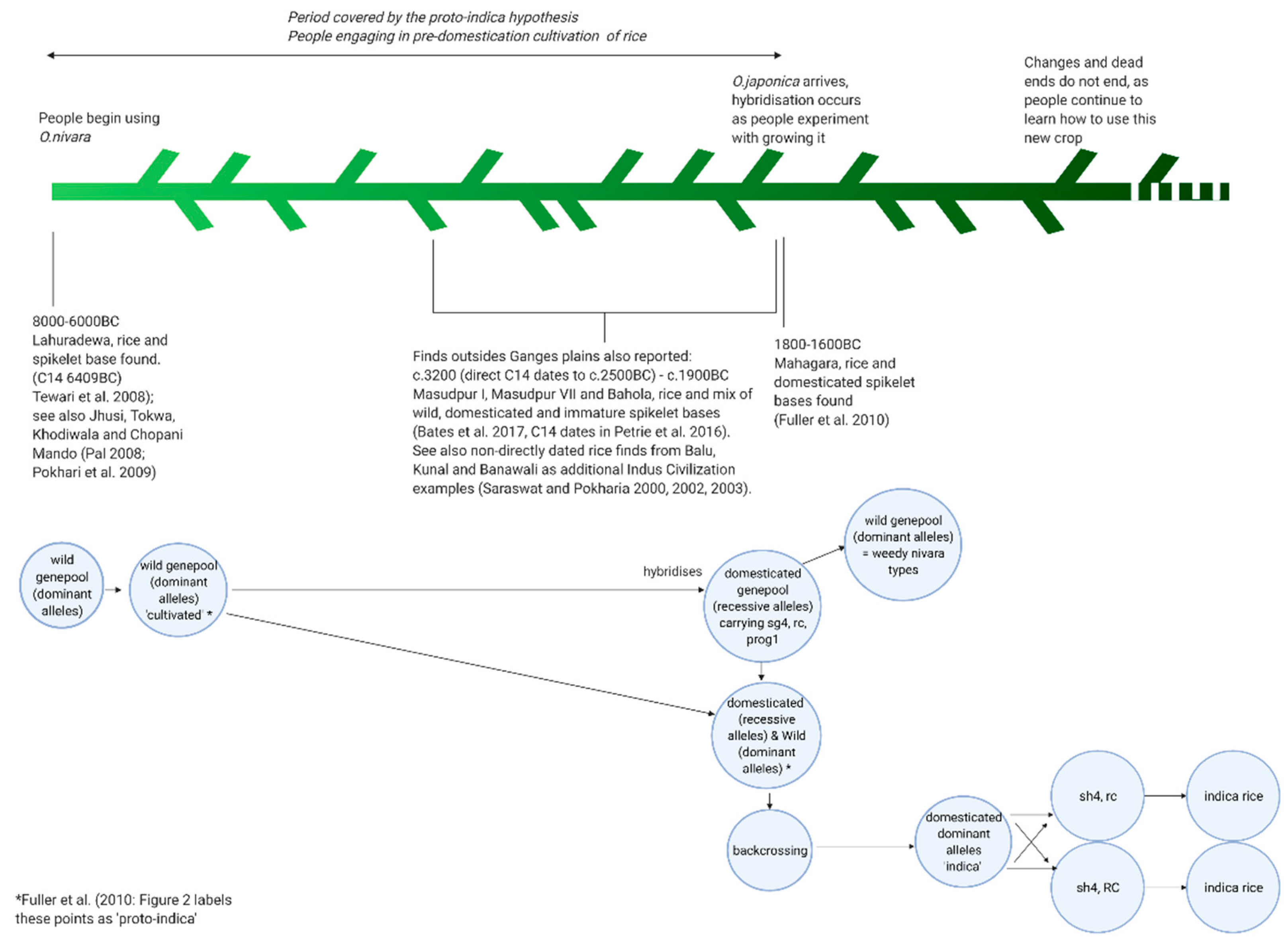
Fuller et al. [56], Fuller [59], and Silva et al. [58] explain this as the ‘proto-indica’ hypothesis. This hypothesis is summed up by Fuller [59] who argued that the “independent rice tradition in north India […] never […] proceeded on its own to full domestication” until the arrival of O. sativa ssp. japonica c.2000 BCE. The proto-indica hypothesis outlines a process by which domesticated japonica was introduced to the region of northern India where a semi-domesticated indica ancestor was present. This semi-domesticated ancestor, proto-indica as they term it, was under pre-domestication cultivation, and hybridized with the japonica. Through back-crossing and transference of domesticated traits, fully domesticated indica evolved [56,58,59].
However, one big question remains: what exactly is ‘proto-indica’? On the one hand, this is discussed as a hypothesis, a fluid part of a process that does not need to be pinned down, but is ever changing between the wild nivara and the eventual hybridization point with japonica when indica is formed. On the other, we have discussions of ‘proto-indica’ as the thing that hybridizes with japonica, a taxonomic and/or evolutionary species. The taxonomic element of the species definitions is therefore present in these models: proto-indica is a thing that is no longer nivara, but not yet indica (“proto-indica cultivars” [59]). An evolutionary species sense of proto-indica is present in the implications of the relationship between nivara—the thing it was—and indica—thing it will become. In both uses of the idea of ‘species’ proto-indica needs to be defined somehow, by characteristics of morphology, behavior, genetics (this may be feasible soon as extinct proto-indica lines are slowly being implied by the new chloroplast data emerging [59,60]), or some other quantifiable or qualifiable feature. Additionally, here comes a further problem—how many features does it take for nivara to become proto-indica?
Fuller et al. [56] and Silva et al. [58], for example, suggest that it is human behavior that makes proto-indica a taxonomic thing—proto-indica is no longer nivara because humans are treating it differently, placing it in the pre-domestication cultivation realm of behavior (or perhaps the ‘cultiwilds’ [61,62]). However, this itself raises another set of questions—should we now have proto-monococcum wheats? As Langlie [54] have noted, to do so “sets up a rigid unilinear model of domestication with an end goal in mind”, “distorted by a clear evolutionary trajectory”—these wild rice plants were always going to end up as indica, they are ‘proto’ in the prototype, precursor, preparator sense of the word. In this sense, the idea of proto-indica (or a hypothetical proto-monococcum) might be divided from a cultiwild banana [61,62], as the proto-indica rice is conceived as on a trajectory to domestication through the use of the term ‘proto’. Cultiwild bananas on the other hand are a cultivated but morphologically and/or genetically wild form, a term describing their use status or human interactions with the plant rather than prescribing their place on a pathway to physical change.
Clearly this is not what proponents of the proto-indica hypothesis intend—as Fuller [59], for example, has shown in the complexity of modelling rice domestication, there was no clear domestication pathway, with many dead ends and branches. However, the species-style use of proto-indica to discuss what is instead the pre-domestication cultivation of nivara rice complicates this and the process-type modelling of the proto-indica hypothesis (Figure 2).
One element that papers have brought up though is the use of the word ‘cultivars’ [59]. This concept is one that may at first appear to avoid the tricky subject of species, but again falls into the hot waters of categorization and classification that species does. While in its original form [64], cultivar and the related construct ‘cultigen’ may provide some sanctuary, providing a useful non-taxonomic structure of cultivated but not necessarily fully domesticated plants, in recent years the term has been refined. Cultivars are specific things, required to be stable, uniform, and distinct to gain their epithets [65]. Proto-indica (and other plants under domestication) were not stable by the very fact they were under domestication (see Fuller’s [59]: Figure 2 for examples of the branching in the model). Are we to have each minor change in gene or morphology to be labelled a new cultivar? When does a cultivar get changed to a new species in the fully domesticated sense? This taxonomic (for it is taxonomic at this point) problem is thus piled on top of the already muddied waters of the hypothesis–thing questions already formed by the process–species issues. Best to leave cultivars and cultigens for a more stable system.
Where then does this leave us? With an evolutionary domestication model in which the long drawn out and non-linear changes in nivara are created through pre-domestication cultivation human interaction with wild rice in South Asia in a branching set of rice ‘things’ over time until domesticated japonica is brought into South Asia and hybridization occurs, forming indica. However, also with a strange quasi-species floating around conceptually to try to help describe the cultivated, but not fully domesticated nivara, no longer wild, but not ‘yet’ indica. Are the semi-domesticated things species in the taxonomic or evolutionary sense? It depends how you define species, how many traits and changes, what changes these are, and whether these are quantifiable/qualifiable or reflationary as to where you want to draw the boundary between nivara and a ‘proto-indica’ species. Additionally, to recognize that in doing so could create a model “distorted by implying there is a clear evolutionary trajectory” (Langlie [54]) between nivara and indica without recognizing the complexity and intricacy of the proto-indica hypothesis (Fuller [59]: Figure 2).
5. Conclusions
Species in the contexts of domestication are therefore messy. That is no surprise—as Hey [48] tells us, species are problematic regardless the context. In the domestication story, however, they bring their own unique problems. By creating discrete categories, they force us to place pinpoints on a long drawn out process, to mark the specific times when change occurs. Subspecies, forms, ecotypes, varieties, morphotypes, and other classificatory terms akin to ‘species’ on the other hand all face the same challenges as ‘species’—a need to define and for clarity in how they fit with evolutionary schema when writing domestication narratives in archaeology. Species (and similar naming terms) suggest, implicitly or explicitly, that something definitive and temporally defined and refined has occurred, and in the context of domestication, that humans caused this. We create a static pattern in a fluid and non-linear story by-passing the cases where plant and animal domestication ‘dead-ends’ occurred. We also imply that changes in cultivation/pastoral behavior beyond that point were less impactful, they do not lead to the same ‘significant’ and category-making moments [54]. This is not unique to domestication. All evolution is non-linear and ‘fluid’, and many dead-ends occur. Domestication is simply another form of evolution, one that humans are simply a part of, and one with parallels in the wild. Given this, perhaps a strong take away of this argument is not only that we should be more aware of the underlying assumptions implicit in our classificatory terms, but also that we, as archaeologists exploring domestication narratives, should engage more with evolutionary biological theory.
Species are ‘created’ to explain the gradated changes between something recognized by taxonomists and geneticist as distinct, thus muddying the waters when trying to describe human action and plant/animal co-evolution with our behavior. The multi-species approach is perhaps one way to handle this, to accept that non-human actors are a pivotal part of the process of domestication and that human-centric approaches are not necessarily the entire picture even if suggested only implicitly (bringing us back to Ingold [12]; see also [66]). It is important in this messy picture, however, to remember that species should not be abandoned wholesale in our pursuit of a complex domestication story. Species as a way to categorize also serve to illustrate change throughout the story, to give us points to explore. Indeed, in doing so, we can take more into account, notions such as comparator moments between ancient domesticates and modern domesticates, how the evolutionary nature of domestication continues and does not have an end point (see work on Chenopodium sp. by [54,67,68]).
However, it is only when our underlying meaning of what ‘species’ is—be it taxonomic species, phylogenetic species, nothospecies, ecospecies, etc. [44], or a more simple distinction between the systems of Hey [48]—are clearly stated, that species can help us to reconsider whether we are looking for physical change in the object of study or changes in the relationships between things, and then to look at what this might mean in the human–plant/human–animal story being told. Terms like ‘proto’ should be abandoned, as they imply directedness, terms with modern burden like ‘cultigen’ likewise should be avoided unless there is due reason to use them, and explicit discussions or outlines of what we mean in our discussions by ‘species’ when thinking about domestication processes and pathways, or why one taxonomic system has been adopted over another (e.g.,: why Harlan and de Wet [50] rather than Zohary et al. [11]), need to be more regularly incorporated into our archaeological literature. A messy model of domestication therefore does not need to be a negative thing; the models we already use are inherently messy with their non-linear pathways and variable temporalities, a fact born out in the archaeobotanical, zooarchaeological, and (archaeo)genetic data. The ‘messiness’, however, needs to be expanded to incorporate more evolutionary perspectives and evolutionary biological theory. A truly messy, and thus more realistic, model is simply a recognition both of the way humans and non-human actors interacted in the past and of the challenging set of empirical facts that come with evolutionary theory.
Perhaps then it is useful to bear in mind in our introductions and conclusions the inherent subjectivity of our naming of ‘things’, and to avoid simplistically stating that a thing is a ‘species’ [48]. We categorize our world to make it easier to manage [69], but we sometimes need to go with the flow, and enjoy the domestication ride. As Darwin [1] commented “I was much struck by how entirely vague and arbitrary is the distinction between species and varieties”.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
This paper is the product of many often passionately argued, but always convivial, discussions during a ‘Domestication’ graduate seminar with students at the Department of Anthropology, University of Pennsylvania. The author would like to acknowledge the inspiring conversations and debates she had with Autumn Melby and Sam Yoder, and guest lecturers BrieAnna Langlie and Logan Kistler. She is also extremely grateful to Lisa Lodwick, Chantel White, Kathleen Morrison, and Glynis Bates for their comments on various iterations of this paper. Theodore Schurr for the encouragement and engaging conversation about the topic at the department social evening. The three reviewers who provided their advice are also gratefully acknowledged, as their comments have improved this paper and provided much for the author to think on further. This paper has been written whilst the author is a post-doctoral researcher at the University of Pennsylvania, and she is grateful to the support and opportunities she has received whilst there. Figures created with BioRender.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Darwin, C. On the Origin of Species; John Murray: London, UK, 1859. [Google Scholar]
- Darwin, C. The Variation of Animals and Plants under Domestication; John Murray: London, UK, 1868. [Google Scholar]
- Childe, V.G. Man Makes Himself; Watts and Company: London, UK, 1936. [Google Scholar]
- Vavilov, N.I. The Origin, Variation, Immunity and Breeding of Cultivated Plants; Chronica Botanica: Waltham, MA, USA, 1951; Volume 13, pp. 1–366. [Google Scholar]
- Fuller, D.Q.; Hildebrand, E. Domesticating Plants in Africa. In The Oxford Handbook of African Archaeology; Mitchell, P., Land, P., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 507–525. [Google Scholar]
- Langlie, B.S.; Mueller, N.G.; Spengler, R.N.; Fritz, G.J. Agricultural Origins from the Ground up: Archaeological Approaches to Plant Domestication. Am. J. Bot. 2014, 101, 1601–1617. [Google Scholar] [CrossRef]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Climer Vigueira, C.; Denham, T.; Dobney, K.; et al. Current Perspectives and the Future of Domestication Studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.-A.; Crawford, G.W.; Liu, L.; Sasaki, Y.; Chen, X. Archaeological Soybean (Glycine Max) in East Asia: Does Size Matter? PLoS ONE 2011, 6, e26720. [Google Scholar] [CrossRef] [PubMed]
- Zeder, M.A.; Emshwiller, E.; Smith, B.D.; Bradley, D.G. Documenting Domestication: The Intersection of Genetics and Archaeology. Trends Genet. 2006, 22, 139–155. [Google Scholar] [CrossRef]
- Gepts, P.; Famula, T.R.; Bettinger, R.L.; Brush, S.B.; Damania, A.B.; McGuire, P.E.; Qualset, C.O. Biodiversity in Agriculture: Domestication, Evolution and Sustainability; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012; ISBN 978-0-19-954906-1. [Google Scholar]
- Ingold, T. The Perception of the Environment: Essays on Livelihood, Dwelling, and Skill; Routledge: New York, NY, USA, 2000. [Google Scholar]
- Hillman, G.C.; Davies, M.S. Measured Domestication Rates in Wild Wheats and Barley under Primitive Cultivation, and Their Archaeological Implications. J. World Prehist. 1990, 4, 157–222. [Google Scholar] [CrossRef]
- Willcox, G.; Stordeur, D. Large-Scale Cereal Processing before Domestication during the Tenth Millennium Cal BC in Northern Syria. Antiquity 2012, 86, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Ma, Z.; Li, J.; Yu, J.; Stevens, C.; Zhuang, Y. Comparing Subsistence Strategies in Different Landscapes of North China 10,000 Years Ago. Holocene 2015, 25, 1957–1964. [Google Scholar] [CrossRef]
- Fuller, D.Q. Contrasting Patterns in Crop Domestication and Domestication Rates: Recent Archaeobotanical Insights from the Old World. Ann. Bot. 2007, 100, 903–924. [Google Scholar] [CrossRef]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and Processes in Crop Domestication: An Historical Review and Quantitative Analysis of 203 Global Food Crops: Tansley Review. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Fuller, D.Q.; Stevens, C.J. Between Domestication and Civilization: The Role of Agriculture and Arboriculture in the Emergence of the First Urban Societies. Veget. Hist. Archaeobot. 2019, 28, 263–282. [Google Scholar] [CrossRef] [Green Version]
- Spengler, R.N.; Petraglia, M.; Roberts, P.; Ashastina, K.; Kistler, L.; Mueller, N.G.; Boivin, N. Exaptation Traits for Megafaunal Mutualisms as a Factor in Plant Domestication. Front. Plant Sci. 2021, 12, 434. [Google Scholar] [CrossRef]
- Harlan, J.R. Crops and Man, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1992. [Google Scholar]
- Hammer, K. Das Domestikationssyndrom. Kulturpflanze 1984, 32, 11–34. [Google Scholar] [CrossRef]
- Abbo, S.; Saranga, Y.; Peleg, Z. Reconsidering Domestication of Legumes Versus Cereals In The Ancient Near East. Q. Rev. Biol. 2009, 84, 29–50. [Google Scholar] [CrossRef] [Green Version]
- Svizzero, S.; Ray, A.; Chakraborty, D. Awn Reduction and the Domestication of Asian Rice: A Syndrome or Crop Improvement Trait? Econ. Bot. 2019, 73, 477–488. [Google Scholar] [CrossRef]
- Sánchez-Pérez, R.; Pavan, S.; Mazzeo, R.; Moldovan, C.; Aiese Cigliano, R.; Del Cueto, J.; Ricciardi, F.; Lotti, C.; Ricciardi, L.; Dicenta, F.; et al. Mutation of a BHLH Transcription Factor Allowed Almond Domestication. Science 2019, 364, 1095–1098. [Google Scholar] [CrossRef] [PubMed]
- Caramelli, D. The Origins of Domesticated Cattle. Hum. Evol. 2006, 21, 107–122. [Google Scholar] [CrossRef]
- Künzl, C.; Sachser, N. The Behavioral Endocrinology of Domestication: A Comparison between the Domestic Guinea Pig (Cavia Apereaf.Porcellus) and Its Wild Ancestor, the Cavy (Cavia Aperea). Horm. Behav. 1999, 35, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Evin, A.; Owen, J.; Larson, G.; Debiais-Thibaud, M.; Cucchi, T.; Vidarsdottir, U.S.; Dobney, K. A Test for Paedomorphism in Domestic Pig Cranial Morphology. Biol. Lett. 2017, 13, 20170321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purugganan, M.D.; Fuller, D.Q. The Nature of Selection during Plant Domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef]
- Villmoare, B. Early Homo and the Role of the Genus in Paleoanthropology. Am. J. Phys. Anthr. 2018, 165, 72–89. [Google Scholar] [CrossRef] [Green Version]
- McNulty, K.P. Hominin Taxonomy and Phylogeny: What’s In a Name? Nat. Educ. Knowl. 2016, 7, 2. [Google Scholar]
- Haile-Selassie, Y.; Melillo, S.M.; Su, D.F. The Pliocene Hominin Diversity Conundrum: Do More Fossils Mean Less Clarity? Proc. Natl. Acad. Sci. USA 2016, 113, 6364–6371. [Google Scholar] [CrossRef] [Green Version]
- Lucas, G. Variations on a Theme: Assemblage Archaeology. CAJ 2017, 27, 187–190. [Google Scholar] [CrossRef]
- Rouse, I. The Classification of Artifacts in Archaeology. Am. Antiq. 1960, 25, 313–323. [Google Scholar] [CrossRef]
- Lefrancq, C.; Hawkes, J.D. Accepting Variation and Embracing Uncertainty: Creating a Regional Pottery Typology in South Asian Archaeology. Open Archaeol. 2020, 6, 269–328. [Google Scholar] [CrossRef]
- Darwin, C.; Wallace, A. On the Tendency of Species to Form Varieties; and on the Perpetuation of Varieties and Species by Natural Means of Selection. J. Proc. Linn. Soc. Lond. Zool. 1858, 3, 45–62. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.A. Darwin and the Inefficacy of Artificial Selection. Stud. Hist. Philos. Sci. Part A 1997, 28, 75–97. [Google Scholar] [CrossRef]
- Gregory, T.R. Artificial Selection and Domestication: Modern Lessons from Darwin’s Enduring Analogy. Evo. Edu. Outreach 2009, 2, 5–27. [Google Scholar] [CrossRef] [Green Version]
- Mendel, G. Versuche Über Plflanzenhybriden (Experiments in Plant Hybridization). Verhandlungen des naturforschenden Vereines in Brünn, Bd. IV für das Jahr 1865; Abhandlugen. 1865. Available online: http://www.netspace.org./MendelWeb/ (accessed on 20 December 2020).
- Mayr, E. Systematics and the Origin of Species; Columbia University Press: New York, NY, USA, 1942. [Google Scholar]
- Mayr, E. Species concepts and definitions. In The Species Problem; Mayr, E., Ed.; AAAS: Washington, DC, USA, 1957; pp. 1–22. [Google Scholar]
- Simpson, G.G. The Species Concept. Evolution 1951, 5, 285–298. [Google Scholar] [CrossRef]
- Templeton, A.R. The meaning of species and speciation: A genetic perspective. In Speciation and its Consequences; Otte, D., Endler, J.A., Eds.; Sinauer Associates: Sunderland, MA, USA, 1989; pp. 3–27. [Google Scholar]
- Mayr, E. Systematics and the Origin of Species, from the Viewpoint of a Zoologist; 1st Harvard University Press pbk. Ed.; Harvard University Press: Cambridge, MA, USA, 1999; ISBN 978-0-674-86250-0. [Google Scholar]
- Wilkins, J. Species Concepts in Modern Literature. Rep. NCSE 2006, 26, 36–45. [Google Scholar]
- Mayden, R.L. A Hierarchy of Species Concepts: The denouement in the saga of the species problem. In Species: The Units of Biodiversity; Claridge, M.F., Ed.; Chapman and Hall: London, UK, 1997; pp. 381–424. [Google Scholar]
- de Queiroz, K. The general Lineage Concept of Species and the defining Properties of the Species Category. In Species; Wilson, R.A., Ed.; MIT Press: Boston, MA, USA, 1999; pp. 49–89. [Google Scholar]
- De Queiroz, K. Species Concepts and Species Delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hey, J. The Mind of the Species Problem. Trends Ecol. Evol. 2001, 16, 326–330. [Google Scholar] [CrossRef]
- Broich, S.L.; Palmer, R.G. Evolutionary Studies of the Soybean: The Frequency and Distribution of Alleles among Collections of Glycine Max and G. Soja of Various Origin. Euphytica 1981, 30, 55–64. [Google Scholar] [CrossRef]
- Harlan, J.R.; Wet, J.M.J. Toward a Rational Classification of Cultivated Plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Harlan, J.R.; Zohary, D. Distribution of Wild Wheats and Barley. Science 1966, 153, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Jacomet, S. Identification of Cereal Remains from Archaeological Sites, 2nd ed.; IPAS: Basel, Switzerland, 2006. [Google Scholar]
- van Slageren, M.W. Wild Wheats: A Monograph of Aegilops L. and Amblyopyrum (Jaub. and Spach) Eig (Poaceae); Wageningen Agricultural University Papers; Wageningen Agricultural University: Wageningen, The Netherlands, 1994. [Google Scholar]
- Langlie, B.S. Morphological Analysis of Late Pre-Hispanic Peruvian Chenopodium Spp. Veget. Hist. Archaeobot. 2019, 28, 51–63. [Google Scholar] [CrossRef]
- Choi, J.Y.; Platts, A.E.; Fuller, D.Q.; Hsing, Y.-I.; Wing, R.A.; Purugganan, M.D. The Rice Paradox: Multiple Origins but Single Domestication in Asian Rice. Mol. Biol. Evol. 2017, 34, 969–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, D.Q.; Sato, Y.-I.; Castillo, C.; Qin, L.; Weisskopf, A.; Kingwell-Banham, E.; Song, J.; Ahn, S.-M.; van Etten, J. Consilience of Genetics and Archaeobotany in the Entangled History of Rice. Archaeol. Anthropol. Sci. 2010, 2, 115–131. [Google Scholar] [CrossRef]
- Gutaker, R.M.; Groen, S.C.; Bellis, E.S.; Choi, J.Y.; Pires, I.S.; Bocinsky, R.K.; Slayton, E.R.; Wilkins, O.; Castillo, C.C.; Negrão, S.; et al. Genomic History and Ecology of the Geographic Spread of Rice. Nature Plants 2020, 6, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.; Weisskopf, A.; Castillo, C.; Murphy, C.; Kingwell-Banham, E.; Qin, L.; Fuller, D.Q. A Tale of Two Rice Varieties: Modelling the Prehistoric Dispersals of Japonica and Proto- Indica Rices. Holocene 2018, 28, 1745–1758. [Google Scholar] [CrossRef] [Green Version]
- Fuller, D.Q. Pathways to Asian Civilizations: Tracing the Origins and Spread of Rice and Rice Cultures. Rice 2011, 4, 78–92. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Sato, Y.; Nakamura, I. Evolutionary Analysis of Two Plastid DNA Sequences in Cultivated and Wild Species of Oryza. Breed. Sci. 2008, 58, 225–233. [Google Scholar] [CrossRef] [Green Version]
- de Langhe, E.; Vrydaghs, L.; de Maret, P.; Perrier, X.; Denham, T. Why Bananas Matter: An Introduction to the History of Banana Domestication. Ethnobot. Res. Appl. 2009, 7, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Denham, T.; Barton, H.; Castillo, C.; Crowther, A.; Dotte-Sarout, E.; Florin, S.A.; Pritchard, J.; Barron, A.; Zhang, Y.; Fuller, D.Q. The Domestication Syndrome in Vegetatively Propagated Field Crops. Ann. Bot. 2020, 125, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Bates, J.; Petrie, C.A.; Singh, R.N. Approaching Rice Domestication in South Asia: New Evidence from Indus Settlements in Northern India. J. Archaeol. Sci. 2017, 78, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.H. Various Cultigens, and Transfers in Nomenclature. Gentes Herbarum 1923, 1, 113–136. [Google Scholar]
- Spencer, R.D.; Cross, R.G. The International Code of Botanical Nomenclature (ICBN), the International Code of Nomenclature for Cultivated Plants (ICNCP), and the Cultigen. Taxon 2007, 56, 938–940. [Google Scholar] [CrossRef]
- Spengler, R.N. Anthropogenic Seed Dispersal: Rethinking the Origins of Plant Domestication. Trends Plant Sci. 2020, 25, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Fritz, G.J.; Bruno, M.C.; Langlie, B.S.; Smith, B.D.; Kistler, L. Cultigen Chenopods in the Americas: A Hemispherical Perspective. In Social Perspectives on Ancient Lives from Paleoethnobotanical Data; Sayre, M.P., Bruno, M.C., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 55–75. ISBN 978-3-319-52847-2. [Google Scholar]
- Mueller, N.G.; White, A.; Szilagyi, P. Experimental Cultivation of Eastern North America’s Lost Crops: Insights into Agricultural Practice and Yield Potential. J. Ethnobiol. 2019, 39, 549. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.E.; Medin, D.L. Categories and Concepts; Harvard University Press: Cambridge, MA, USA, 1981. [Google Scholar]
Figure 1.
Schematic comparison of taxonomic and evolutionary approaches to categorizing taxa using a hypothetical cereal. Under the taxonomic scheme the different ‘features’, in this case morphological, are used to lump or split the taxa into categories. This can be as many or as few as the accessioner deems appropriate. The multiple boxes drawn show how this could be achieved—it could be highly split into four groups, or into two based on whether the awn presence or rachis toughness, or into a single group based on this being a taxa distinct from others that look different from it over all. On the other hand, the evolutionary approach, using the same features, again morphological, is looking to see relatedness between the taxa, and suggests that the loss of defensive mechanism is an indicator of relatedness, and the plant with no loss of defenses should be placed in a group on its own. The motivations for ‘creating’ species/categories can therefore have profound effects on how we divide and identify the objects of our study, and create models for how they come into being. [Created with BioRender.com, under a CC BY license, with permission from Biorender, original copyright 2020].
Figure 1.
Schematic comparison of taxonomic and evolutionary approaches to categorizing taxa using a hypothetical cereal. Under the taxonomic scheme the different ‘features’, in this case morphological, are used to lump or split the taxa into categories. This can be as many or as few as the accessioner deems appropriate. The multiple boxes drawn show how this could be achieved—it could be highly split into four groups, or into two based on whether the awn presence or rachis toughness, or into a single group based on this being a taxa distinct from others that look different from it over all. On the other hand, the evolutionary approach, using the same features, again morphological, is looking to see relatedness between the taxa, and suggests that the loss of defensive mechanism is an indicator of relatedness, and the plant with no loss of defenses should be placed in a group on its own. The motivations for ‘creating’ species/categories can therefore have profound effects on how we divide and identify the objects of our study, and create models for how they come into being. [Created with BioRender.com, under a CC BY license, with permission from Biorender, original copyright 2020].

Figure 2.
The complex pathways to rice domestication, schematized and oversimplified to place them in the archaeological context. At the start of the process, we have sites like Lahuradewa with evidence for human use of likely wild rice, and archaeogenetic evidence that suggests wild exploitation of nivara, at the other end we have sites like Mahagara where the spikelet bases show domesticated rice use, and the (archaeo)genetics suggests the hybridization was if not complete then certainly under way. At what point in between these does nivara become another species? Does it indeed become another species? The dead ends seen (archaeo)genetically and in the increasing evidence for domesticated spikelet bases found alongside wild types suggests some changes were occurring before hybridization—should these be given the almost species-like name ‘proto-indica’? The determinism behind this epithet—that these will become, are becoming, indica is problematic due to the hindsight placed upon them by us, and the dead ends we see in the genetic data. Or instead is proto-indica perhaps another name for the human processes behind the changes occurring, for pre-domestication cultivation? Genetics taken from Fuller et al. ([56]: Figure 2), and the original archaeobotanical data for Masudpur I, Masudpur VII, and Bahola can be found in [63]. [Created with BioRender.com, under a CC BY license, with permission from Biorender, original copyright 2020].
Figure 2.
The complex pathways to rice domestication, schematized and oversimplified to place them in the archaeological context. At the start of the process, we have sites like Lahuradewa with evidence for human use of likely wild rice, and archaeogenetic evidence that suggests wild exploitation of nivara, at the other end we have sites like Mahagara where the spikelet bases show domesticated rice use, and the (archaeo)genetics suggests the hybridization was if not complete then certainly under way. At what point in between these does nivara become another species? Does it indeed become another species? The dead ends seen (archaeo)genetically and in the increasing evidence for domesticated spikelet bases found alongside wild types suggests some changes were occurring before hybridization—should these be given the almost species-like name ‘proto-indica’? The determinism behind this epithet—that these will become, are becoming, indica is problematic due to the hindsight placed upon them by us, and the dead ends we see in the genetic data. Or instead is proto-indica perhaps another name for the human processes behind the changes occurring, for pre-domestication cultivation? Genetics taken from Fuller et al. ([56]: Figure 2), and the original archaeobotanical data for Masudpur I, Masudpur VII, and Bahola can be found in [63]. [Created with BioRender.com, under a CC BY license, with permission from Biorender, original copyright 2020].

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bates, J. Is Domestication Speciation? The Implications of a Messy Domestication Model in the Holocene. Agronomy 2021, 11, 784. https://doi.org/10.3390/agronomy11040784
AMA Style
Bates J. Is Domestication Speciation? The Implications of a Messy Domestication Model in the Holocene. Agronomy. 2021; 11(4):784. https://doi.org/10.3390/agronomy11040784
Chicago/Turabian StyleBates, Jennifer. 2021. "Is Domestication Speciation? The Implications of a Messy Domestication Model in the Holocene" Agronomy 11, no. 4: 784. https://doi.org/10.3390/agronomy11040784
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.