
Combined Use of Two Trichoderma Strains to Promote Growth of Pakchoi (Brassica chinensis L.)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Samples and Fungal Strain Culture
2.2. In Vitro Promoting Traits of Strains
2.3. Bioassay for Germination Test
2.4. Determination of Germination Parameters
2.5. Test for the Capacity to Promote Plant Growth (Pot Experiment)
2.6. Biomass and Quality Measurement
2.7. Statistical Analysis
3. Results
3.1. In Vitro-Promoting Traits of Strains
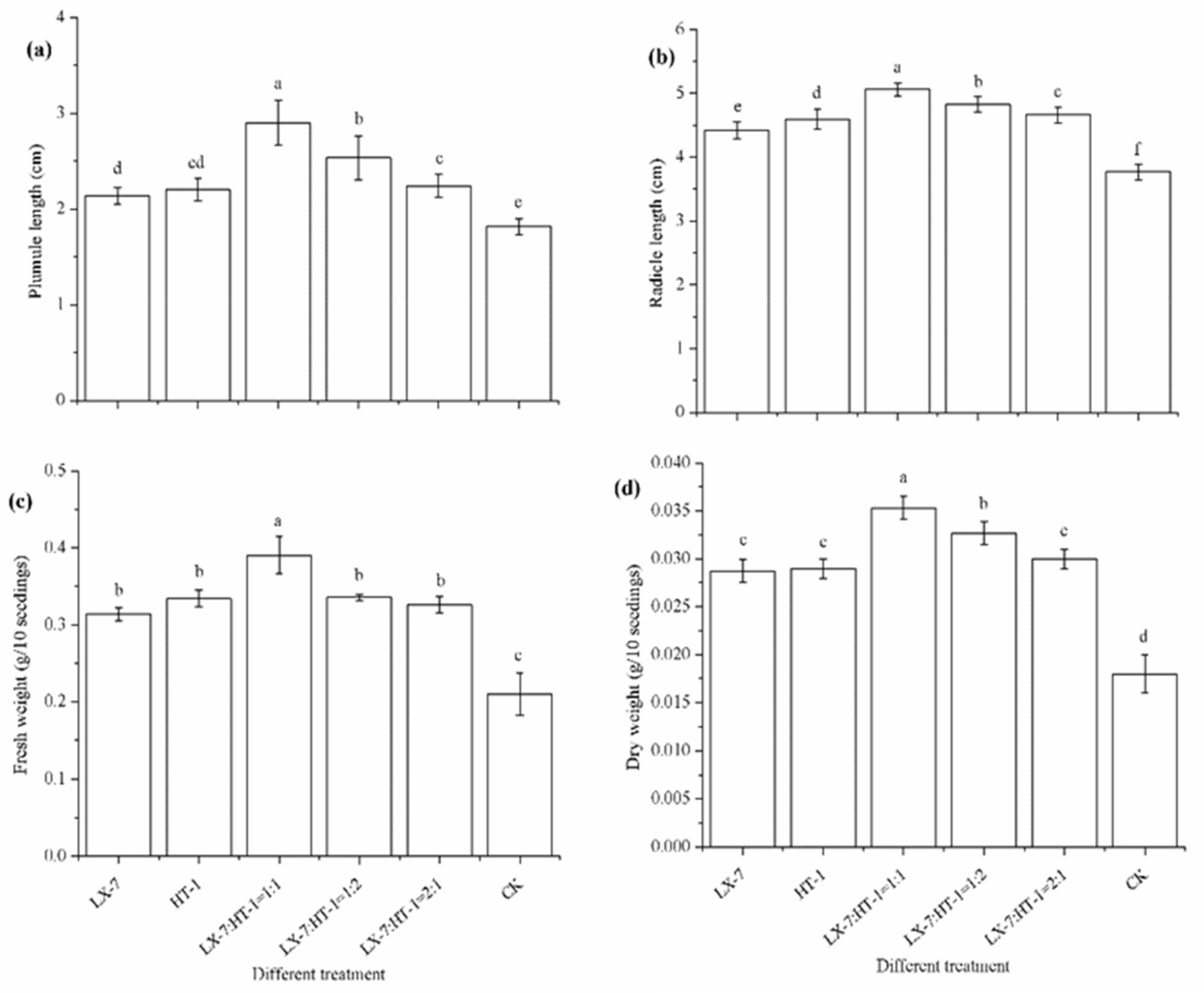
3.2. Effects of HT-1 and LX-7 Strains on Seeds Germination and Seedlings Growth of Pakchoi
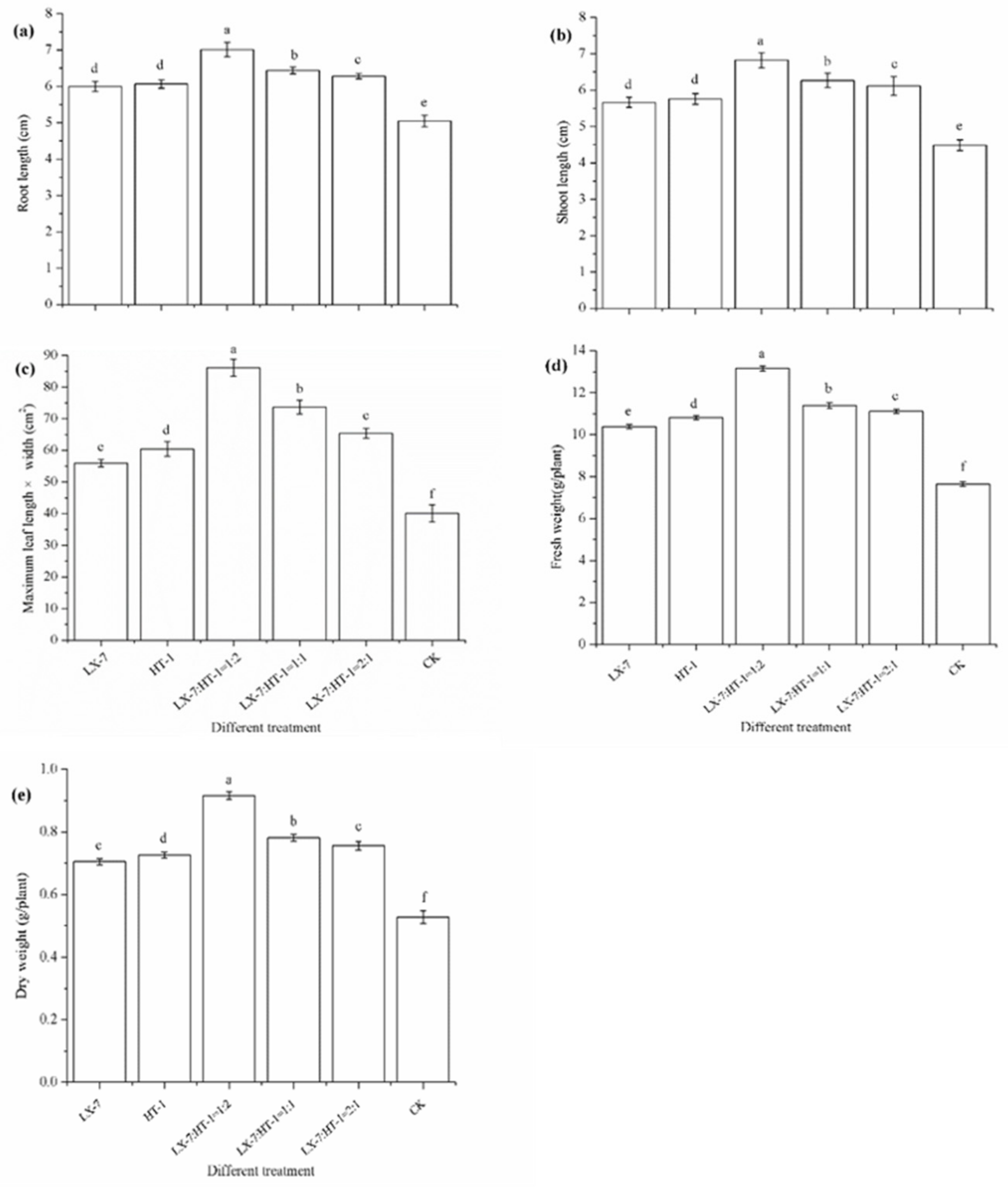
3.3. Effect of HT-1 and LX-7 Strains on Pakchoi Growth in Greenhouse
3.4. Effect of HT-1 and LX-7 Strains on Pakchoi Quality in Greenhouse
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Compant, S.; van der Heijden, M.G.; Sessitsch, A. Climate change effects on beneficial plant–microorganism interactions. FEMS Microbiol. Ecol. 2010, 73, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.F.; Chaparro, J.M.; Reardon, K.F.; Zhang, R.; Shen, Q.; Vivanco, J.M. Rhizosphere interactions: Root exudates, microbes, and microbial communities. Botany 2014, 92, 267–275. [Google Scholar] [CrossRef]
- Balestrini, R.; Bonfante, P. The interface compartment in arbuscularmycorrhizae: A special type of plant cell wall? Plant Biosyst. 2005, 139, 8–15. [Google Scholar] [CrossRef]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Halifu, S.; Deng, X.; Song, X.H.; Song, R. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef] [Green Version]
- Lorito, M.; Harman, G.E.; Mastouri, F. Translational research on Trichoderma: From omics to the field. Annu. Rev. Phytopathol. 2010, 48, 395–417. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Chang, Y.C.; Baker, R.; Kleifeld, O.; Chet, I. Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis. 1986, 70, 145–148. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Aoki, Y.; Haga, S.; Suzuki, S. Direct antagonistic activity of chitinase produced by Trichoderma sp. SANA20 as biological control agent for grey mould caused by Botrytis cinerea. Cogent Biol. 2020, 6, 1747903. [Google Scholar] [CrossRef]
- Al-Askar, A.A.; Saber, W.; Ghoneem, K.M.; Hafez, E.E.; Ibrahim, A.A. Crude Citric Acid of Trichoderma asperellum: Tomato Growth Promotor and Suppressor of Fusarium oxysporum f. sp. lycopersici. Plants 2021, 10, 222. [Google Scholar] [CrossRef]
- Ferreira, F.V.; Herrmann-Andrade, A.M.; Calabrese, C.D.; Bello, F.; Vázquez, D.; Musumeci, M.A. Effectiveness of Trichoderma strains isolated from the rhizosphere of citrus tree to control Alternaria alternata, Colletotrichum gloeosporioides and Penicillium digitatum A21 resistant to pyrimethanil in post-harvest oranges (Citrus sinensis L. (Osbeck)). J. Appl. Microbiol. 2020, 129, 712–727. [Google Scholar] [CrossRef]
- Geffriaud, V.E.; Vicente, R.; Díaz, O.V.; Reinaldo, J.J.N.; Trillas, M.I. Application of Trichoderma asperellum T34 on maize (Zea mays) seeds protects against drought stress. Planta 2020, 252, 8. [Google Scholar] [CrossRef]
- Manna, M.C.; Sahu, A.; De, N.; Thakur, J.K.; Mandal, A.; Bhattacharjya, S.; Ghosh, A.; Rahman, M.M.; Naidu, R.; Singh, U.B.; et al. Novel bio-filtration method for the removal of heavy metals from municipal solid waste. Environ. Technol. Innov. 2020, 17, 100619. [Google Scholar] [CrossRef]
- Velmourougane, K.; Prasanna, R.; Chawla, G.; Nain, L.; Kumar, A.; Saxena, A.K. Trichoderma–Azotobacter biofilm inoculation improves soil nutrient availability and plant growth in wheat and cotton. J. Basic Microbiol. 2019, 59, 632–644. [Google Scholar] [CrossRef]
- Eslahi, N.; Kowsari, M.; Moghaddam, A.; Zamani, M. Influence of recombinant Trichoderma strains on growth of bean (Phaseolus vulgaris L.) by increased root colonization and induction of root growth related genes. Sci. Hortic. 2020, 261, 108932. [Google Scholar] [CrossRef]
- Camacho, M.; Santamaría, C.; Temprano, F.; Rodriguez-Navarro, D.N.; Daza, A. Co-inoculation with Bacillus sp. CECT 450 improves nodulation in Phaseolus vulgaris L. Can. J. Microbiol. 2001, 47, 1058–1062. [Google Scholar] [CrossRef]
- Eşitken, A.; Yıldız, H.; Ercisli, S.; Donmez, M.F.; Turan, M.; Gunes, A. Effects of plant growth promoting bacteria (PGPB) on yield, growth and nutrient contents of organically grown strawberry. Sci. Hortic Amst. 2010, 124, 62–66. [Google Scholar] [CrossRef]
- Banchio, E.; Bogino, P.; Zygadlo, J.A.; Giordano, W. Plant growth promoting rhizobacteria improve growth and essential oil yield in Origanum majorana L. Biochem. Syst. Ecol. 2008, 36, 766–771. [Google Scholar] [CrossRef]
- Karlidag, H.; Esitken, A.; Turan, M.; Sahin, F. Effects of root inoculation of plant growth promoting rhizobacteria(PGPR)on yield, growth and nutrient element contents of leaves of apple. Sci. Hortic. 2007, 114, 16–20. [Google Scholar] [CrossRef]
- Emine, O.; Ahmet, E.; Sezai, E.; Metin, T.; Fikrettin, S. Effects of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient contents in organically growing raspberry. Sci. Hortic. 2006, 110, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, W.; He, J.; Zhang, L.; Wei, Y.; Yang, M. Nitric oxide alleviates salt stress in seed germination and early seedling growth of pakchoi (Brassica chinensis L.) by enhancing physiological and biochemical parameters. Ecotoxicol. Environ. Saf. 2020, 87, 109785. [Google Scholar] [CrossRef]
- Hanson, P.; Yang, R.; Chang, L.; Ledesma, L.; Ledesma, D. Contents of carotenoids, ascorbic acid, minerals and total glucosinolates in leafy brassica pakchoi (Brassica rapa, L. chinensis) as affected by season and variety. J. Sci. Food Agric. 2009, 89, 906–914. [Google Scholar] [CrossRef]
- Zheng, S.; Jiang, J.; He, M.; Zou, S.; Wang, C. Effect of Kelp Waste Extracts on the Growth and Development of Pakchoi (Brassica chinensis L.). Sci. Rep. 2016, 6, 38683. [Google Scholar] [CrossRef] [Green Version]
- Wani, Z.A.; Mirza, D.N.; Arora, P.; Riyaz-Ul-Hassan, S. Molecular phylogeny, diversity, community structure, and plant growth promoting properties of fungal endophytes associated with the corms of saffron plant: An insight into the microbiome of Crocus sativus Linn. Fungal Biol. 2016, 120, 1509–1524. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Yuan, Z.S.; Liu, F.; Zhang, G.F. Characteristics and biodiversity of endophytic phosphorus- and potassium-solubilizing bacteria in Moso Bamboo (Phyllostachys edulis). Acta Biol. Hung. 2015, 66, 449. [Google Scholar] [CrossRef] [Green Version]
- Gratani, L. A non-destructive method to determine chlorophyll content of leaves. Photosynthetica 1992, 26, 469–473. [Google Scholar] [CrossRef]
- Song, J.; Huang, H.; Hao, Y.; Song, S.; Zhang, Y.; Su, W.; Liu, H. Nutritional quality, mineral and antioxidant content in lettuce affected by interaction of light intensity and nutrient solution concentration. Sci. Rep. 2020, 10, 2796. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Song, S.W.; Liao, G.X.; Liu, H.C.; Sun, G.W.; Chen, R.Y. Effect of ammonium and nitrate ratio on nutritional quality of Chinese kale. Adv. Mater. Res. 2012, 461, 13–16. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid colorimetric determination of nitrate in plant tissue by nitration of salicylic acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Nomkong, R.F.; Ejoh, R.A.; Dibanda, R.F.; Gabriel, M.N. Bioavailability of Iron and Related Components in Cooked Green Leafy Vegetables Consumed in Cameroon. Food Nutr. Sci. 2019, 10, 1096–1111. [Google Scholar] [CrossRef] [Green Version]
- Kotasthane, A.; Agrawal, T.; Kushwah, R.; Rahatkar, O. In-vitro antagonism of Trichoderma spp. Against Sclerotium rolfsii and Rhizoctonia solani and their response towards growth of cucumber, bottle gourd and bitter gourd. Eur. J. Plant Pathol. 2015, 141, 523–543. [Google Scholar] [CrossRef]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Graham, N.; Inzé, D.; Sandberg, G.; Casero, P.J.; et al. Auxin Transport Promotes Arabidopsis Lateral Root Initiation. Plant Cell 2001, 13, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Mweetwa, A.M.; Chilombo, G.; Gondwe, B.M. Nodulation, nutrient uptake and yield of common bean inoculated with Rhizobia and Trichoderma in an Acid Soil. J. Agric. Sci. 2009, 8, 61. [Google Scholar] [CrossRef]
- Wang, C.; Zhuang, W.Y. Carbon metabolic profiling of Trichoderma strains provides insight into potential ecological niches. Mycologia 2020, 112, 213–223. [Google Scholar] [CrossRef]
- Babana, A.H.; Antoun, H. Biological system for improving the availability of Tilemsi phosphate rock for wheat (Triticum aestivum L.) cultivated in Mali. Nutr. Cycl. Agroecosyst. 2005, 72, 147–157. [Google Scholar] [CrossRef]
- Fravel, D.R.; Lewis, J.A. Effect of label and sublabel rates of metam sodium in combination with Trichoderma hamatum, T. harzianum, T. virens, T. viride on survival and growth of Rhizoctonia solani. Phytoparasitica 2004, 32, 111–118. [Google Scholar] [CrossRef]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multi level properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Chandanie, W.A.; Kubota, M.; Hyakumachi, M. Interaction between the arbuscular mycorrhizal fungus Glomus mosseae and plant growth-promoting fungi and their significance for enhancing plant growth and suppressing damping-off of cucumber (Cucumis sativus L.). Appl. Soil Ecol. 2009, 41, 336–341. [Google Scholar] [CrossRef]
- Martinez, A.; Obertello, M.; Pardo, A.; Ocampo, J.A.; Godeas, A. Interactions between Trichoderma pseudokoningii strains and the arbuscular mycorrhizal fungi Glomus mosseae and Gigaspora rosea. Mycorrhiza 2004, 14, 79–84. [Google Scholar] [CrossRef]
- De Jaeger, N.; Declerck, S.; de la Providencia, I.E. Mycoparasitism of arbuscular mycorrhizal fungi: A pathway for the entry of saprotrophic fungi into roots. FEMS Microbiol. Ecol. 2010, 73, 312–322. [Google Scholar] [CrossRef] [Green Version]
- De Jaeger, N.; de la Providencia, I.E.; Rouhier, H.; Declerck, S. Co-entrapment of Trichoderma harzianum and Glomus sp. within alginate beads: Impact on the arbuscular mycorrhizal fungi life cycle. J. Appl. Microbiol. 2011, 111, 125–135. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | IAA (mg/L) | Siderophore (%) | Organic Phosphate Solubilization (mg/L) | Inorganic Phosphate Solubilization (mg/L) | Potassium Absorbable (mg/L) |
|---|---|---|---|---|---|
| HT-1 | 10.06 ± 0.08 d | 39.41 ± 0.27 d | 0 | 0 | 0.00 ± 0.00 d |
| LX-7 | 9.29 ± 0.09 e | 31.22 ± 0.23 e | 0 | 0 | 2.05 ± 0.07 a |
| LX-7:HT-1 = 1:2 | 23.07 ± 0.05 a | 52.75 ± 0.15 a | 0 | 0 | 1.23 ± 0.06 c |
| LX-7:HT-1 = 1:1 | 20.63 ± 0.12 b | 47.96 ± 0.32 b | 0 | 0 | 1.46 ± 0.07 b |
| LX-7:HT-1 = 2:1 | 16.19 ± 0.05 c | 43.77 ± 0.19 c | 0 | 0 | 1.57 ± 0.07 b |
| Treatment | GP (%) | GI | GE (%) | VI |
|---|---|---|---|---|
| LX-7 | 91.00 ± 1.00 b | 127.91 ± 3.01 c | 62.67 ± 1.15 c | 565.45 ± 23.00 d |
| HT | 91.67 ± 1.15 b | 132.33 ± 2.51 b | 66.67 ± 2.08 b | 608.10 ± 10.80 c |
| LX-7:HT-1 = 1:1 | 94.67 ± 1.15 a | 139.36 ± 1.01 a | 71.33 ± 1.53 a | 705.43 ± 6.62 a |
| LX-7:HT-1 = 1:2 | 92.67 ± 1.53 ab | 135.51 ± 1.79 ab | 68.33 ± 153 b | 654.41 ± 7.76 b |
| LX-7:HT-1 = 2:1 | 92.33 ± 1.15 ab | 135.44 ± 2.19 ab | 68.00 ± 1.00 b | 631.43 ± 19.26 bc |
| CK | 88.67 ± 1.53 c | 112.54 ± 3.08 d | 57.33 ± 1.53 d | 424.14 ± 12.95 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Hou, Q.; Jia, L.; Sun, K. Combined Use of Two Trichoderma Strains to Promote Growth of Pakchoi (Brassica chinensis L.). Agronomy 2021, 11, 726. https://doi.org/10.3390/agronomy11040726
Chen D, Hou Q, Jia L, Sun K. Combined Use of Two Trichoderma Strains to Promote Growth of Pakchoi (Brassica chinensis L.). Agronomy. 2021; 11(4):726. https://doi.org/10.3390/agronomy11040726
Chicago/Turabian StyleChen, Dawei, Qinzheng Hou, Lingyun Jia, and Kun Sun. 2021. "Combined Use of Two Trichoderma Strains to Promote Growth of Pakchoi (Brassica chinensis L.)" Agronomy 11, no. 4: 726. https://doi.org/10.3390/agronomy11040726