
Defense Responses in the Interactions between Medicinal Plants from Lamiaceae Family and the Two-Spotted Spider Mite Tetranychus urticae Koch (Acari: Tetranychidae)

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. Measurement of Trichomes
2.3. Choice Test
2.4. Non-Choice Test: Mites and Plant Colonization
2.5. Biological Parameters of T. urticae
2.6. Physiological Analysis
2.6.1. Hydrogen Peroxide Concentration
2.6.2. Malondialdehyde Content (MDA)
2.6.3. Determination of the Antioxidant Enzymes Activity
Preparation of Extract
2.7. Statistical Analysis
3. Results
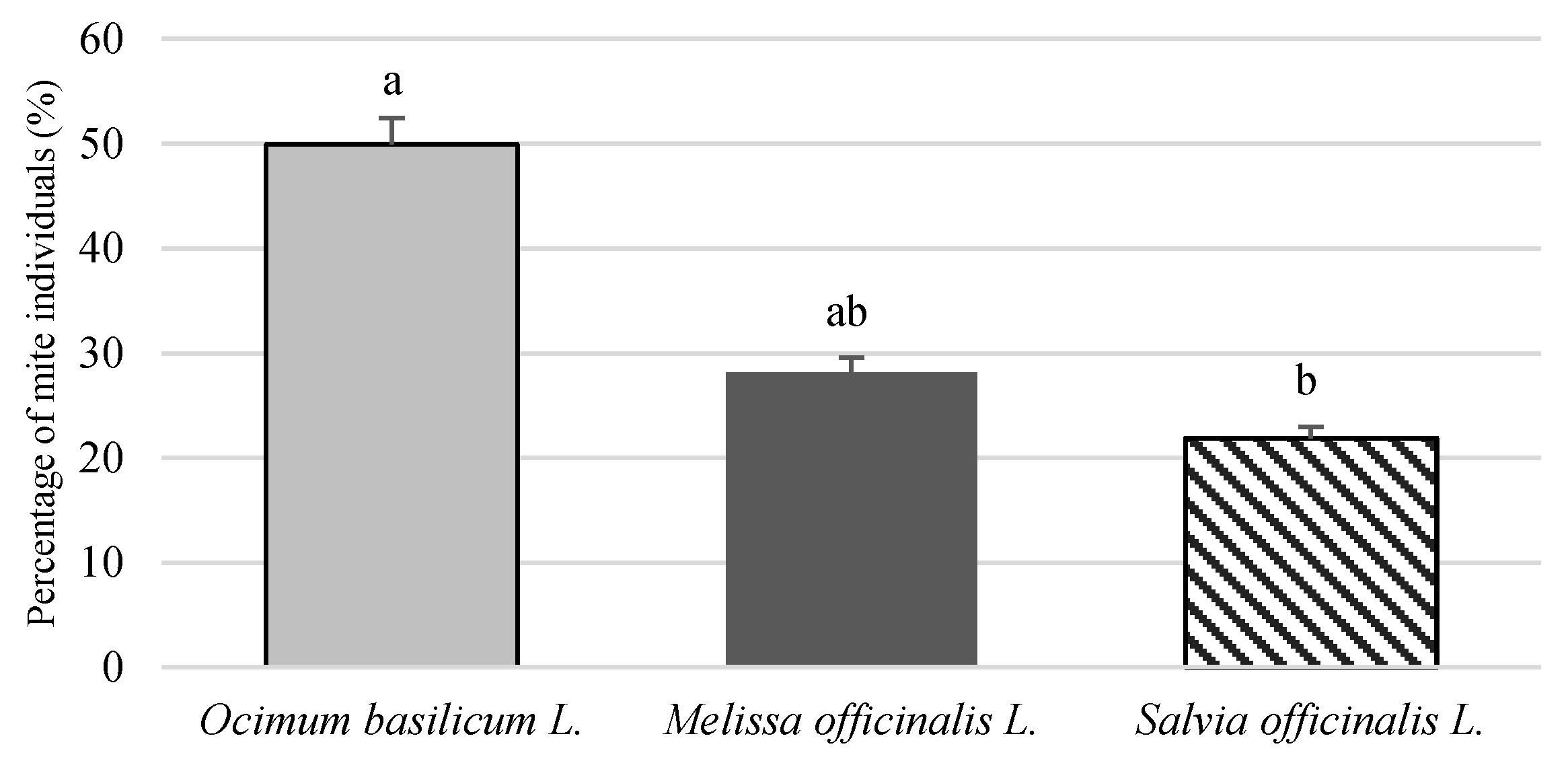
3.1. Plants Infestation by T. urticae
3.2. Changes in Hydrogen Peroxide Concentration
3.3. Changes in Malondialdehyde Concentration
3.4. Changes in Antioxidant Enzymes Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WHO guidelines on good agricultural and collection practices [GACP] for medicinal plants. World Health Organization, 2003. Available online: https://apps.who.int/iris/handle/10665/42783 (accessed on 2 February 2020).
- Hishe, M.; Asfaw, Z.; Giday, M. Review on value chain analysis of medicinal plants and the associated challenges. J. Med. Plants Stud. 2016, 4, 45–55. [Google Scholar]
- Lubiarz, M.; Goszczyński, W.; Cichocka, E. Invertebrates inhabiting culinary herbs grown under cover. J. Plant Prot. Res. 2008, 53, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Spychalski, G. Determinants of growing herbs in Polish agriculture. Herba Pol. 2013, 59, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Hikosaka, S.; Iwamoto, N.; Goto, E.; Ching-Hui, C. Effects of supplemental lighting on growth and medicinal compounds of Japanese honeysuckle (Lonicera japonica Thunb.). Environ. Control Biol. 2017, 55, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Majkowska-Gadomska, J.; Kulczycka, A.; Dobrowolski, A.; Mikulewicz, E. Yield and nutritional value of basil grown in a greenhouse. Acta Agrophys. 2017, 24, 455–464. [Google Scholar]
- Cazaux, M.; Navarro, M.; Bruinsma, K.A.; Zhurov, V.; Negrave, T.; Van Leeuwen, T.; Grbic, V.; Grbic, M. Application of Two-spotted Spider Mite Tetranychus urticae for plant-pest. Interaction Studies. JoVE 2014, 89, 51738. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Gupta, S.K. Some Records of Mites on Medicinal Plants from South Bengal with their Economic Importance. BFAIJ 2016, 8, 108–111. [Google Scholar]
- Vacante, V. The Handbook of Mites of Economic Plants: Identification, Bio-Ecology and Control; CABI International: Wallingford, UK, 2016; p. 890. [Google Scholar]
- Migeon, A.; Dorkeld, F. A Comprehensive Database for the Tetranychidae. 2006–2020. Institute for Agronomy Research, Center for Biology and Management of Populations: Montpellier, France. Available online: www.montpellier.inra.fr/CBGP/spmweb (accessed on 2 March 2020).
- Golan, K.; Kot, I.; Górska-Drabik, E.; Garrido Jurado, I.; Kmieć, K.; Łagowska, B. Physiological response of basil plants to two-spotted spider mite (Acari: Tetranychidae) infestation. J. Econ. Entomol. 2019, 112, 948–956. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.L.; Lee, J.H. Leaf cell and tissue damage of cucumber caused by two spotted spider mite (Acari: Tetranychidae). J. Econ. Entomol. 2002, 95, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Zhurov, V.; Navarro, M.; Bruinsma, K.A.; Arbona, V.; Santamaria, M.E.; Cazaux, M.; Wybouw, N.; Osborne, E.J.; Ens, C.; Rioja, C.; et al. Reciprocal responses in the interaction between Arabidopsis and the cell-content-feeding chelicerate herbivore spider mite. Plant Physiol. 2014, 164, 384–399. [Google Scholar] [CrossRef] [Green Version]
- Bensoussan, N.; Santamaria, M.E.; Zhurov, V.; Diaz, I.; Grbic, M.; Grbic, V. Plant-herbivore interaction: Dissection of the cellular pattern of Tetranychus urticae feeding on the host plant. Front. Plant Sci. 2016, 7, 1105. [Google Scholar] [CrossRef] [Green Version]
- Rioja, C.; Zhurov, V.; Bruinsma, K.; Grbic, M.; Grbic, V. Plant-Herbivore Interactions: A Case of an Extreme Generalist, the Two-Spotted Spider Mite Tetranychus urticae. Mol. Plant Microbe Interact. 2017, 30, 935–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, M.E.; Arnaiz, A.; Velasco-Arroyo, B.; Grbic, V.; Diaz, I.; Martinez, M. Arabidopsis response to the spider mite Tetranychus urticae depends on the regulation of reactive oxygen species homeostasis. Sci. Rep. 2018, 8, 9432. [Google Scholar] [CrossRef]
- Santamaria, M.E.; Arnaiz, A.; Rosa-Diaz, I.; González-Melendi, P.; Romero-Hernandez, G.; Ojeda-Martinez, D.A.; Garcia, A.; Contreras, E.; Martinez, M.; Diaz, I. Plant Defenses Against Tetranychus urticae: Mind the Gaps. Plants 2020, 9, 464. [Google Scholar] [CrossRef] [Green Version]
- Ohtsuka, K.; Osakabe, M. Deleterious effects of UV-B radiation on herbivorous spider mites: They can avoid it by remaining on lower leaf surfaces. Environ. Entomol. 2009, 38, 920–929. [Google Scholar] [CrossRef] [Green Version]
- Sivritepe, N.; Kumral, N.A.; Erturk, U.; Yerlikaya, C.; Kumral, A. Responses of Grapevines to Two-Spotted Spider Mite Mediated Biotic Stress. J. Biol. Sci. 2009, 9, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Gahukar, R.T. Pest and disease management in important medicinal plants in India: A review. NFS J. 2017, 220–224. [Google Scholar] [CrossRef]
- Agrawal, A.A. Host-range evolution: Adaptation and trade-off in fitness of mites on alternative hosts. Ecology 2000, 81, 500–508. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Rincon, R.A.; Rodriguez, D.; Coy-Barrera, E. Botanicals against Tetranychus urticae Koch under laboratory conditions: A survey of alternatives for controlling pest mites. Plants 2019, 8, 272. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, Z.T. Basics of Plant Resistance to Pests, 2nd ed.; PWRiL: Warsaw, Poland, 1988; p. 260. [Google Scholar]
- Mur, L.; Kenton, P.; Draper, J. In-plant measurements of oxidative bursts elicited by avirulent and virulent bacterial pathogens suggests that H2O2 is insufficient to elicit cell death in tobacco. Plant Cell Environ. 2005, 28, 548–561. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Tariq, A.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [Green Version]
- Caverzan, A.; Casassola, A.; Patussi Brammer, S. Antioxidant responses of wheat plants under stress. Genet. Mol. Biol. 2016, 39, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Finkel, T. Free radicals and senescence. Exp. Cell Res. 2008, 314, 1918–1922. [Google Scholar] [CrossRef]
- Nasir Kham, M.; Manzer, H.S.; Mohammad, F.; Naeem, M.; Masroor, M.; Kham, A. Calcium chloride and gibberellic acid protect linseed from NaCl stress by inducing antioxidative defence system and osmoprotectant accumulation. Acta Physiol. Plant 2010, 32, 121–132. [Google Scholar] [CrossRef]
- Gulsen, O.; Eickhoff, T.; Heng-Moss, R.; Shearman, F.; Baxendale, G.; Sarath Lee, D. Characterization of peroxidase changes in resistant and susceptible warm-season turfgrasses challenged by Blissus occiduus. Arthropod-Plant Interact. 2010, 4, 45–55. [Google Scholar] [CrossRef]
- Mai, V.C.; Bednarski, W.; Borowiak-Sobkowiak, B.; Wilkaniec, B.; Samardakiewicz, S.; Morkunas, I. Oxidative stress in pea seedling leaves in response to Acyrtosiphon pisum infestation. Phytochemistry 2013, 93, 49–62. [Google Scholar] [CrossRef]
- Kmieć, K.; Kot, I.; Rubinowska, K.; Łagowska, B.; Golan, K.; Górska-Drabik, E. Physiological reaction of Phalaenopsis × hybridum ‘Innocence’ on Pseudococcus longispinus (Targoni Tozetti) feeding. Acta Sci. Pol-Hortoru 2014, 13, 85–95. [Google Scholar]
- Kmieć, K.; Kot, I.; Golan, K.; Górska-Drabik, E.; Łagowska, B.; Rubinowska, K.; Michałek, W. Physilogical response of orchids to melaybugs (Hemiptera: Pseudococcidae) infestation. J. Econ. Entomol. 2016, 109, 2489–2494. [Google Scholar] [CrossRef] [PubMed]
- Kot, I.; Kmieć, K.; Górska-Drabik, E.; Golan, K.; Rubinowska, K.; Łagowska, B. The effect of mealybug Pseudococcus longispinus (Targioni Tozzetti) infestation of different density on physiological responses of Phalaenopsis × hybridum ‘Innocence’. Bull. Entomol. Res. 2015, 105, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Conklin, P.L. Recent advances in the role and biosynthesis of ascorbic acid in plants. Plant Cell Environ. 2001, 24, 383–394. [Google Scholar] [CrossRef]
- Świątek, M.; Kiełkiewicz, M.; Zagdańska, B. Insect-resistant Bt-maize response to the short-term non-target mite-pest infestation and soil drought. Acta Physiol. Plant 2014, 36, 2705–2715. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.X.; Zhang, S.X.; Liang, Z.S. Nitrogen rates and water stress effect on production, lipid peroxidation and antioxidative enzyme activities in two maize (Zea mays L.) genotypes. J. Agron. Crop Sci. 2007, 193, 387–397. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signaling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, K.; Kaur, N. Differential responses of antioxidative defence system to long-term field drought in wheat (Triticum aestivum L.) genotypes differing in drought tolerance. J. Agron. Crop Sci. 2012, 198, 185–195. [Google Scholar] [CrossRef]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A large family of Class III Plant Peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Maffei, M.E.; Mithöfer, A.; Boland, W. Before gene expression: Early events in plant–insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, A.; Smeets, K.; Ruytinx, J. The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings. J. Plant Physiol. 2011, 168, 309–316. [Google Scholar] [CrossRef]
- Agut, B.; Pastor, V.; Jaques, J.A.; Flors, V. Can plant defence mechanisms provide new approaches for the sustainable control of the two-spotted spider mite Tetranychus urticae? Int. J. Mol. Sci. 2018, 19, 614. [Google Scholar] [CrossRef] [Green Version]
- Grbic, M.; van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouze, P.; Grbic, V.; Osborne, E.J.; Dermauw, W.; Thi Ngoc, P.C.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Farouk, S.; Osman, M.A. Alleviation of oxidative stress induced by spider mite invasion through application of elicitors in bean plants. Egypt. J. Exp. Biol. 2012, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jena, S.; Choudhuri, M.A. Glycolate metabolism of three submerged aquatic angiosperms during aging. Aquat. Bot. 1981, 12, 345–354. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Effect of light on lipid peroxidation in chloroplasts. Biochem. Biophys. Res. Commun. 1968, 19, 716–720. [Google Scholar] [CrossRef]
- Małolepsza, A.; Urbanek, H.; Polit, J. Some biochemical of strawberry plants to infection with Botrytis cinerea and salicylic acid treatment. Acta Agrobot. 1994, 47, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Chance, B.; Meahly, S.K. Assays of catalase and peroxidase. Meth. Enzymol. 1955, 2, 764–775. [Google Scholar]
- Wiloch, U.; Mioduszewska, H.; Banaś, A. The influence of alloxydim on the antioxidant enzymatic activity in the roots maize (Zea mays L.). Acta Physiol. Plant. 1999, 21, 535–541. [Google Scholar]
- Statistica StatSoft Inc., Poland. Data Analysis Software System Version 13.1. Available online: www.statsoft.com (accessed on 23 July 2020).
- Yano, S.; Wakabayashi, M.; Takabayashi, J.; Takafuji, A. Factors determining the host plant range of the phytophagous mite, Tetranychus urticae (Acari: Tetranychidae): A method for quantifying host plant acceptance. Exp. Appl. Acarol. 1998, 22, 595–601. [Google Scholar] [CrossRef]
- Berenbaum, M.R. Chemical mediation of coevolution: Phylogenetic evidence for Apiaceae and associates. Ann. Missouri Bot. Gard. 2001, 88, 45–59. [Google Scholar] [CrossRef]
- Praslika, J.; Huszar, J. Influence of temperature and host plants on the developmental and fecundity of the spider mite (Tetranychus urticae). Plant Prot. Sci. 2004, 40, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Atalay, E.; Kumral, N.A. Biological features and life tables of Tetranychus urticae (Koch) (Acari: Tetranycidae) on different table tomato varieties. Turk. Entomol. Derg. 2013, 37, 329–341. [Google Scholar]
- Marinosci, C.; Magalhães, S.; Macke, E.; Navajas, M.; Carbonell, D.; Devaux, C.; Olivieri, I. Effects of host plant on life-history traits in the polyphagous spider mite Tetranychus urticae. Ecol. Evol. 2015, 5, 3151–3158. [Google Scholar] [CrossRef]
- Osman, M.A.; Al Dhafar, Z.M.; Alqahtani, A.M. Biological responses of the two-spotted spider mite, Tetranychus urticae to different host plant. Arch. Phytopathol. Pflanzenschutz. 2019, 52, 1229–1238. [Google Scholar] [CrossRef]
- Razmjou, J.; Tavakkoli, H.; Nemati, M. Life history traits of Tetranychus urticae Koch on three legumes (Acari: Tetranychidae). Mun. Ent. Zool. 2009, 4, 204–211. [Google Scholar]
- Golizadeh, A.; Razmjou, J. Life table parameters of Phthorimaea operculella (Lepidoptera: Gelechiidae), feeding on tubers of six potato cultivars. J. Econ. Entomol. 2010, 103, 966–972. [Google Scholar] [CrossRef] [Green Version]
- Golizadeh, A.; Ghavidel, S.; Razmjou, J.; Fathi, S.; Hassanpour, M. Comparative life table analysis of Tetranychus urticae Koch (Acari: Tetranychidae) on ten rose cultivars. Acarologia 2017, 57, 607–616. [Google Scholar]
- Khanamani, M.; Fathipour, Y.; Hajiqanbar, H.; Sedaratian, A. Reproductive performance and life expectancy of Tetranychus urticae (Acari: Tetranychidae) on seven eggplant cultivars. J. Crop Prot. 2012, 1, 57–66. [Google Scholar]
- Shoorooei, M.; Hoseinzadeh, A.H.; Maali-Amiri, R.; Allahyari, H.; Torkzadeh-Mahanti, M. Antixenosis and antibiosis response of common bean (Phaseolus vulgaris) to two-spotted spider mite (Tetranychus uriticae). Exp. Appl. Acarol. 2018, 74, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Van Den Boom, C.E.M.; Beek, T.A.; Dicke, M. Differences among plant species in acceptance by the spider mite Tetranychus urticae Koch. J. Appl. Entomol. 2003, 127, 177–183. [Google Scholar] [CrossRef]
- Hoy, M.A. Agricultural Acarology: Introduction to Integrated Mite Management, 7th ed.; CRC Press Taylor & Francis Group: New York, NY, USA, 2011; pp. 295–296. [Google Scholar]
- Glas, J.J.; Schimmel, B.C.J.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, J.R.F.; de Resende, J.T.V.; Maluf, W.R.; Lucini, T.; de Lima Filho, R.B.; de Lima, I.P.; Nardi, C. Trichomes and allelochemicals in tomato genotypes have antagonistic effects upon behavior and biology of Tetranychus urticae. Front. Plant Sci. 2018, 9, 1132. [Google Scholar] [CrossRef]
- Kaya, A.; Demirci, B.; Baher, K.H.C. Glandular trichomes and essential oil of Salvia glutinosa L. S. Afr. J. Bot. 2003, 69, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Kaya, A.; Goger, F.; Baher, K.H.C. Morphological, anatomical and palynological characteristics of Salvia halophia endemic to Turkey. Nord. J. Bot. 2007, 25, 351–358. [Google Scholar] [CrossRef]
- Shirsat, R.; Kokate, P.; Surdakar, S. Morphological and anatomical characterization of Salvia plebeia from Maharashtra (India). Biosci. Disc. 2012, 3, 165–168. [Google Scholar]
- Oku, K.; Yano, S.; Takafuji, A. Host plant acceptance by the phytophagous mite Tetranychus kanzawai Kishida is affected by the availability of a refuge on the leaf surface. Ecol. Res. 2006, 21, 446–452. [Google Scholar] [CrossRef]
- Kostić, M.; Dražic, S.; Popović, Z.; Stanković, S.; Sivčev, I.; Živanović, T. Developmental and feeding alternations in Leptinotarsa decemlineata Say. (Coleoptera: Chrysomelidae) caused by Salvia officinalis L. (Lamiaceae) essential oil. Biotechnol. Biotechnol. Equip. 2007, 21, 426–430. [Google Scholar] [CrossRef] [Green Version]
- Ebadollahi, A.; Parchin, R.A.; Farjaminezhad, M. Phytochemistry, toxicity and feeding inhibitory activity of Melissa officinalis L. essential oil against a cosmopolitan insect pest; Tribolium castaneum Herbst. Toxin Rev. 2016, 35, 77–82. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential Oils Extracted from Different Species of the Lamiaceae Plant Family as Prospective Bioagents against Several Detrimental Pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalski, R.; Kowalska, G.; Jankowska, M.; Nawrocka, A.; Kałwa, K.; Pankiewicz, U.; Włodarczyk-Stasiak, M. Secretory structures and essential oil composition of selected industrial species of Lamiaceae. Acta Sci. Pol-Hortoru 2019, 18, 53–69. [Google Scholar] [CrossRef]
- Juliani, H.R.; Simon, J.E. Antioxidant Activity of Basil; Trends in new crops and new uses; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002. [Google Scholar]
- Pavela, R. Repellent effect of ethanol extracts from plants of the family Lamiaceae on Colorado Potato Beetle adults (Leptinotarsa decemlineata Say.). Natl. Acad. Sci. Lett. 2004, 27, 195–203. [Google Scholar]
- Ali, A.; Tabanca, N.; Demirci, B.; Blythe, E.K.; Ali, Z.; Can Baser, H.; Khan, I.A. Chemical composition and biological activity of four Salvia essential oils and individual compounds against two species of mosquitoes. J. Agric. Food Chem. 2015, 63, 447–456. [Google Scholar] [CrossRef]
- Won-Il, C.; Sang-Geui, L.; Hyung-Man, P.; Young Joon, A. Toxicity of Plant Essential Oils to Tetranychus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis (Acari: Phytoseiidae). J. Econ. Entomol. 2004, 97, 553–558. [Google Scholar]
- Al-Assiuty, B.A.; Nenaah, G.E.; Ageba, M.E. Chemical profile, characterization and acaricidal activity of essential oils of three plant species and their nanoemulsions against Tyrophagus putrescentiae, a stored-food mite. Exp. Appl. Acarol. 2019, 79, 359–376. [Google Scholar] [CrossRef] [PubMed]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Biol. 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Harborne, J.B. Introduction to Ecological Biochemistry, 4th ed.; Academic Press: London, UK; San Diego, CA, USA, 1994; p. 243. [Google Scholar]
- Cuvelier, M.E.; Richard, H.; Berset, C. Antioxidative activity and phenolic composition of pilot-plant and commercial extracts of sage and rosemary. JAOCS 1996, 73, 645–652. [Google Scholar] [CrossRef]
- Kerchev, P.I.; Fenton, B.; Foyer, C.H.; Hancock, R.D. Infestation of potato (Solanum tuberosum L.) by the peach-potato aphid (Myzus persicae Sulzer) alters cellular redox status and is influenced by ascorbate. Plant Cell Environ. 2012, 35, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Takeda, S.; Mochizuki, S.; Jindal, R.; Dhalla, N.S. Mechanisms of Hydrogen Peroxide-Induced Increase in Intracellular Calcium in Cardiomyocytes. JCPT 1999, 4, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Tan, J.; Zhuo, C.; Wang, C.; Xiang, B.; Wang, Z. Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. 2014, 12, 601–612. [Google Scholar] [CrossRef]
- Łukasik, I.; Kornacka, A.; Goławska, S.; Sytykiewicz, H.; Sprawka, I.; Wójcicka, A. Effects of Acyrtosiphon pisum (Harris) infestation on the hydrogen peroxide content and activity of antioxidant enzymes in Fabaceae plants. Allelopath. J. 2017, 40, 143–150. [Google Scholar] [CrossRef]
- Huag, W.; Zhikuan, J.; Qingfang, H. Effects of herbivore stress by Aphis medicaginis Koch on the malondialdehyde contents and activities of protective enzymes in different alfalfa varieties. Acta Ecol. Sin. 2007, 27, 2177–2183. [Google Scholar]
- Khattab, H. The defense mechanism of cabbage plant against phloem-sucking aphid (Brevicoryne brassicae L.). AJBAS 2007, 1, 56–62. [Google Scholar]
- Tayefi-Nasrabadi, H.; Dehghan, G.; Daeihassani, B.; Movafegi, A.; Samadi, A. Some biochemical properties of guaiacol peroxidases as modified by salt stress in leaves of salt-tolerant and salt-sensitive safflower (Carthamus tinctorius L.cv.) cultivars. Afr. J. Biotechnol. 2011, 10, 751–763. [Google Scholar]
- Biswas, S.M.; Chakraborty, N.; Baidyanath, P. Foliar gall and antioxidant enzyme responses in Alstonia scholaris, R. Br. after psylloid herbivory—An experimental and statistical analysis. Glob. J. Bot. 2014, 2, 12–20. [Google Scholar]
- Mohase, L.; van der Westhuizen, A.J. Salicylic acid is involved in resistant response in the Russia wheat aphid-wheat interaction. J. Plant Physiol. 2002, 159, 585–590. [Google Scholar] [CrossRef]


{kind=link}
| Host Plant | Number of Mites/1 cm2 of Leaves Mean (SE) (%) | Damage of Leaves after 14 Days Feeding Mean (SE) (%) | Number of Leaf Trichomes/1 mm2 Mean (SE) |
|---|---|---|---|
| Ocimum basilicum L. | 3.98 (±0.18) a | 47.36 (±0.49) a | 5.17 (±0.02) c |
| Melissa officinalis L. | 1.02 (±0.048) b | 39.53 (±0.29) b | 72.99 (±0.13) b |
| Salvia officinalis L. | 0.28 (±0.02) c | 17.30 (±0.15) c | 89.90 (±0.38) a |
| H (2, N = 45) P | 39.13 1 × 10−14 | 39.13 1 × 10−14 | 39.13 1 × 10−14 |
| Plant Species | Periods in Days | Total Fecundity | Daily Rate | ||
|---|---|---|---|---|---|
| Oviposition Period | Female Adult Longevity | Life Cycle | |||
| Ocimum basilicum L. | 10.81 (±0.12) b | 13.96 (±0.12) c | 24.30 (±0.20) c | 55.81 (±0.13) a | 5.16 (±0.08) a |
| Melissa officinalis L. | 13.67 (±0.20) a | 17.20 (±0.12) b | 26.43 (±0.65) b | 43.18 (±0.21) b | 3.16 (±0.05) b |
| Salvia officinalis L. | 14.01 (±0.13) a | 18.34 (±0.08) a | 29.22 (±0.53) a | 29.12 (±0.03) c | 2.08 (±0.12) c |
| Kruskal Wallis’ test H (2, N = 45) P | 29.80 1 × 10−14 | 38.45 1 × 10−14 | 36.71 1 × 10−14 | 36.70 1 × 10−14 | 39.15 1 × 10−14 |
| Treatments 1 | Host Plant 2 | ||
|---|---|---|---|
| O. basilicum | M. officinalis | S. officinalis | |
| Control | 8.86 (±0.01) aA | 2.52 (±0.25) aB | 3.33 (±0.17) aAB |
| 1 day | 36.14 (±0.02) abA | 3.26 (±0.13) aB | 6.05 (±0.45) aAB |
| 7 days | 49.36(±0.01) bA | 2.71 (±0.19) aB | 3.67 (±0.16) aAB |
| 14 days | 29.64 (±0.02) abA | 2.83 (±0.03) aB | 6.81 (±0.50) aAB |
| Treatments 1 | Host Plant 2 | ||
|---|---|---|---|
| O. basilicum | M. officinalis | S. officinalis | |
| Control | 4.03 (±0.00) bA | 3.25 (±0.16) bA | 3.15 (±0.14) bA |
| 1 day | 14.84 (±0.02) abA | 3.87 (±0.12) abB | 4.57 (±0.21) abAB |
| 7 days | 15.81 (±0.00) aA | 4.08 (±0.21) abB | 4.89 (±0.14) abAB |
| 14 days | 13.74 (±0.02) abA | 5.89 (±0.05) aAB | 5.47 (±0.34) aB |
| Treatments 1 | Guaiacol Peroxidase Activity (GPX) 2 | ||
|---|---|---|---|
| O. basilicum | M. officinalis | S. officinalis | |
| Control | 1.62 (±0.01) bAB | 0.29 (±0.05) bA | 4.14 (±0.19) bB |
| 1 day | 3.73 (±0.02) abA | 4.09 (±0.56) abA | 58.38 (±3.74) aA |
| 7 days | 6.16 (±0.02) abB | 6.74 (±0.34) aB | 54.24 (±1.84) abA |
| 14 days | 9.98 (±0.00) aAB | 2.96 (±0.03) abA | 27.07 (±1.42) abA |
| Treatments 1 | Catalase Activity (CAT) 2 | ||
|---|---|---|---|
| O. basilicum | M. officinalis | S. officinalis | |
| Control | 0.05 (±0.02) aA | 0.13 (±0.02) aA | 0.18 (±0.04) aA |
| 1 day | 0.03 (±0.00) aB | 0.14 (±0.02) aAB | 0.19 (±0.02) aA |
| 7 days | 0.04 (±0.02) aA | 0.14 (±0.02) aA | 0.06 (±0.02) aA |
| 14 days | 0.02 (±0.00) aB | 0.17 (±0.02) aA | 0.05 (±0.00) aAB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golan, K.; Jurado, I.G.; Kot, I.; Górska-Drabik, E.; Kmieć, K.; Łagowska, B.; Skwaryło-Bednarz, B.; Kopacki, M.; Jamiołkowska, A. Defense Responses in the Interactions between Medicinal Plants from Lamiaceae Family and the Two-Spotted Spider Mite Tetranychus urticae Koch (Acari: Tetranychidae). Agronomy 2021, 11, 438. https://doi.org/10.3390/agronomy11030438
Golan K, Jurado IG, Kot I, Górska-Drabik E, Kmieć K, Łagowska B, Skwaryło-Bednarz B, Kopacki M, Jamiołkowska A. Defense Responses in the Interactions between Medicinal Plants from Lamiaceae Family and the Two-Spotted Spider Mite Tetranychus urticae Koch (Acari: Tetranychidae). Agronomy. 2021; 11(3):438. https://doi.org/10.3390/agronomy11030438
Chicago/Turabian StyleGolan, Katarzyna, Inmaculada Garrido Jurado, Izabela Kot, Edyta Górska-Drabik, Katarzyna Kmieć, Bożena Łagowska, Barbara Skwaryło-Bednarz, Marek Kopacki, and Agnieszka Jamiołkowska. 2021. "Defense Responses in the Interactions between Medicinal Plants from Lamiaceae Family and the Two-Spotted Spider Mite Tetranychus urticae Koch (Acari: Tetranychidae)" Agronomy 11, no. 3: 438. https://doi.org/10.3390/agronomy11030438