
Micronutrients and Soil Microorganisms in the Suppression of Potato Common Scab

Abstract
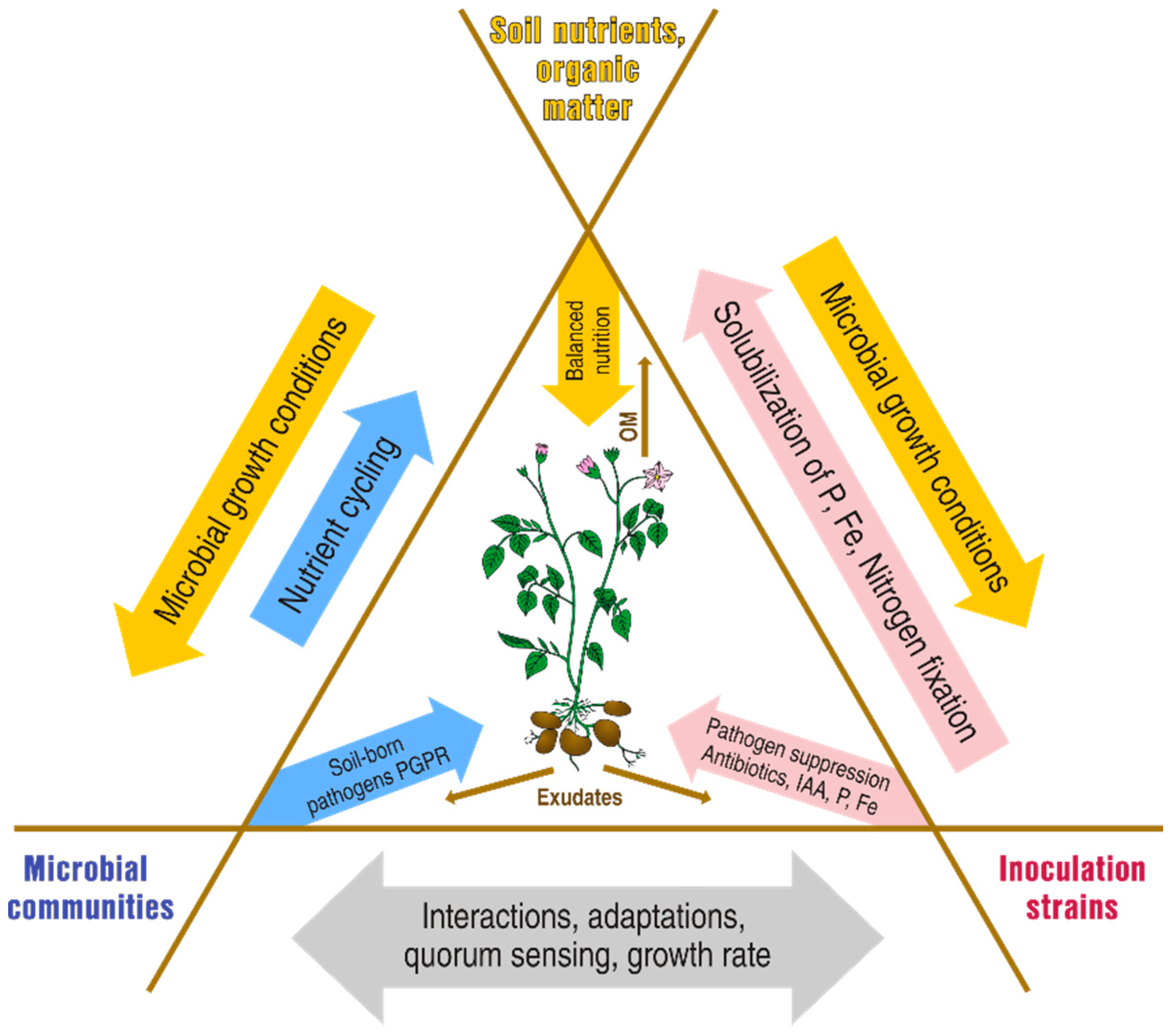
:1. Introduction
2. Individual Nutrients Affect CS Severity
2.1. Micronutrients
2.1.1. Calcium (Ca)
2.1.2. Sulphur (S)
2.1.3. Magnesium (Mg)
2.1.4. Manganese (Mn)
2.1.5. Iron (Fe)
2.2. Macronutrients
2.2.1. Nitrogen (N)
2.2.2. Phosphorus (P)
2.2.3. Potassium
3. Organic Matter Modifies Microbial Communities and Increases Antibiosis
4. Rhizosphere Microorganisms Influence Nutrient Recycling and Produce Beneficial Metabolites
5. Inoculation with Antagonistic Strains
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, A.; Latif, Z.; Zhang, S.; Zhu, J.; Zechel, D.L.; Bechthold, A. Biological control of potato common scab with rare isatropolone C compound produced by plant growth promoting Streptomyces A1RT. Front. Microbiol. 2018, 9, 1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Hooda, P.S. Trace Elements in Soils; John Wiley & Sons Ltd.: Chichester, UK, 2010. [Google Scholar]
- Aznar, A.; Chen, N.W.G.; Thomine, S.; Dellagi, A. Immunity to plant pathogens and iron homeostasis. Plant Sci. 2015, 240, 90–97. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2020, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Huber, D.M.; Jones, J.B. The role of magnesium in plant disease. Plant Soil 2013, 368, 73–85. [Google Scholar] [CrossRef]
- Rosen, C.J.; Kelling, K.A.; Stark, J.C.; Porter, G.A. Optimizing phosphorus fertilizer management in potato production. Am. J. Potato Res. 2014, 91, 145–160. [Google Scholar] [CrossRef]
- Spann, T.M.; Schumann, A.W. Mineral Nutrition Contributes to Plant Disease and Pest Resistance; HS1181, One of a Series of the Horticultural Sciences Department; University of Florida/IFAS Extension: Gainesville, FL, USA, 2013. [Google Scholar]
- Abujabhah, I.S.; Bound, S.A.; Doyle, R.; Bowman, J.P. Effects of biochar and compost amendments on soil physico-chemical properties and the total community within a temperate agricultural soil. Appl. Soil Ecol. 2016, 98, 243–253. [Google Scholar] [CrossRef]
- Gao, Z.; Karlsson, I.; Geisen, S.; Kowalchuk, G.; Jousset, A. Protists: Puppet masters of the rhizosphere microbiome. Trends Plant Sci. 2019, 24, 165–176. [Google Scholar] [CrossRef]
- Lambert, D.H.; Powelson, M.L.; Stevenson, W.R. Nutritional interactions influencing diseases of potato. Am. J. Potato Res. 2005, 82, 309–319. [Google Scholar] [CrossRef]
- Lazarovits, G.; Hill, J.; Patterson, G.; Conn, K.L.; Crump, N.S. Edaphic soil levels of mineral nutrients, pH, organic matter, and cationic exchange capacity in the geocaulosphere associated with potato common scab. Phytopathology 2007, 97, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.R.; McDole, R.E.; Callihan, R.H. Fertilizer effects on common scab of potato and the relation of calcium and phosphate-phosphorus. Phytopathology 1976, 66, 1236–1241. [Google Scholar] [CrossRef]
- Soltani, N.; Conn, K.L.; Abbasi, P.A.; Lazarovits, G. Reduction of potato scab and verticillium wilt with ammonium lignosulfonate soil amendment in four Ontario potato fields. Can. J. Plant Pathol. 2002, 24, 332–339. [Google Scholar] [CrossRef]
- Lazarovits, G. Managing soilborne disease of potatoes using ecologically based approaches. Am. J. Potato Res. 2010, 87, 401–411. [Google Scholar] [CrossRef]
- Bailey, K.L.; Lazarovits, G. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Conn, K.L.; Lazarovits, G. Effect of fish emulsion used as a preplanting soil amendment on verticillium wilt, scab, and tuber yield of potato. Can. J. Plant Pathol. 2006, 28, 509–518. [Google Scholar] [CrossRef]
- Kyselková, M.; Moënne-Loccoz, Y. Pseudomonas and other microbes in disease-suppressive soils. In Organic Fertilisation, Soil Quality and Human Health, Sustainable Agriculture Reviews 9; Lichtfouse, E., Ed.; Springer Science+Business Media B.V.: Dordrecht, The Netherlands, 2012; Volume 9, pp. 93–140. [Google Scholar] [CrossRef]
- Shi, W.; Li, M.; Wei, G.; Tian, R.; Li, C.; Wang, B.; Lin, R.; Shi, C.; Chi, X.; Zhou, B.; et al. The occurrence of potato common scab correlates with the community composition and function of the geocaulosphere soil microbiome. Microbiome 2019, 7, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Andreote, F.D.; Rocha, U.N.D.; Araújo, W.L.; Azevedo, J.L.; Van Overbeek, L.S. Effect of bacterial inoculation, plant genotype and developmental stage on root-associated and endophytic bacterial communities in potato (Solanum tuberosum). Antonie van Leeuwenhoek 2010, 97, 389–399. [Google Scholar] [CrossRef]
- Manter, D.K.; Delgado, J.A.; Holm, D.G.; Stong, R.A. Pyrosequencing reveals a highly diverse and cultivar-specific bacterial endophyte community in potato roots. Microb. Ecol. 2010, 60, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Gevens, A.; Charkowski, A.; Allen, C.; Jansky, S. Potato common scab: A review of the causal pathogens, management practices, varietal resistance screening methods, and host resistance. Am. J. Potato Res. 2017, 94, 283–296. [Google Scholar] [CrossRef]
- Sagova-Mareckova, M.; Daniel, O.; Omelka, M.; Kristufek, V.; Divis, J.; Kopecky, J. Determination of factors associated with natural soil suppressivity to potato common scab. PLoS ONE 2015, 10, e0116291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopecky, J.; Samkova, Z.; Sarikhani, E.; Kyselková, M.; Omelka, M.; Kristufek, V.; Divis, J.; Grundmann, G.G.; Moënne-Loccoz, Y.; Sagova-Mareckova, M. Bacterial, archaeal and micro-eukaryotic communities characterize a disease-suppressive or conducive soil and a cultivar resistant or susceptible to common scab. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Neeno-Eckwall, E.C.; Kinkel, L.L.; Schottel, J.L. Competition and antibiosis in the biological control of potato scab. Can. J. Microbiol. 2001, 47, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kobayashi, Y.O.; Someya, N.; Ikeda, S. Community analysis of root-and tuber-associated bacteria in field-grown potato plants harboring different resistance levels against common scab. Microbes Environ. 2015, 30, 301–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease suppressive soils: New insights from the soil microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- Sagova-Mareckova, M.; Omelka, M.; Kopecky, J. Sequential analysis of soil factors related to common scab of potatoes. FEMS Microbiol. Ecol. 2017, 93, fiw201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarikhani, E.; Sagova-Mareckova, M.; Omelka, M.; Kopecky, J. The effect of peat and iron supplements on the severity of potato common scab and bacterial community in tuberosphere soil. FEMS Microbiol. Ecol. 2017, 93, fiw206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, B.G.; Horneck, D.A.; MacGuidwin, A.E. Improving phosphorus use efficiency through potato rhizosphere modification and extension. Am. J. Potato Res. 2014, 91, 161–174. [Google Scholar] [CrossRef]
- Lambert, D.H.; Manzer, F.E. Relationship of calcium to potato scab. Phytopathology 1991, 81, 632–636. [Google Scholar] [CrossRef]
- Krištůfek, V.; Diviš, J.; Dostálková, I.; Kalčík, J. Accumulation of mineral elements in tuber periderm of potato cultivars differing in susceptibility to common scab. Potato Res. 2000, 43, 107–114. [Google Scholar] [CrossRef]
- Lacey, M.J.; Wilson, C.R. Relationship of common scab incidence of potatoes grown in Tasmanian ferrosol soils with pH, exchangeable cations and other chemical properties of those soils. J. Phytopathol. 2001, 149, 679–683. [Google Scholar] [CrossRef]
- Schroeder, R.A.; Albrecht, W.A. Plant nutrition and the hydrogen ion: II. Potato scab. Soil Sci. 1942, 53, 481–488. [Google Scholar] [CrossRef]
- Wiechel, T.J.; Crump, N.S.; Gilkes, R.J. Soil nutrition and common scab disease of potato in Australia. In Proceedings of the 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; pp. 1–6. [Google Scholar]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The importance of nutrient management for potato production Part I: Plant nutrition and yield. Potato Res. 2020, 63, 97–119. [Google Scholar] [CrossRef] [Green Version]
- Krauss, A. Balanced nutrition and biotic stress. In Proceedings of the IFA Agricultural Conference on Managing Plant Nutrition, Barcelona, Spain, 29 June–2 July 1999. [Google Scholar]
- Loria, R.; Kers, J.; Joshi, M. Evolution of plant pathogenicity in Streptomyces. Annu. Rev. Phytopathol. 2006, 44, 469–487. [Google Scholar] [CrossRef]
- Busse, J.S.; Palta, J.P. Investigating the in vivo calcium transport path to developing potato tuber using 45Ca: A new concept in potato tuber calcium nutrition. Physiol. Plant 2006, 128, 313–323. [Google Scholar] [CrossRef]
- Krištůfek, V.; Diviš, J.; Omelka, M.; Kopecký, J.; Sagová-Marečková, M. Site, year and cultivar effects on relationships between periderm nutrient contents and common scab severity. Am. J. Potato Res. 2015, 92, 473–482. [Google Scholar] [CrossRef]
- Loria, R.; Bukhalid, R.A.; Fry, B.A.; King, R.R. Plant pathogenicity in the genus Streptomyces. Plant Dis. 1997, 81, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Klikocka, H. Influence of NPK fertilization enriched with S, Mg, and micronutrients contained in liquid fertilizer Insol 7 on potato tubers yield (Solanum tuberosum L.) and infestation of tubers with Streptomyces scabies and Rhizoctonia solani. J. Elem. 2009, 14, 271–288. [Google Scholar] [CrossRef]
- Davis, J.R.; Garner, J.G.; Callihan, R.H. Effects of gypsum, sulfur, terraclor and terraclor super-x for potato scab control. Am. Potato J. 1974, 51, 35–43. [Google Scholar] [CrossRef]
- Klikocka, H.; Haneklaus, S.; Bloem, E.; Schnug, E. Influence of sulfur fertilization on infection of potato tubers with Rhizoctonia solani and Streptomyces scabies. J. Plant Nutr. 2005, 28, 819–833. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Mirleau, P. The role of soil microbes in plant sulphur nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inceoǧlu, Ö.; Salles, J.F.; van Elsas, J.D. Soil and cultivar type shape the bacterial community in the potato rhizosphere. Microb. Ecol. 2012, 63, 460–470. [Google Scholar] [CrossRef] [Green Version]
- Bloem, E.; Haneklaus, S.; Schnug, E. Milestones in plant sulfur research on sulfur-induced-resistance (SIR) in Europe. Front. Plant Sci. 2015, 5, 779. [Google Scholar] [CrossRef] [PubMed]
- Klikocka, H.; Kobialka, A.; Juszczak, D.; Glowacka, A. The influence of sulphur on phosphorus and potassium content in potato tubers (Solanum tuberosum L.). J. Elem. 2015, 20, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Senbayram, M.; Gransee, A.; Wahle, V.; Thiel, H. Role of magnesium fertilisers in agriculture: Plant–soil continuum. Crop. Pasture Sci. 2015, 66, 1219–1229. [Google Scholar] [CrossRef]
- Gransee, A.; Führs, H. Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil 2013, 368, 5–21. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. Marschner’s Mineral. Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Shaul, O. Magnesium transport and function in plants: The tip of the iceberg. BioMetals 2002, 15, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.M.; Wilhelm, N.S. The role of manganese in resistance to plant diseases. In Manganese in Soils and Plants. Developments in Plant and Soil Sciences; Graham, R.D., Hannam, R.J., Uren, N.C., Eds.; Springer: Dordrecht, The Netherlands, 1988; Volume 33, pp. 155–173. [Google Scholar] [CrossRef]
- McGregor, A.J.; Wilson, G.C.S. The influence of manganese on the development of potato scab. Plant Soil 1966, 25, 3–16. [Google Scholar] [CrossRef]
- Barnes, E.D. The effects of irrigation, manganese sulphate and sulphur applications on common scab of the potato. Rec. Agric. Res. 1972, 20, 35–44. [Google Scholar]
- Orlov, D.S. Soil Chemistry; A.A. Balkema Publishers: Rotterdam, The Netherlands, 1992. [Google Scholar]
- Mukhopadhyay, M.J.; Sharma, A. Manganese in cell metabolism of higher plants. Bot. Rev. 1991, 57, 117–149. [Google Scholar] [CrossRef]
- Westermann, D.T. Nutritional requirements of potatoes. Am. J. Potato Res. 2005, 82, 301–307. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants: Third Edition; Marschner, P., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Huber, D.M.; Haneklaus, S. Managing nutrition to control plant disease. Landbauforsch. Volkenrode 2007, 57, 313–322. [Google Scholar]
- Rogers, P.F. Organic manuring for potato scab control and its relation to soil manganese. Ann. Appl. Biol. 1969, 63, 371–378. [Google Scholar] [CrossRef]
- Roy, R.N.; Finck, A.; Blair, G.J.; Tandon, H.L.S. Plant Nutrition for Food Security: A Guide for Integrated Nutrient Management; FAO: Rome, Italy, 2006; Volume 16. [Google Scholar]
- Rout, G.R.; Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, C.; Palumbo, G.; He, J.-Z.; Pinton, R.; Cesco, S. Review on iron availability in soil: Interaction of Fe minerals, plants, and microbes. J. Soils Sediments 2014, 14, 538–548. [Google Scholar] [CrossRef]
- Adamski, J.M.; Peters, J.A.; Danieloski, R.; Bacarin, M.A. Excess iron-induced changes in the photosynthetic characteristics of sweet potato. J. Plant Physiol. 2011, 168, 2056–2062. [Google Scholar] [CrossRef]
- Öztürk, E.; Atsan, E.; Polat, T.; Kara, K. Variation in heavy metal concentrations of potato (Solanum tuberosum L.) cultivars. J. Anim. Plant Sci. 2011, 21, 235–239. [Google Scholar]
- Kramer, J.; Özkaya, Ö.; Kümmerli, R. Bacterial siderophores in community and host interactions. Nat. Rev. Microbiol. 2020, 18, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Seipke, R.F.; Song, L.; Bicz, J.; Laskaris, P.; Yaxley, A.M.; Challis, G.L.; Loria, R. The plant pathogen Streptomyces scabies 87-22 has a functional pyochelin biosynthetic pathway that is regulated by TetR- and AfsR-family proteins. Microbiology 2011, 157, 2681–2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodani, S.; Bicz, J.; Song, L.; Deeth, R.J.; Ohnishi-Kameyama, M.; Yoshida, M.; Ochi, K.; Challis, G.L. Structure and biosynthesis of scabichelin, a novel tris-hydroxamate siderophore produced by the plant pathogen Streptomyces scabies 87.22. Org. Biomol. Chem. 2013, 11, 4686–4694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, R.D.; Webb, M.J. Micronutrients and disease resistance and tolerance in plants. In Micronutrients in Agriculture; Mortvedt, J.J., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1991; pp. 329–370. [Google Scholar] [CrossRef]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- van der Wal, A.; de Boer, W. Dinner in the dark: Illuminating drivers of soil organic matter decomposition. Soil Biol. Biochem. 2017, 105, 45–48. [Google Scholar] [CrossRef]
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by rhizobia—The roots of a success story. Curr. Opin. Plant Biol. 2018, 44, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Gaby, J.C.; Buckley, D.H. A global census of nitrogenase diversity. Environ. Microbiol. 2011, 13, 1790–1799. [Google Scholar] [CrossRef]
- Clark, D.R.; McKew, B.A.; Dong, L.F.; Leung, G.; Dumbrell, A.J.; Stott, A.; Grant, H.; Nedwell, D.B.; Trimmer, M.; Whitby, C. Mineralization and nitrification: Archaea dominate ammonia-oxidising communities in grassland soils. Soil Biol. Biochem. 2020, 143, 107725. [Google Scholar] [CrossRef]
- Holloway, J.M.; Dahlgren, R.A. Nitrogen in rock: Occurrences and biogeochemical implications. Glo. Biogeochem. Cycles 2002, 16. [Google Scholar] [CrossRef]
- Lazarovits, G.; Conn, K.L.; Potter, J. Reduction of potato scab, verticillium wilt, and nematodes by soymeal and meat and bone meal in two Ontario potato fields. Can. J. Plant Pathol. 1999, 21, 345–353. [Google Scholar] [CrossRef]
- Conn, K.L.; Lazarovits, G. Impact of animal manures on verticillium wilt, potato scab, and soil microbial populations. Can. J. Plant Pathol. 1999, 21, 81–92. [Google Scholar] [CrossRef]
- Ospina, C.A.; Lammerts van Bueren, E.T.; Allefs, J.J.H.M.; Engel, B.; van der Putten, P.E.L.; van der Linden, C.G.; Struik, P.C. Diversity of crop development traits and nitrogen use efficiency among potato cultivars grown under contrasting nitrogen regimes. Euphytica 2014, 199, 13–29. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalali, M. Phosphorus availability as influenced by organic residues in five calcareous soils. Compost Sci. Util. 2009, 17, 241–246. [Google Scholar] [CrossRef]
- Hammond, J.P.; Broadley, M.R.; White, P.J. Genetic responses to phosphorus deficiency. Ann. Bot. 2004, 94, 323–332. [Google Scholar] [CrossRef]
- Johnson, J.F.; Vance, C.P.; Allan, D.L. Phosphorus deficiency in Lupinus albus (altered lateral root development and enhanced expression of phosphoenolpyruvate carboxylase). Plant Physiol. 1996, 112, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2019, 133, 146–159. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, X.; Chen, W.; Huang, Q. Isolation and identification of three potassium-solubilizing bacteria from rape rhizospheric soil and their effects on ryegrass. Geomicrobiol. J. 2017, 34, 873–880. [Google Scholar] [CrossRef]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Bernard, E.; Larkin, R.P.; Tavantzis, S.; Erich, M.S.; Alyokhin, A.; Gross, S.D. Rapeseed rotation, compost and biocontrol amendments reduce soilborne diseases and increase tuber yield in organic and conventional potato production systems. Plant Soil 2014, 374, 611–627. [Google Scholar] [CrossRef]
- Larkin, R.P.; Honeycutt, C.W.; Griffin, T.S.; Olanya, O.M.; Halloran, J.M.; He, Z. Effects of different potato cropping system approaches and water management on soilborne diseases and soil microbial communities. Phytopathology 2011, 101, 58–67. [Google Scholar] [CrossRef]
- Mazzola, M. Mechanisms of natural soil suppressiveness to soilborne diseases. Antonie van Leeuwenhoek 2002, 81, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Al-Mughrabi, K.I.; Bertheleme, C.; Livingston, T.; Burgoyne, A.; Poirier, R.; Vikram, A. Aerobic compost tea, compost and a combination of both reduce the severity of common scab (Streptomyces scabiei) on potato tubers. J. Plant Sci. 2008, 3, 168–175. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop. Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Wilson, C.; Zebarth, B.J.; Goyer, C.; Burton, D.L. Effect of diverse compost products on soilborne diseases of potato. Compost Sci. Util. 2018, 26, 156–164. [Google Scholar] [CrossRef]
- Ninh, H.T.; Grandy, A.S.; Wickings, K.; Snapp, S.S.; Kirk, W.; Hao, J. Organic amendment effects on potato productivity and quality are related to soil microbial activity. Plant Soil 2014, 386, 223–236. [Google Scholar] [CrossRef]
- Curless, M.A.; Kelling, K.A.; Speth, P.E.; Stevenson, W.R.; James, R.V. Effect of manure application timing on potato yield, quality, and disease incidence. Am. J. Potato Res. 2012, 89, 363–373. [Google Scholar] [CrossRef]
- Alabouvette, C.; Hoeper, H.; Lemanceau, P. Soil suppressiveness to diseases induced by soilborne plant pathogens. In Soil Biochemistry; Stotzky, G., Bollag, J.-M., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1996; Volume 9, pp. 371–413. [Google Scholar]
- Brevik, E.C.; Cerdà, A.; Mataix-Solera, J.; Pereg, L.; Quinton, J.N.; Six, J.; Van Oost, K. The interdisciplinary nature of soil. Soil 2015, 1, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.J.; Liu, H.; Donis-Gonzalez, I.R.; Lu, X.H.; Jones, D.; Fulbright, D.W. Antimicrobial activity of chestnut extracts for potential use in managing soilborne plant pathogens. Plant Dis. 2012, 96, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Millard, W.A. Common scab of potatoes. Ann. Appl. Biol. 1923, 10, 70–88. [Google Scholar] [CrossRef]
- Larkin, R.P.; Lynch, R.P. Use and effects of different brassica and other rotation crops on soilborne diseases and yield of potato. Horticulturae 2018, 4, 37. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, F.; Maeda, M.; Takahashi, M.; Hashizume, K.; Kondo, N. Suppression of common scab of potato caused by Streptomyces turgidiscabies using lopsided oat green manure. Plant Dis. 2011, 95, 1124–1130. [Google Scholar] [CrossRef] [Green Version]
- Tomihama, T.; Nishi, Y.; Mori, K.; Shirao, T.; Iida, T.; Uzuhashi, S.; Ohkuma, M.; Ikeda, S. Rice bran amendment suppresses potato common scab by increasing antagonistic bacterial community levels in the rhizosphere. Phytopathology 2016, 106, 719–728. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.K.; Srivastava, J.S. Soil amendments to control common scab of potato. Potato Res. 2004, 47, 101–109. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.A.M.; Hassan, E.A.; Elbana, E.A.; Galal, A.A. Management of potato common scab caused by Streptomyces scabies using compost and compost tea. Minia J. Agric. Res. Dev. 2016, 36, 293–305. [Google Scholar]
- Roy, J.; Lafontaine, P.; Chabot, R.; Beaulieu, C. Dehydrated pork manure by-product: Effect of a chitosan amendment on bacterial community and common scab incidence. Phytoprotection 2009, 90, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Larkin, R.P.; Honeycutt, C.W.; Griffin, T.S.; Olanya, O.M.; He, Z.; Halloran, J.M. Cumulative and residual effects of different potato cropping system management strategies on soilborne diseases and soil microbial communities over time. Plant Pathol. 2017, 66, 437–449. [Google Scholar] [CrossRef]
- Sun, P.; Otto-Hanson, L.K.; Arenz, B.E.; Ma, Q.; Kinkel, L.L. Molecular and functional characteristics of streptomycete communities in relation to soil factors and potato common scab. Eur. J. Soil Biol. 2015, 70, 58–66. [Google Scholar] [CrossRef]
- Schlatter, D.; Fubuh, A.; Xiao, K.; Hernandez, D.; Hobbie, S.; Kinkel, L. Resource amendments influence density and competitive phenotypes of Streptomyces in soil. Microb. Ecol. 2009, 57, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, B.E.; Kinkel, L.L. Green manures and crop sequences influence potato diseases and pathogen inhibitory activity of indigenous streptomycetes. Phytopathology 2005, 95, 178–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinkel, L.L.; Schlatter, D.C.; Bakker, M.G.; Arenz, B.E. Streptomyces competition and co-evolution in relation to plant disease suppression. Res. Microbiol. 2012, 163, 490–499. [Google Scholar] [CrossRef]
- Fernández-Romero, M.L.; Clark, J.M.; Collins, C.D.; Parras-Alcántara, L.; Lozano-García, B. Evaluation of optical techniques for characterising soil organic matter quality in agricultural soils. Soil Tillage Res. 2016, 155, 450–460. [Google Scholar] [CrossRef]
- Dippenaar, B.J. Environmental and Control Studies of the Common Scab Disease of Potatoes Caused by Actinomyces Scabies (Thaxt.) Guss. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 1933. [Google Scholar]
- Wach, M.J.; Krasnoff, S.B.; Loria, R.; Gibson, D.M. Effect of carbohydrates on the production of thaxtomin A by Streptomyces acidiscabies. Arch. Microbiol. 2007, 188, 81–88. [Google Scholar] [CrossRef]
- Jourdan, S.; Francis, I.M.; Kim, M.J.; Salazar, J.J.C.; Planckaert, S.; Frère, J.M.; Matagne, A.; Kerff, F.; Devreese, B.; Loria, R.; et al. The CebE/MsiK transporter is a doorway to the cello-oligosaccharide-mediated induction of Streptomyces scabies pathogenicity. Sci. Rep. 2016, 6, 27144. [Google Scholar] [CrossRef] [PubMed]
- Deflandre, B.; Thiébaut, N.; Planckaert, S.; Jourdan, S.; Anderssen, S.; Hanikenne, M.; Devreese, B.; Francis, I.; Rigali, S. Deletion of bglC triggers a genetic compensation response by awakening the expression of alternative beta-glucosidase. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194615. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N. Embracing the unknown: Disentangling the complexities of the soil microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, S.A.; Griffiths, J.; Ton, J. Crying out for help with root exudates: Adaptive mechanisms by which stressed plants assemble health-promoting soil microbiomes. Curr. Opin. Microbiol. 2019, 49, 73–82. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Geddes, B.A.; Paramasivan, P.; Joffrin, A.; Thompson, A.L.; Christensen, K.; Jorrin, B.; Brett, P.; Conway, S.J.; Oldroyd, G.E.D.; Poole, P.S. Engineering transkingdom signalling in plants to control gene expression in rhizosphere bacteria. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guyonnet, J.P.; Guillemet, M.; Dubost, A.; Simon, L.; Ortet, P.; Barakat, M.; Heulin, T.; Achouak, W.; Haichar, F.E.Z. Plant nutrient resource use strategies shape active rhizosphere microbiota through root exudation. Front. Plant Sci. 2018, 9, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef]
- Huber, D.; Römheld, V.; Weinmann, M. Relationship between nutrition, plant diseases and pests. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 283–298. [Google Scholar] [CrossRef]
- Gu, Y.; Wei, Z.; Wang, X.; Friman, V.-P.; Huang, J.; Wang, X.; Mei, X.; Xu, Y.; Shen, Q.; Jousset, A. Pathogen invasion indirectly changes the composition of soil microbiome via shifts in root exudation profile. Biol. Fertil. Soils 2016, 52, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Lorang, J.M.; Liu, D.; Anderson, N.A.; Schottel, J.L. Identification of potato scab inducing and suppressive species of Streptomyces. Phytopathology 1995, 85, 261–268. [Google Scholar] [CrossRef]
- Meng, Q.X.; Jiang, H.H.; Hanson, L.E.; Hao, J.J. Characterizing a novel strain of Bacillus amyloliquefaciens BAC03 for potential biological control application. J. Appl. Microbiol. 2012, 113, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, N.; Tiedje, J.M.; Quensen, J.F.; Meng, Q.; Hao, J.J. Microbial communities associated with potato common scab-suppressive soil determined by pyrosequencing analyses. Plant Dis. 2012, 96, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Almario, J.; Kyselková, M.; Kopecký, J.; Ságová-Marečková, M.; Muller, D.; Grundmann, G.L.; Moënne-Loccoz, Y. Assessment of the relationship between geologic origin of soil, rhizobacterial community composition and soil receptivity to tobacco black root rot in Savoie region (France). Plant Soil 2013, 371, 397–408. [Google Scholar] [CrossRef]
- Tegg, R.S.; Gill, W.M.; Thompson, H.K.; Davies, N.W.; Ross, J.J.; Wilson, C.R. Auxin-induced resistance to common scab disease of potato linked to inhibition of thaxtomin A toxicity. Plant Dis. 2008, 92, 1321–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiore-Donno, A.M.; Weinert, J.; Wubet, T.; Bonkowski, M. Metacommunity analysis of amoeboid protists in grassland soils. Sci. Rep. 2016, 6, 19068. [Google Scholar] [CrossRef]
- Jousset, A. Application of protists to improve plant growth in sustainable agriculture. In Rhizotrophs: Plant Growth Promotion to Bioremediation; Mehnaz, S., Ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2017; pp. 263–273. [Google Scholar] [CrossRef]
- Weinert, N.; Piceno, Y.; Ding, G.C.; Meincke, R.; Heuer, H.; Berg, G.; Schloter, M.; Andersen, G.; Smalla, K. PhyloChip hybridization uncovered an enormous bacterial diversity in the rhizosphere of different potato cultivars: Many common and few cultivar-dependent taxa. FEMS Microbiol. Ecol. 2011, 75, 497–506. [Google Scholar] [CrossRef]
- Hu, H.W.; Zhang, L.M.; Yuan, C.L.; He, J.Z. Contrasting Euryarchaeota communities between upland and paddy soils exhibited similar pH-impacted biogeographic patterns. Soil Biol. Biochem. 2013, 64, 18–27. [Google Scholar] [CrossRef]
- Meng, Q.X.; Yin, J.F.; Rosenzweig, N.; Douches, D.; Hao, J.J. Culture-based assessment of microbial communities in soil suppressive to potato common scab. Plant Dis. 2012, 96, 712–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanner, L.A. High proportions of nonpathogenic Streptomyces are associated with common scab-resistant potato lines and less severe disease. Can. J. Microbiol. 2007, 53, 1062–1075. [Google Scholar] [CrossRef] [PubMed]
- Roquigny, R.; Novinscak, A.; Léger, G.; Marcoux, N.; Joly, D.L.; Filion, M. Deciphering the rhizosphere and geocaulosphere microbiomes of potato following inoculation with the biocontrol agent Pseudomonas fluorescens strain LBUM223. Phytobiomes J. 2018, 2, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Ardanov, P.; Ovcharenko, L.; Zaets, I.; Kozyrovska, N.; Pirttilä, A.M. Endophytic bacteria enhancing growth and disease resistance of potato (Solanum tuberosum L.). Biol. Control 2011, 56, 43–49. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Jobin, G.; Couture, G.; Goyer, C.; Brzezinski, R.; Beaulieu, C. Streptomycete spores entrapped in chitosan beads as a novel biocontrol tool against common scab of potato. Appl. Microbiol. Biotechnol. 2005, 68, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.O.; Kobayashi, A.; Maeda, M.; Takenaka, S. Isolation of antagonistic Streptomyces sp. against a potato scab pathogen from a field cultivated with wild oat. J. Gen. Plant Pathol. 2012, 78, 62–72. [Google Scholar] [CrossRef]
- Lee, H.B.; Cho, J.W.; Park, D.J.; Li, C.T.; Ko, Y.H.; Song, J.H.; Koh, J.S.; Kim, B.J.; Kim, C.J. In vivo screening for biocontrol agents (BCAs) against Streptomyces scabiei causing potato common scab. Plant Pathol. J. 2004, 20, 110–114. [Google Scholar] [CrossRef]
- Sarwar, A.; Latif, Z.; Zhang, S.; Hao, J.; Bechthold, A. A potential biocontrol agent Streptomyces violaceusniger AC12AB for managing potato common scab. Front. Microbiol. 2019, 10, 202. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, Y.; Zhuang, L.; Yu, Y.; Liu, J.; Zhang, L.; Gao, Z.; Wu, Y.; Gao, W.; Ding, G.; et al. A rhizosphere-derived consortium of Bacillus subtilis and Trichoderma harzianum suppresses common scab of potato and increases yield. Comput. Struct. Biotechnol. J. 2019, 17, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Schmiedeknecht, G.; Bochow, H.; Junge, H. Use of Bacillus subtilis as biocontrol agent. II. Biological control of potato diseases. J. Plant Dis. Prot. 1998, 105, 376–386. [Google Scholar]
- Chen, S.; Zhang, M.; Wang, J.; Lv, D.; Ma, Y.; Zhou, B.; Wang, B. Biocontrol effects of Brevibacillus laterosporus AMCC100017 on potato common scab and its impact on rhizosphere bacterial communities. Biol. Control 2017, 106, 89–98. [Google Scholar] [CrossRef]
- Arseneault, T.; Goyer, C.; Filion, M. Pseudomonas fluorescens LBUM223 increases potato yield and reduces common scab symptoms in the field. Phytopathology 2015, 105, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhai, P.K.; Sarma, B.K.; Srivastava, J.S. Biological management of common scab of potato through Pseudomonas species and vermicompost. Biol. Control 2011, 57, 150–157. [Google Scholar] [CrossRef]
- Larkin, R.P.; Tavantzis, S. Use of biocontrol organisms and compost amendments for improved control of soilborne diseases and increased potato production. Am. J. Potato Res. 2013, 90, 261–270. [Google Scholar] [CrossRef]
- Kobayashi, Y.O.; Kobayashi, A.; Soejima, H.; Takenaka, S. Enhanced suppressive effect of antagonistic Streptomyces sp. WoRs-501 on potato scab in conjunction with other control methods. Jpn. Agric. Res. Q. 2017, 51, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, Y.O.; Kobayashi, A.; Maeda, M.; Someya, N.; Takenaka, S. Biological control of potato scab and antibiosis by antagonistic Streptomyces sp. WoRs-501. J. Gen. Plant Pathol. 2015, 81, 439–448. [Google Scholar] [CrossRef]
- Han, J.S.; Cheng, J.H.; Yoon, T.M.; Song, J.; Rajkarnikar, A.; Kim, W.G.; Yoo, I.D.; Yang, Y.Y.; Suh, J.W. Biological control agent of common scab disease by antagonistic strain Bacillus sp. sunhua. J. Appl. Microbiol. 2005, 99, 213–221. [Google Scholar] [CrossRef]
- Cui, L.; Yang, C.; Wei, L.; Li, T.; Chen, X. Isolation and identification of an endophytic bacteria Bacillus velezensis 8-4 exhibiting biocontrol activity against potato scab. Biol. Control 2020, 141, 104156. [Google Scholar] [CrossRef]
- Nanri, N.; Gohda, Y.; Ohno, M.; Miyabe, K.; Furukawa, K.; Hayashida, S. Growth promotion of fluorescent pseudomonads and control of potato common scab in field soil with non-antibiotic actinomycete-biofertilizer. Biosci. Biotechnol. Biochem. 1992, 56, 1289–1292. [Google Scholar] [CrossRef] [Green Version]
- Wanner, L.A.; Kirk, W.W.; Qu, X.S. Field efficacy of nonpathogenic Streptomyces species against potato common scab. J. Appl. Microbiol. 2014, 116, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, L.H.; Kelloniemi, J.; Valkonen, J.P.T. Repeated applications of a nonpathogenic Streptomyces strain enhance development of suppressiveness to potato common scab. Plant Dis. 2017, 101, 224–232. [Google Scholar] [CrossRef]
- Arseneault, T.; Goyer, C.; Filion, M. Biocontrol of potato common scab is associated with high Pseudomonas fluorescens LBUM223 populations and phenazine-1-carboxylic acid biosynthetic transcript accumulation in the potato geocaulosphere. Phytopathology 2016, 106, 963–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagoz, K.; Dadasoglu, F.; Mohammadi, P.; Kotan, R. Screening bacterial antagonists to common scab disease. J. Anim. Plant Sci. 2018, 28, 1068–1073. [Google Scholar]
- Lin, L.; Xu, X.; Zheng, Y.; Zhang, C. Effects of AttM lactonase on the pathogenicity of Streptomyces scabies. Lett. Appl. Microbiol. 2018, 67, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Goyer, C. Isolation and characterization of phages Stsc1 and Stsc3 infecting Streptomyces scabiei and their potential as biocontrol agents. Can. J. Plant Pathol. 2005, 27, 210–216. [Google Scholar] [CrossRef]
- St-Onge, R.; Goyer, C.; Filion, M. Pseudomonas spp. can inhibit Streptomyces scabies growth and repress the expression of genes involved in pathogenesis. J. Bacteriol. Parasitol. 2010, 1, 101. [Google Scholar] [CrossRef] [Green Version]
- Arseneault, T.; Roquigny, R.; Novinscak, A.; Goyer, C.; Filion, M. Phenazine-1-carboxylic acid-producing Pseudomonas synxantha LBUM223 alters the transcriptome of Streptomyces scabies, the causal agent of potato common scab. Physiol. Mol. Plant Pathol. 2020, 110, 101480. [Google Scholar] [CrossRef]
- Liu, Q.; Shen, Y.R.; Yin, K.D. The degradation fragments of gamma-glutamyl transpeptidase from Bacillus subtilis BU108 have antimicrobial activity against Streptomyces scabiei. J. Plant Dis. Prot. 2020. [Google Scholar] [CrossRef]
- Agbessi, S.; Beauséjour, J.; Déry, C.; Beaulieu, C. Antagonistic properties of two recombinant strains of Streptomyces melanosporofaciens obtained by intraspecific protoplast fusion. Appl. Microbiol. Biotechnol. 2003, 62, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Schottel, J.L.; Shimizu, K.; Kinkel, L.L. Relationships of in vitro pathogen inhibition and soil colonization to potato scab biocontrol by antagonistic Streptomyces spp. Biol. Control 2001, 20, 102–112. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.W.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.; Zhao, X.; Shangguan, N.; Chang, D.; Ma, Q. The roles of inoculants’ carbon source use in the biocontrol of potato scab disease. Can. J. Microbiol. 2015, 61, 257–262. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Element | Factors of Availability to Plants | Function in Plant | Relation to Plant Defense/Pathogenesis in General | Possible Relation to CS | Main References |
|---|---|---|---|---|---|
| Cation Exchange Capacity (CEC) | Higher CEC in clay than sandy soils; low CEC and Ca especially correlated with low pH | A measure of soil capacity to hold nutrients (Ca, Mg, K, Na, Al and H) | Different effects of Ca, Mg, K and their ratio; imbalanced nutrients promote impaired biosynthesis and accumulation of low-molecular weight substances readily available for parasites | CS disease severity related to exchangeable Ca, Mg and K cations; CEC is lower in suppressive soil; the higher CEC, the greater uptake of Ca2+; uptake of monovalent cations (K+) increases at lower CEC | [5,37,60,66] |
| K | Leached out in acid soils; Al dominates the CEC, limits the soil’s ability to absorb and hold K | Control of cation–anion homeostasis, membrane polarization, more than 60 enzymes in photosynthesis and transport of photosynthetic products to storage organs; starch synthesis; increased plant resistance to pests, diseases and abiotic stresses | Correlated with K:Mg, K:Ca and K:N ratios; decrease in some plant diseases if N and P are sufficient; high content of N increases plant susceptibility to diseases—this adverse effect can be neutralized by balanced N:K ratios of fertilizers | Special Ca:K ratio reduces scab severity and incidence, while the imbalance or excess of K or Ca promotes the disease | [38,40,64,66] |
| Ca | pH | Stability and function of membranes and cell walls; second messenger triggered by different stimuli including pathogens | At low Ca level, cells leak compounds used as food by parasites; supports some pathogens by stimulating the action of pectolytic enzymes dissolving plant cell wall; inhibits the activity of other pectolytic enzymes | Ca may simulate the aerial mycelium formation and spore germination of S. scabiei; high calcium levels in the absence of changes in pH induce scab | [35,40,41,66] |
| Mg | Mg deficiency can be induced by higher K supply | Allosteric activator of more than 300 enzymes; a central part of chlorophyll molecules; in the structural integrity of cell components | Decreases the susceptibility to pathogen-produced macerating enzymes as long as Ca level remains sufficient | Soils suppressive to CS disease had a higher content of Mg; the CS-resistant potato cultivar has a higher Mg content in periderm | [27,28,40,53] |
| Mn | More available with lower pH; Mn uptake increased by seed inoculation with pseudomonads, organic amendments increased Mn-reducing microorganisms | Co-factor of enzymatic activity/an inhibitor at high concentrations; control of lignin and suberin biosynthesis, phenol biosynthesis, photosynthesis | Mn is required at much higher concentrations by higher plants than by fungi and bacteria | High Mn correlates with low CS; S. scabiei oxidize Mn making it unavailable for the plant; soil amendments reducing Mn availability (liming, nitrate fertilizers) increase CS severity; herbicide glyphosate, toxic to Mn reducing organisms | [55,57] |
| Fe | Predominant ferric ion (Fe+3) is sparingly soluble; more available at lower pH as reduced form Fe+2; bacteria supply iron to plants and backward | DNA synthesis, respiration, photosynthesis; in prosthetic groups of many enzymes (cytochromes); synthesis of chlorophyll; essential for maintenance of chloroplast structure and function | Plants, bacteria and fungi compete for Fe in the rhizosphere; microorganisms have lower Fe requirements than plants; promotes antifungals’ production by soil bacteria for plant benefit; activates both enzymes involved in the infection and in plant defense | S. scabiei produces siderophores desferrioxamine, scabichelin and pyochelin to compete for iron; lower CS severity in soils with more available iron; higher Fe in the periderm of tubers grown in suppressive soil; enrichment with available iron and peat suppressed CS | [5,27,33,67,68,69,70] |
| Material | Quantity | Method | Effect | Reference |
|---|---|---|---|---|
| Compost | 0.4 mt ha−1 | CS suppression 42% | [95] | |
| Aerobic compost tea | 140 L ha−1 | Back-pack sprayer | CS suppression 81% | |
| Indian mustard green manure | In the year before potatoes | CS suppression 25% | [96] | |
| Barley/ryegrass rotations | Rotated prior to potatoes | CS suppression 13–34% | [104] | |
| Lopsided oat (Avena strigosa) | 150 kg ha−1 | Rotovator prior to potatoes | CS suppression | [105] |
| Rice bran | 3 t ha−1 | In the furrow with the seed tubers | CS suppression (DS) | [106] |
| Soybean green manure, with wheat straw | Incorporated before planting | CS suppression | [107] | |
| Green manure (Brassica napus) | Grown for approx. 2 months in the fall | Decrease in CS incidence and severity | [92] | |
| Compost | 2.5 kg m−2 | 5 days before planting | CS suppression: 61% to 28% DI, 30% to 16% DS a | [108] |
| Compost tea | 1 kg m−2 in 5 L water | Soil drenching | CS suppression: 61% to 32% DI, 30% to 15% DS | |
| Fish emulsion | 1% | Soil amendment | No effect | [21] |
| Processed swine manure | 2 g per seed | Seed covering | Increased DI | [109] |
| Processed swine manure with chitosan | 2 g per seed; 1% chitosan | Seed covering | no effect | |
| Chicken manure | 66 t ha−1 | Incorporated to 15-cm depth | CS suppression | [83] |
| Swine manure | 5.5 m3 ha−1 | CS suppression | ||
| Dairy cattle manure | 100 t ha−1 | No effect | ||
| Poultry manure | 1.54–12.32 Mg C ha−1 | Applied dehydrated, pelletized prior to planting potatoes | CS suppression | [98] |
| Poultry manure, forestry residues and organic waste compost | 45 Mg ha−1 | Incorporated to a depth of 15 cm in October, a year before planting | CS suppression | [97] |
| Meat and bone meal | 37 t ha−1 | Incorporated to 15-cm depth | CS suppression | [16] |
| Soymeal | 37 t ha−1 | CS suppression | ||
| Poultry manure | 66 t ha−1 | CS suppression | ||
| Poultry manure | 20 t ha−1 | No effect | ||
| Nature Safe 10-2-8 (poultry feathers) | 10 t ha−1 | CS suppression by 50–100% | ||
| Ammonium lignosulfonate | 1000 L ha−1 | Marketable yield 60–80% compared to 10% in the control | ||
| Peat | 2.5 L peat /10 L soil | In submerged pots | CS suppression | [33] |
| Manipulation | Affected Bacterial Groups | Reference |
|---|---|---|
| Suppressive soil | Higher frequency of antagonistic pseudomonads and streptomycetes and higher proportion of pathogenic streptomycetes by cultivation; different microbial community by terminal restriction fragment analysis | [137] |
| Suppressive soil | Higher proportion of Acetobacteraceae, Bacillaceae and Lysobacter (Xanthomonadaceae), lower Acidobacteria group 6, Nocardioidaceae, Pseudomonadaceae, unclassified Acidobacteria group 11 and unclassified Bacilli | [130] |
| Suppressive soil | Enriched in Pseudomonadaceae, Bradyrhizobiaceae, Acetobacteraceae and Paenibacillaceae; decreased ratio of Thaumarchaeota/Euryarchaeota in tuberosphere of susceptible cultivar; higher proportions of Ascomycota and Basidiomycota and lower proportions of Chlorophyta, Ciliophora, Myxogastria and Apicomplexa | [28] |
| Resistant cultivar | Same as in suppressive soil and enriched in Nitrospirae and Acidobacteria; decreased tuberosphere ratio of Thaumarchaeota/Euryarchaeota in conducive soil; Chlorophyta and Cercozoa in lower proportions | |
| Resistant cultivar | More non-pathogenic streptomycetes than in susceptible cultivar | [138] |
| Resistant cultivar | Cloning/Sanger sequencing analysis of root, tuber and rhizosphere bacterial communities of 8 cultivars differing in resistance to CS; higher abundance of S. turgidiscabies in susceptible cultivars by qPCR | [30] |
| Susceptible cultivar | Variovorax, Stenotrophomonas and Agrobacterium were positively, and Geobacillus, Curtobacterium and unclassified Geodermatophilaceae negatively, correlated with the scab severity level, estimated absolute abundance of pathogenic Streptomyces and txtAB genes | [23] |
| Biocontrol strain Pseudomonas fluorescens LBUM223 | Inoculation does not significantly alter the autochthonous rhizosphere nor geocaulosphere microbiomes in the field | [139] |
| Rice bran amendment | Bacillus, Streptomyces, Chitinophaga and Actinomadura significantly increased; unclassified Koribacteraceae and unclassified Gaiellaceae were significantly reduced | [106] |
| Iron amendment | Bacillales and Gaiellales | [33] |
| Peat amendment | Rhizobiales, Burkholderiales, Xanthomonadales and Bacillales | |
| Peat and iron amendment | Proteobacteria and Bacteroidetes increased at the proportional expense of Actinobacteria; proteobacterial Burkholderiales, Xanthomonadales and Sphingomonadales and actinobacterial order of Gaiellales were the most responsive groups; actinobacterial families Micromonosporaceae and Thermomonosporaceae were elevated |
| A. Complex Inocula | ||
| Inoculum | Effect | Reference |
| Swine feces with coprophilous actinobacteria | CS suppression; viable counts of Streptomyces scabies decreased, antagonistic fluorescent pseudomonads increased | [156] |
| Broth treatment and inoculation with antagonistic Bacillus sp. in pot assay with sterile soil | CS suppression by 40% | [154] |
| Pseudomonas mosselii with vermicompost | CS suppression | [150] |
| Bacillus subtilis GB03 and Rhizoctonia solani hypovirulent isolate Rhs1A1, compost | CS suppression by 10–34%, increased yield, reduction in stem and stolon canker by 20–38%, black scurf by 30–58% | [151] |
| Trichoderma virens, Bacillus subtilis and Rhizoctonia solani, compost, rapeseed rotation | Reduced disease and increased yield | [92] |
| Vermiculite cultures of non-pathogenic Streptomyces isolates mixed with soil | CS suppression | [157] |
| Conducive soil inoculated with Streptomyces isolates | No effect of inoculation; variability of streptomycete community increasing from planting to mid-season (by pyrosequencing); pathogen suppressive capacity of antagonistic streptomycetes negatively correlating with CS severity (by cultivation) | [111] |
| B. Individual strains | ||
| Inoculum | Effect | Reference |
| Streptomyces melanosporofaciens EF-76 in chitosan beads | Increased numbers of geldanamycin-resistant actinobacteria on harvested potato tubers | [142] |
| Pesticide and antibiotic-resistant Streptomyces spp. | CS suppression by 55–>60% | [144] |
| S. violaceusniger AC12AB | CS reduction up to 90% in greenhouse and field, increased yield up to 26.8% in field trial | [145] |
| Streptomyces sp. WoRs-501 | CS severity decreased by 78–94% in field pot trial | [143] |
| Streptomyces strain 272 | CS severity reduced by 43% on susceptible potato cultivar Bintje, both disease incidence and severity reduced by 43 and 59% on the scab-tolerant cultivar Nicola | [158] |
| Pseudomonas fluorescens LBUM223 | CS reduced by approximately 30% in plots after biweekly applications, increased yield by 46%; did not reduce pathogen soil populations, down-regulated txtA expression in the geocaulosphere | [149,159] |
| Bacillus megaterium KBA-10, P. putida K-19B, B. megaterium TV- 91C, Pantoea agglomerans RK-92 | Biocontrol efficacy 18.7–60.3%, tuber yield increase by 20.4–40% | [160] |
| B. subtilis and Trichoderma harzianum in diatomaceous earth (225–300 kg ha−1) | CS index decreased by 30.6–46.1%; 19-23-fold higher Pseudomonadales; CS severity negatively correlated with relative abundances of Agrobacterium, Achromobacter and Pseudomonas and positively with Acidobacteria, Actinobacteria, Chloroflexi and Gemmatimonadetes | [146] |
| Brevibacillus laterosporus AMCC100017 | CS severity decreased from 2.60 to 0.77, i.e., biocontrol efficacy 70.51%; reduced pathogen, transient impact on the native bacteria community | [148] |
| Bacillus subtilis | Decrease in common scab severity up to 70%, and 67% in field trials | [147] |
| B. amyloliquefaciens Ba01 | CS reduced from 14.4 ± 2.9% (naturally occurring) to 5.6 ± 1.1% in the field | [161] |
| B. velezensis 8-4 | CS control efficiency reached 51.83 ± 8.53%, the yield increased by 19.91 ± 3.56% | [155] |
| Trichoderma virens | Decrease in CS incidence and severity | [92] |
| Phages Stsc1 and Stsc3 | Prevented CS symptoms on radish seedlings | [162] |
| C. Mechanisms of CS suppression | ||
| Biocontrol Strain | Observed Property | Reference |
| Streptomyces violaceusniger AC12AB | Production of azalomycin, indole-3-acetic acid and siderophores, nitrogen fixation and phosphate solubilization => CS reduction up to 90% in greenhouse and field trials, increased yield up to 26.8% in field trial | [145] |
| Pseudomonas fluorescens LBUM223 | Phenazine-1-carboxylic acid (PCA) production => growth inhibition of S. scabiei, repression of thaxtomin biosynthesis genes (txtA and txtC); activities were lost in phzC- mutant deficient in PCA production | [163] |
| Phenazine-1-carboxylic acid production => 12%–14% of all S. scabiei genes were differentially expressed, including key genes involved in pathogenicity/virulence, mycelium differentiation and increased oxidative stress | [164] | |
| Fragments of γ-glutamyl transpeptidase from Bacillus subtilis BU108 | In vitro growth inhibition of S. scabiei | [165] |
| S. melanosporofaciens EF-76 | Geldanamycin production => disease index was reduced from 6.30 to 4.81 and from 2.83 to 2.49 in growth chamber and field experiments, respectively | [166] |
| Streptomyces A1RT | Production of isatropolone C and indole-3-acetic acid => reduction in average disease severity index by 82.4–95.7% | [4] |
| Streptomyces isolates (strains 93 and 63) and their spontaneous mutants | The mutants lost in vitro inhibitory activity against S. scabiei in antibiotic and co-plate assay, while retaining the biocontrol ability in soil | [167] |
| B. amyloliquefaciens subsp. plantarum FZB42 | Production of cyclic lipopeptides and volatiles => pathways of induced systemic resistance | [168] |
| Agrobacterium tumefaciens C58 attM gene introduced to S. scabiei | No disease symptoms in planta, altered morphological differentiation—quorum quenching paralyzing γ-butyrolactone signaling pathway | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kopecky, J.; Rapoport, D.; Sarikhani, E.; Stovicek, A.; Patrmanova, T.; Sagova-Mareckova, M. Micronutrients and Soil Microorganisms in the Suppression of Potato Common Scab. Agronomy 2021, 11, 383. https://doi.org/10.3390/agronomy11020383
Kopecky J, Rapoport D, Sarikhani E, Stovicek A, Patrmanova T, Sagova-Mareckova M. Micronutrients and Soil Microorganisms in the Suppression of Potato Common Scab. Agronomy. 2021; 11(2):383. https://doi.org/10.3390/agronomy11020383
Chicago/Turabian StyleKopecky, Jan, Daria Rapoport, Ensyeh Sarikhani, Adam Stovicek, Tereza Patrmanova, and Marketa Sagova-Mareckova. 2021. "Micronutrients and Soil Microorganisms in the Suppression of Potato Common Scab" Agronomy 11, no. 2: 383. https://doi.org/10.3390/agronomy11020383