
Morphological Characteristics, Fruit Qualities and Evaluation of Reproductive Functions in Autotetraploid Satsuma Mandarin (Citrus unshiu Marcow.)


Abstract
:1. Introduction
2. Materials and Methods
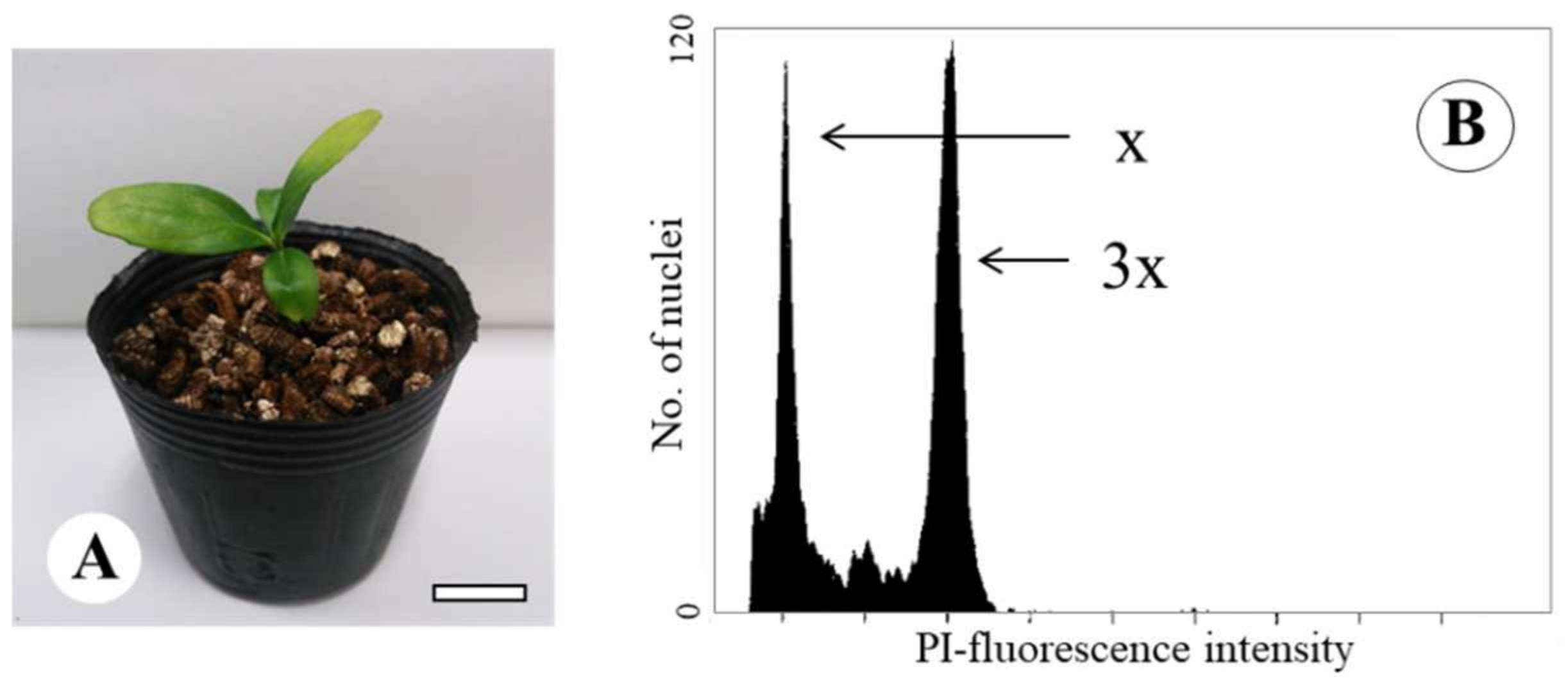
2.1. Plant Materials
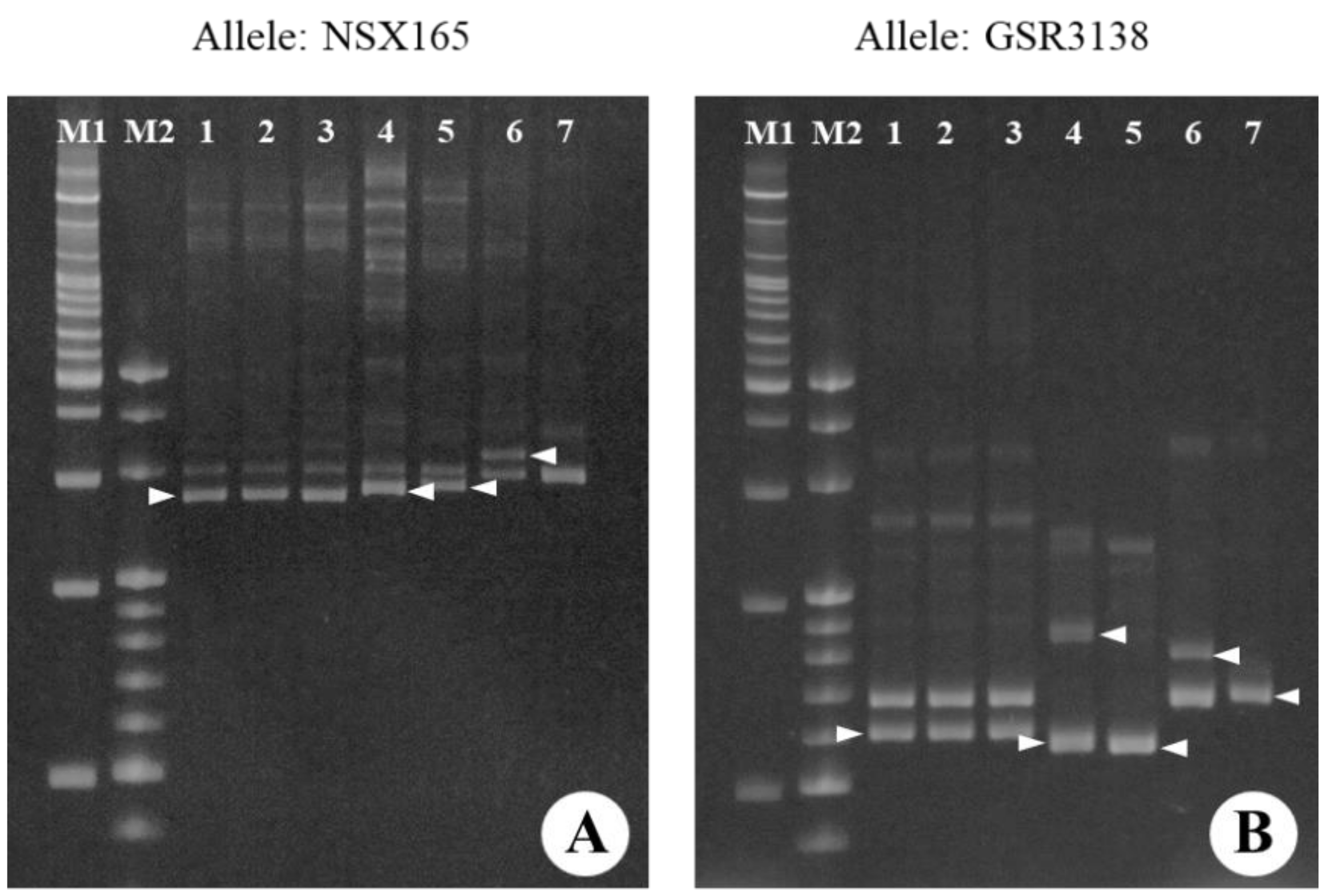
2.2. Confirmation of the Origin of the Tetraploid Satsuma Mandarin by SSR Analysis
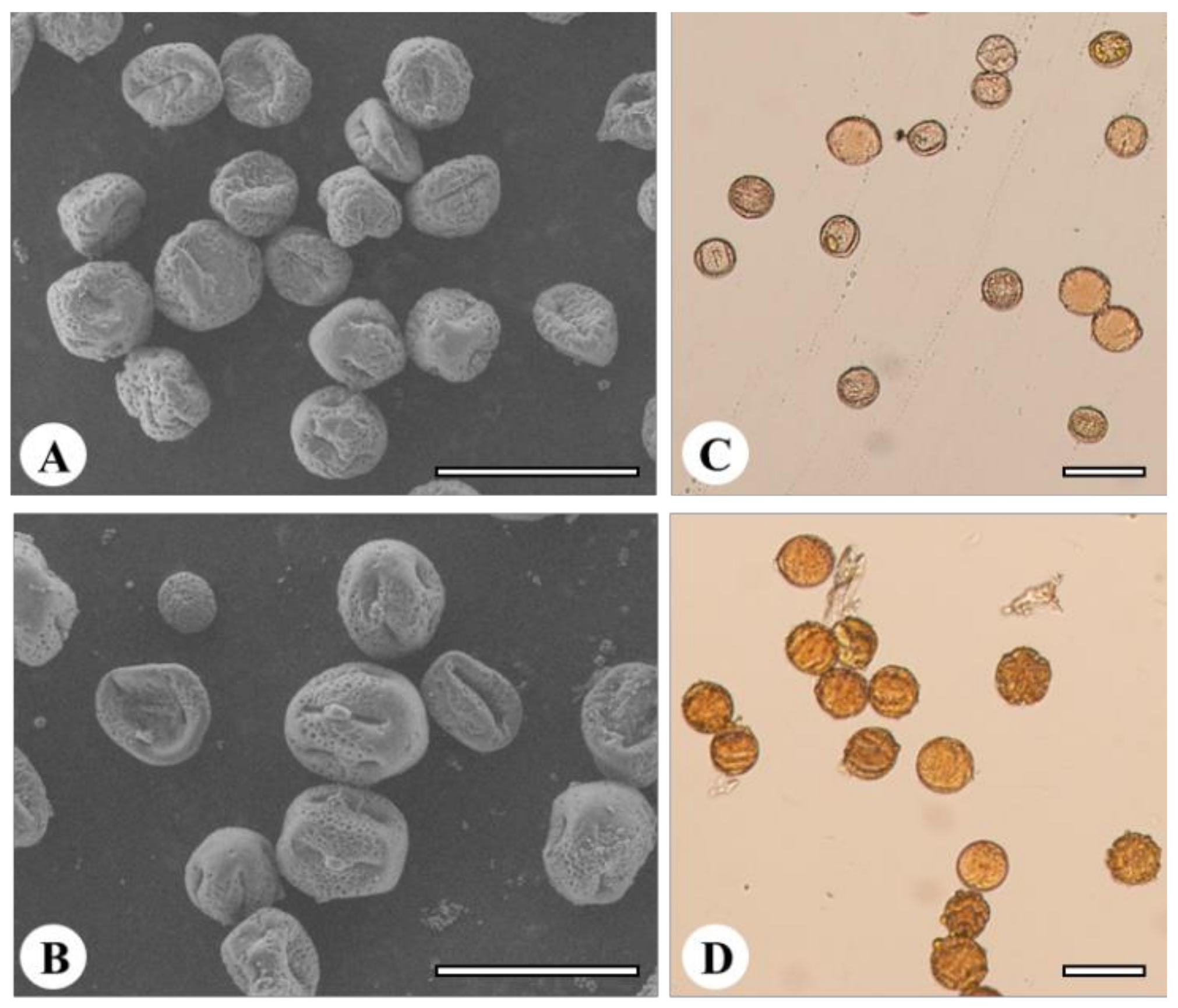
2.3. Morphological Characteristics of Leaf, Flower and Pollen
2.4. Evaluation of Fruit Quality
2.5. Crossing for Evaluation of the Reproductive Functions
3. Results
3.1. Confirmation of the Origin of the Tetraploid Satsuma Mandarin by SSR Analysis
3.2. Morphological Characteristics of Leaf, Flower and Pollen
3.3. Evaluation of Fruit Quality
3.4. Crossing for Evaluation of the Reproductive Functions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ikoma, Y.; Matsumoto, H.; Kato, M. Diversity in the carotenoid profiles and the expression of genes related to carotenoid accumulation among citrus genotypes. Breed. Sci. 2016, 66, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Yoshinaga, A.; Takabe, K.; Yoshioka, T.; Ogawa, K.; Sakamoto, M.; Azuma, J.; Honda, Y. In situ detection and identification of hesperidin crystals in Satsuma mandarin (Citrus unshiu) peel cells. Phytochem. Anal. 2014, 26, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jin, R.; Yang, Z.; Wang, X.; You, G.; Guo, J.; Zhang, Y.; Liu, F.; Pan, S. Comparative study on physicochemical, nutritional and enzymatic properties of two Satsuma mandarin (Citrus unshiu Marc.) varieties from different regions. J. Food Compos. Anal. 2021, 95, 103614. [Google Scholar] [CrossRef]
- Sugiura, M.; Nakamura, M.; Ogawa, K.; Ikoma, Y.; Yano, M. High serum carotenoids associated with lower risk for the metabolic syndrome and its components among Japanese subjects: Mikkabi cohort study. Br. J. Nutr. 2015, 114, 1674–1682. [Google Scholar] [CrossRef] [Green Version]
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. Segregation and heritability of male sterility in populations derived from progeny of Satsuma mandarin. PLoS ONE 2016, 11, e0162408. [Google Scholar] [CrossRef]
- Omura, M.; Shimada, T. Citrus breeding, genetics and genomics in Japan. Breed. Sci. 2016, 66, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, T.; Kuroda, H.; Sugiura, H. Influence of the current state of global warming on fruit tree growth in Japan. Hort. Res. 2007, 6, 257–263, (In Japanese with English abstract). [Google Scholar] [CrossRef] [Green Version]
- Sugiura, T.; Sumida, H.; Yokoyama, S.; Ohno, H. Overview of recent effects of global warming on agricultural production in Japan. Jpn. Agric. Res. Q. JARQ 2012, 46, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Recupero, G.R.; Russo, G.; Recupero, S. New promising Citrus triploid hybrid selected from crosses between monoembryonic diploid female and tetraploid male parents. HortScience 2005, 40, 516–520. [Google Scholar] [CrossRef]
- Soost, R.K.; Cameron, J.W. ‘Oroblanco’, a triploid pummelo-grapefruit hybrid. HortScience 1980, 15, 667–669. [Google Scholar]
- Soost, R.K.; Cameron, J.W. ‘Melogold’, a triploid pummelo-grapefruit hybrid. HortScience 1985, 20, 1134–1135. [Google Scholar]
- Nukaya, T.; Sudo, M.; Yahata, M.; Nakajo, Y.; Ohta, T.; Yasuda, K.; Tominaga, A.; Mukai, H.; Kunitake, H. Characteristics in autotetraploid kumquats (Fortunella spp.) induced by colchicine treatment to nucellar embryos and their utilization for triploid breeding. Sci. Hortic. 2019, 245, 210–217. [Google Scholar] [CrossRef]
- Viloria, Z.; Grosser, J.W. Acid citrus fruit improvement via interploid hybridization using allotetraploid somatic hybrid and autotetraploid breeding parents. J. Amer. Soc. Hortic. Sci. 2005, 130, 392–402. [Google Scholar] [CrossRef]
- Xie, K.D.; Yuan, D.Y.; Wang, W.; Xia, Q.M.; Wu, X.M.; Chen, C.W.; Chen, C.L.; Grosser, J.W.; Guo, W.W. Citrus triploid recovery based on 2x×4x crosses via an optimized embryo rescue approach. Sci. Hortic. 2019, 252, 104–109. [Google Scholar] [CrossRef]
- Nukaya, T.; Yahata, M.; Suzuki, K.; Yasuda, K.; Kunitake, H.; Komatsu, H.; Mukai, H.; Harada, H.; Takagi, T. Fruit characteristics in autotetraploid Meiwa kumquat induced by colchicine treatment to nucellar embryos. Hortic. Environ. Biotechnol. 2009, 50, 188–190. [Google Scholar]
- Tan, F.Q.; Zhang, M.; Xie, K.D.; Fan, Y.J.; Song, X.; Wang, R.; Wu, X.M.; Zhang, H.Y.; Guo, W.W. Polyploidy remodels fruit metabolism by modifying carbon source utilization and metabolic flux in Ponkan mandarin (Citrus reticulata Blanco). Plant Sci. 2019, 289, 110276. [Google Scholar] [CrossRef]
- Tan, F.Q.; Tu, H.; Wang, R.; Wu, X.M.; Xie, K.D.; Chen, J.J.; Zhang, H.Y.; Xu, J.; Guo, W.W. Metabolic adaptation following genome doubling in citrus doubled diploids revealed by non-targeted metabolomics. Metabolomics 2017, 13, 143. [Google Scholar] [CrossRef]
- Wei, T.; Wang, Y.; Liu, J.H. Comparative transcriptome analysis reveals synergistic and disparate defense pathways in the leaves and roots of trifoliate orange (Poncirus trifoliata) autotetraploids with enhanced salt tolerance. Hort. Res. 2020, 7, 88. [Google Scholar] [CrossRef]
- Wei, T.; Wang, Y.; Xie, Z.; Guo, D.; Chen, C.; Fan, Q.; Deng, X.; Liu, J.H. Enhanced ROS scavenging and sugar accumulation contribute to drought tolerance of naturally occurring autotetraploids in Poncirus trifoliata. Plant Biotech. J. 2019, 17, 1394–1407. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities fresh leaf tissue. Phytochemistry Bull. 1987, 19, 11–15. [Google Scholar]
- Shimizu, T.; Kitajima, A.; Nonaka, K.; Yoshioka, T.; Ohta, S.; Goto, S.; Toyoda, A.; Fujiyama, A.; Mochizuki, T.; Nagasaki, H.; et al. Hybrid origins of citrus varieties inferred from DNA marker analysis of nuclear and organelle genomes. PLoS ONE 2016, 11, e0166969. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Yahata, M.; Sato, M.; Sudo, M.; Tominaga, A.; Kunitake, H. Identification of parental genome construction and inherited morphological characteristics in triploid and aneuploid intergeneric hybrids from a diploid−diploid cross between Citrus and Fortunella. Agronomy 2021, 11, 1988. [Google Scholar] [CrossRef]
- Kawase, K.; Hirai, M. Growth, sugar accumulation and puffiness of the mandarin peel during coloring. J. Japan. Soc. Hort. Sci. 1983, 52, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Mukai, H.; Takagi, T.; Teshima, Y.; Suzuki, T. Sugar contents in parts of fruit of satsuma mandarin in tree under water stress in authumn. J. Japan. Soc. Hort. Sci. 1996, 65, 479–484, (In Japanease with English abstract). [Google Scholar] [CrossRef]
- Kato, M.; Ikoma, Y.; Matsumoto, H.; Sugiura, M.; Hyodo, H.; Yano, M. Accumulation of carotenoids and expression of carotenoid biosynthetic genes during maturation in citrus fruit. Plant Physiol. 2004, 134, 824–837. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Yahata, M.; Nukaya, T.; Sudo, M.; Ohta, T.; Yasuda, K.; Inagaki, H.; Mukai, H.; Harada, H.; Takagi, T.; Komatsu, H.; et al. Morphological characteristics of a doubled haploid line from ‘Banpeiyu’ pummelo [Citrus maxima (Burm.) Merr.] and its reproductive function. Hortic. J. 2015, 84, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Yahata, M.; Harusaki, S.; Komatsu, H.; Takami, K.; Kunitake, H.; Yabuya, T.; Yamashita, K.; Toolapong, P. Morphological characterization and molecular verification of a fertile haploid pummelo (Citrus grandis Osbeck). J. Am. Soc. Hort. Sci. 2005, 130, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Lee, L. Citrus polyploidy. -Origin and potential for cultivar improvement. Aust. J. Agric. Res. 1988, 39, 735–747. [Google Scholar] [CrossRef]
- Kato, M. Mechanism of carotenoid accumulation in citrus fruit. J. Japan. Soc. Hort. Sci. 2012, 81, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Li, X.; Zhao, P.; Zhang, X.; Qiao, O.; Huang, L.; Guo, L.; Gao, W. A review of chemical constituents and health-promoting effects of citrus peels. Food Chem. 2021, 365, 130585. [Google Scholar] [CrossRef] [PubMed]
- Nogata, Y.; Sakamoto, K.; Shiratsuchi, H.; Ishii, T.; Yano, M.; Ohta, H. Flavonoid composition of fruit tissues of Citrus species. Biosci. Biotechnol. Biochem. 2006, 70, 178–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, T.; Ueno, Y.; Hamada, Y.; Furukawa, C.; Fujimoto, T.; Yamada, T.; Tanaka, R. Five new limonoids from peels of Satsuma orange (Citrus reticulata). Molecules 2017, 22, 907. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Mitsuhashi, H. Newly Revised Illustrated Medicinal Plants of the World; Hokuryukan Co., Ltd.: Tokyo, Japan, 2002; p. 249. (In Japanese) [Google Scholar]
- Dhawan, O.P.; Lavania, U.C. Enhancing the productivity of secondary metabolites via induced polyploidy: A review. Euphytica 1996, 87, 81–89. [Google Scholar] [CrossRef]
- Iannicelli, J.; Guariniello, J.; Tossi, V.E.; Regalado, J.J.; Di Ciaccio, L.; van Baren, C.M.; Pitta Álvarez, S.I.; Escandon, A.S. The “polyploid effect” in the breeding of aromatic and medicinal species. Sci. Hortic. 2020, 260, 108854. [Google Scholar] [CrossRef]
- Kim, H.E.; Han, J.E.; Lee, H.; Kim, J.H.; Kim, H.H.; Lee, K.Y.; Shin, J.H.; Kim, H.K.; Park, S.Y. Tetraploidization increases the contents of functional metabolites in Cnidium officinale. Agronomy 2021, 11, 1561. [Google Scholar] [CrossRef]
- Madani, H.; Escrich, A.; Hosseini, B.; Sanchez-Muñoz, R.; Khojasteh, A.; Palazon, J. Effect of polyploidy induction on natural metabolite production in medicinal plants. Biomolecules 2021, 11, 899. [Google Scholar] [CrossRef]
- Ravandi, E.G.; Rezanejad, F.; Zolala, J.; Dehghan, E. The effects of chromosome-doubling on selected morphological and phytochemical characteristics of Cichorium intybus L. J. Hortic. Sci. Biotechnol. 2015, 88, 701–709. [Google Scholar] [CrossRef]
- Frost, H.B.; Soost, R.K. The botany of Citrus and its wild relatives in the orange subfamily. In The Citrus Industry; Reuther, W., Bachelor, L.D., Webber, H.J., Eds.; University of California: Berkeley, CA, USA, 1968; Volume II, pp. 290–324. [Google Scholar]
- Osawa, I. Cytological and experimental studies in Citrus. J. Coll. Agr. Univ. Tokyo 1912, 4, 83–116. [Google Scholar]
- Yang, H.J.; Nakagawa, S. 1969. Cyto-histological studies on the male sterility of Satsuma orange (Citrus unshiu Marc.). J. Japan. Soc. Hort. Sci. 1969, 38, 33–39. [Google Scholar] [CrossRef]
- Goto, S.; Yoshioka, T.; Ohta, S.; Kita, M.; Hamada, H.; Shimizu, T. QTL mapping of male sterility and transmission pattern in progeny of Satsuma mandarin. PLoS ONE 2018, 13, e0200844. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Matsumoto, R.; Okudai, N.; Yamada, Y. Aborted anthers of Citrus result from gene-cytoplasmic male sterility. Sci. Hort. 1997, 70, 9–14. [Google Scholar] [CrossRef]
- Oiyama, I.; Kobayashi, S.; Yoshinaga, K.; Ohgawara, T.; Ishii, S. Use of pollen from a somatic hybrid between Citrus and Poncirus in the production of triploids. HortScience 1991, 26, 1082. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Leaf Blade (mm) | Shape Index of Leaf Blade 2 | Guard Cell (µm) | Stoma Density (No. mm−2) | ||
|---|---|---|---|---|---|---|
| Length | Width | Length | Width | |||
| Diploid | 99.4 | 41.7 | 2.4 | 24.3 | 20.2 | 495.7 |
| Tetraploid | 109.9 | 58.6 | 1.9 | 27.9 | 26.1 | 319.3 |
| t-test | ** 1 | ** | ** | * | ** | ** |
| Strains | Flower Bud (mm) | No. of Petals | Petal (mm) | Pistil (mm) | Ovary (mm) | No. of Stamens | |||
|---|---|---|---|---|---|---|---|---|---|
| Length | Width | Length | Width | Height | Diameter | ||||
| Diploid | 19.0 | 7.5 | 5.0 | 19.0 | 7.8 | 19.7 | 5.1 | 3.7 | 14.5 |
| Tetraploid | 19.1 | 10.8 | 4.9 | 19.6 | 9.5 | 21.2 | 5.8 | 5.2 | 15.0 |
| t-test | NS 1 | ** | NS | NS | ** | ** | ** | ** | ** |
| Strains | Pollen Grain (µm) | Shape Index of Pollen Grain 2 | Fertility (%) | ||
|---|---|---|---|---|---|
| Length | Width | Stainability | In Vitro Germination | ||
| Diploid | 29.7 | 23.4 | 1.3 | 56.6 | 1.4 |
| Tetraploid | 42.6 | 32.7 | 1.3 | 75.7 | 5.3 |
| t-test | ** 1 | ** | NS | * | ** |
| Year | Strains | Fruit Weight (g) | Rind Per a Fruit (%) | Fruit (cm) | Shape Index of Fruit 2 | Rind Color | Rind Puffing | No. of Seeds | Soluble Solid Content (%) | Titratable Acidity (%) | Soluble Solid-Acid Ratio 3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diameter | Height | |||||||||||
| 2014 | Diploid Tetraploid | 126.5 | 18.5 | 67.6 | 52.6 | 1.3 | 7.7 | 0.2 | 0 | 13.6 | 0.8 | 17.0 |
| 126.7 | 26.5 | 71.2 | 48.7 | 1.5 | 8.6 | 0.4 | 0 | 10.4 | 1.0 | 10.4 | ||
| t-test | NS 1 | ** | * | ** | ** | ** | NS | NS | ** | ** | ** | |
| 2015 | Diploid | 124.8 | 21.8 | 67.2 | 53.5 | 1.3 | 7.7 | 0.2 | 0 | 11.8 | 0.8 | 14.8 |
| Tetraploid | 128.3 | 28.5 | 70.0 | 50.8 | 1.4 | 8.6 | 0.4 | 0 | 10.4 | 1.0 | 10.4 | |
| t-test | NS | ** | * | * | ** | ** | NS | NS | ** | ** | ** | |
| Year | Strains | Sugar Content (%) | Organic Acid Content (%) | |||||
|---|---|---|---|---|---|---|---|---|
| Sucrose | Glucose | Fructose | Total | Citric Acid | Malic Acid | Total | ||
| 2014 | Diploid | 7.06 | 2.99 | 2.71 | 12.76 | 0.73 | 0.06 | 0.79 |
| Tetraploid | 5.16 | 2.56 | 2.42 | 10.14 | 0.93 | 0.11 | 1.04 | |
| t-test | ** 1 | * | NS | ** | ** | * | * | |
| 2015 | Diploid | 8.98 | 2.29 | 2.28 | 13.55 | 0.58 | 0.10 | 0.68 |
| Tetraploid | 6.72 | 2.15 | 2.25 | 11.11 | 0.76 | 0.18 | 0.95 | |
| t-test | ** | NS | NS | ** | ** | ** | ** | |
| Strains | Juice (µg g−1) | Flavedo (µg g−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| All-trans-Violaxanthin | 9-cis-Violaxanthin | Lutein | β-Cryptoxanthin | Total | All-trans-Violaxanthin | 9-cis-Violaxanthin | Lutein | β-Cryptoxanthin | Total | |
| Diploid | 0.6 | 2.2 | 0.6 | 23.2 | 26.6 | 15.0 | 87.3 | 13.3 | 81.5 | 197.1 |
| Tetraploid | 0.5 | 2.7 | 0.2 | 20.6 | 24.0 | 29.5 | 184.2 | 13.6 | 113.1 | 340.3 |
| t-test | NS 1 | NS | NS | NS | NS | ** | ** | NS | * | ** |
| Cross Combination | No. of Flowers Pollinated | No. of Fruit Set | Fruit Set (%) | No. of Seeds | No. of Developed Seeds Per Fruit 1 | |||
|---|---|---|---|---|---|---|---|---|
| Seed Parent | Pollen Parent | Developed | Undeveloped | Empty | ||||
| Diploid | Banpeiyu pummelo | 20 | 15 | 75.0 | 9 | 0 | 0 | 0.60 |
| Tetraploid | Banpeiyu pummelo | 10 | 4 | 40.0 | 3 | 0 | 0 | 0.75 |
| Harehime | Diploid | 20 | 8 | 40.0 | 1 | 4 | 0 | 0.12 |
| Harehime | Tetraploid | 20 | 12 | 60.0 | 15 | 53 | 0 | 1.25 |
| Cross Combination | Kind of Seed | No. of Seedlings Examined | Ploidy Level | |||
|---|---|---|---|---|---|---|
| Seed Parent | Pollen Parent | 2X | 3X | 4X | ||
| Diploid | Banpeiyu pummelo | Developed | 20 | 20 | 0 | 0 |
| Tetraploid | Banpeiyu pummelo | Developed | 6 | 0 | 2 | 4 |
| Harehime | Diploid | Developed | 1 | 1 | 0 | 0 |
| Undeveloped | 1 | 1 | 0 | 0 | ||
| Harehime | Tetraploid | Developed | 2 | 0 | 0 | 2 |
| Undeveloped | 7 | 0 | 7 | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudo, M.; Yasuda, K.; Yahata, M.; Sato, M.; Tominaga, A.; Mukai, H.; Ma, G.; Kato, M.; Kunitake, H. Morphological Characteristics, Fruit Qualities and Evaluation of Reproductive Functions in Autotetraploid Satsuma Mandarin (Citrus unshiu Marcow.). Agronomy 2021, 11, 2441. https://doi.org/10.3390/agronomy11122441
Sudo M, Yasuda K, Yahata M, Sato M, Tominaga A, Mukai H, Ma G, Kato M, Kunitake H. Morphological Characteristics, Fruit Qualities and Evaluation of Reproductive Functions in Autotetraploid Satsuma Mandarin (Citrus unshiu Marcow.). Agronomy. 2021; 11(12):2441. https://doi.org/10.3390/agronomy11122441
Chicago/Turabian StyleSudo, Miki, Kiichi Yasuda, Masaki Yahata, Mai Sato, Akiyoshi Tominaga, Hiroo Mukai, Gang Ma, Masaya Kato, and Hisato Kunitake. 2021. "Morphological Characteristics, Fruit Qualities and Evaluation of Reproductive Functions in Autotetraploid Satsuma Mandarin (Citrus unshiu Marcow.)" Agronomy 11, no. 12: 2441. https://doi.org/10.3390/agronomy11122441