
Potassium Bioavailability in a Tropical Kaolinitic Soil

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
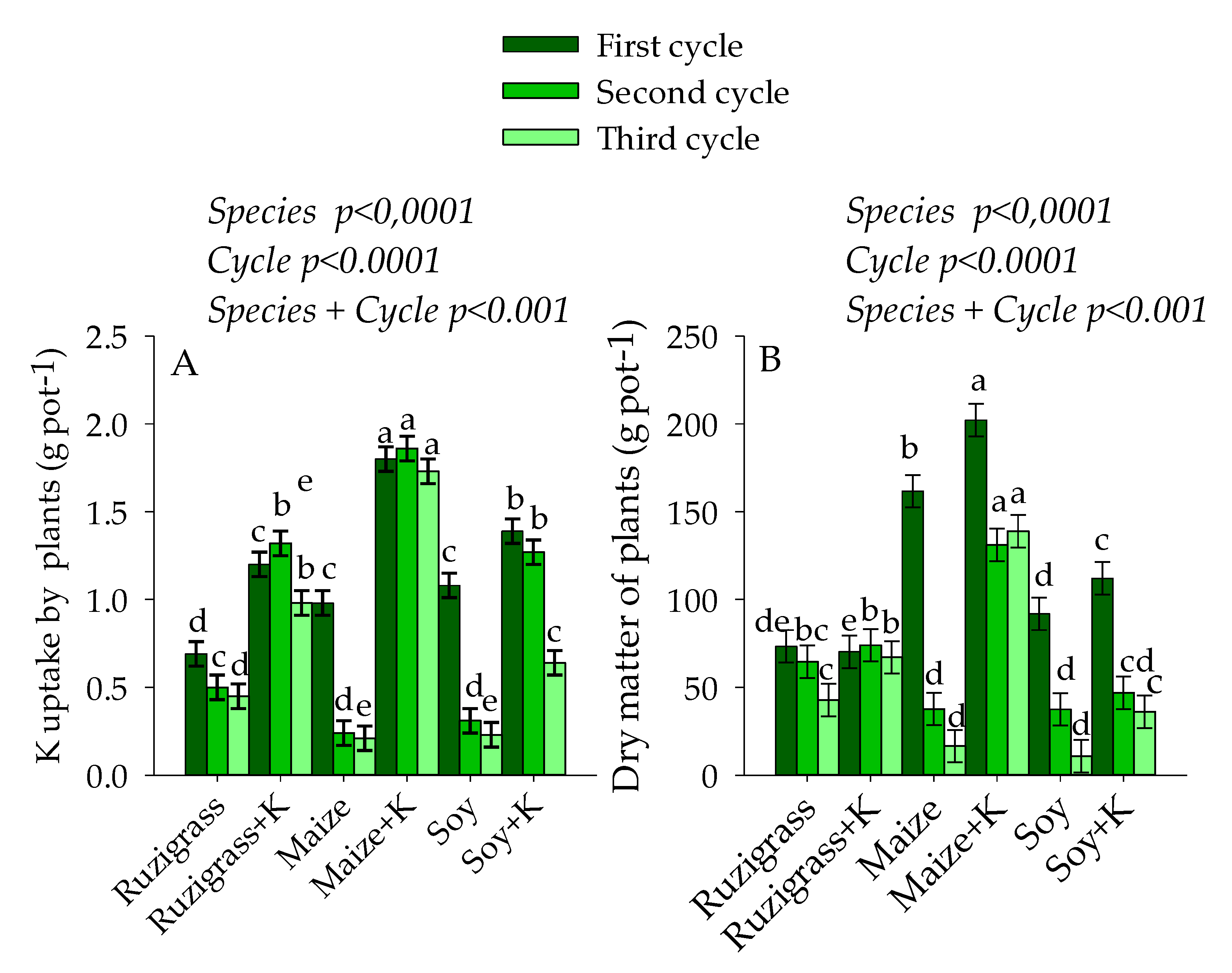
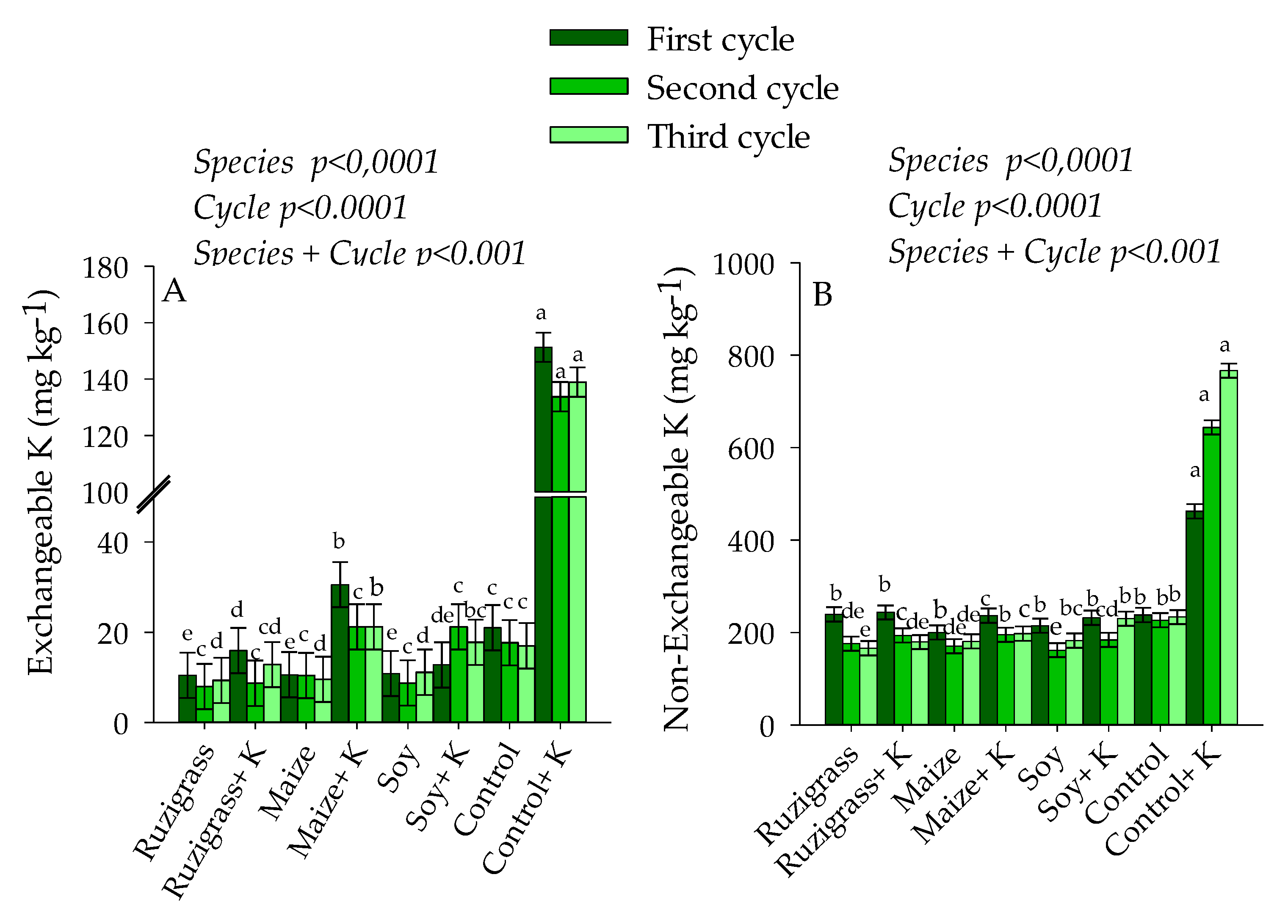
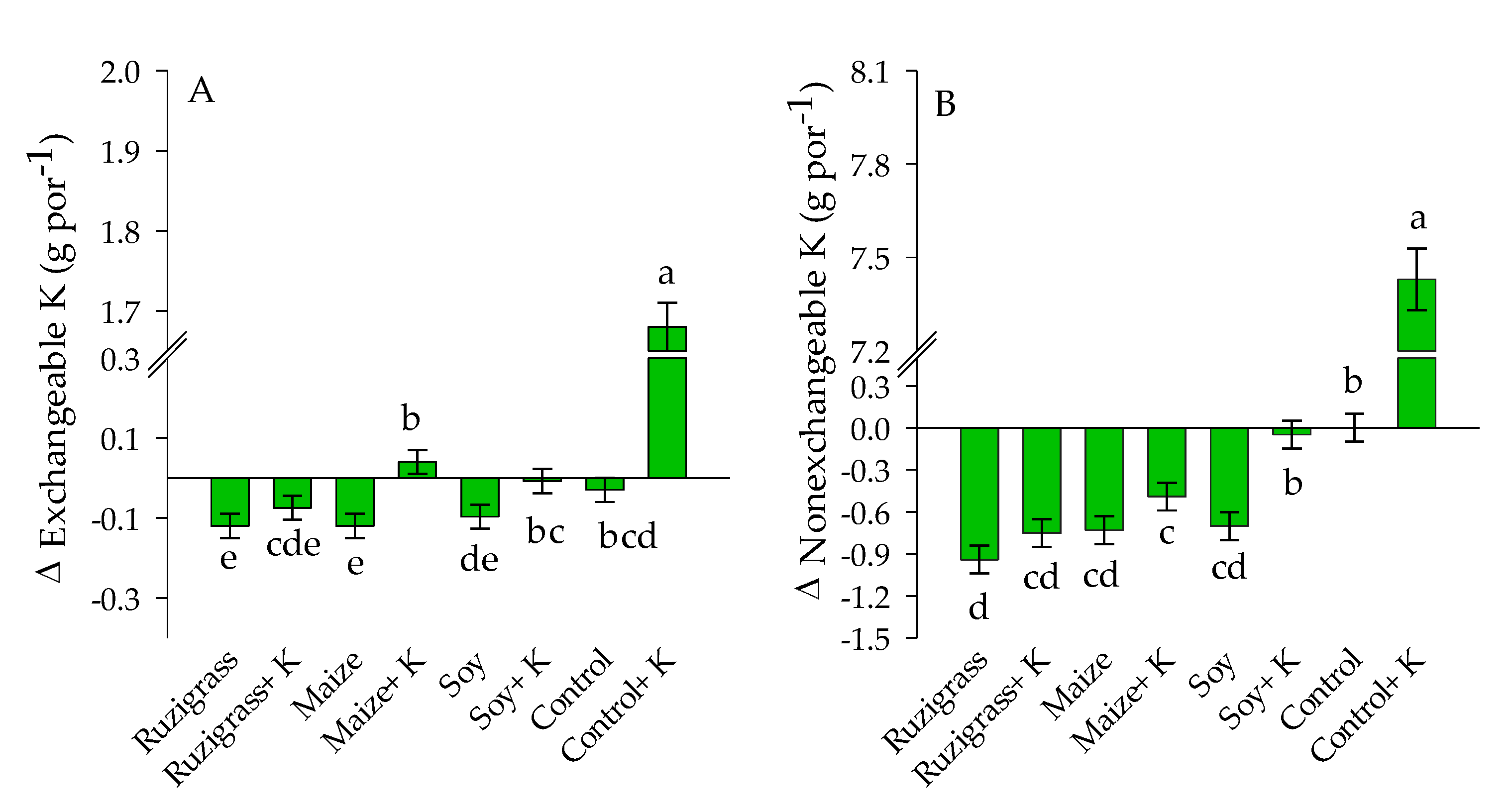
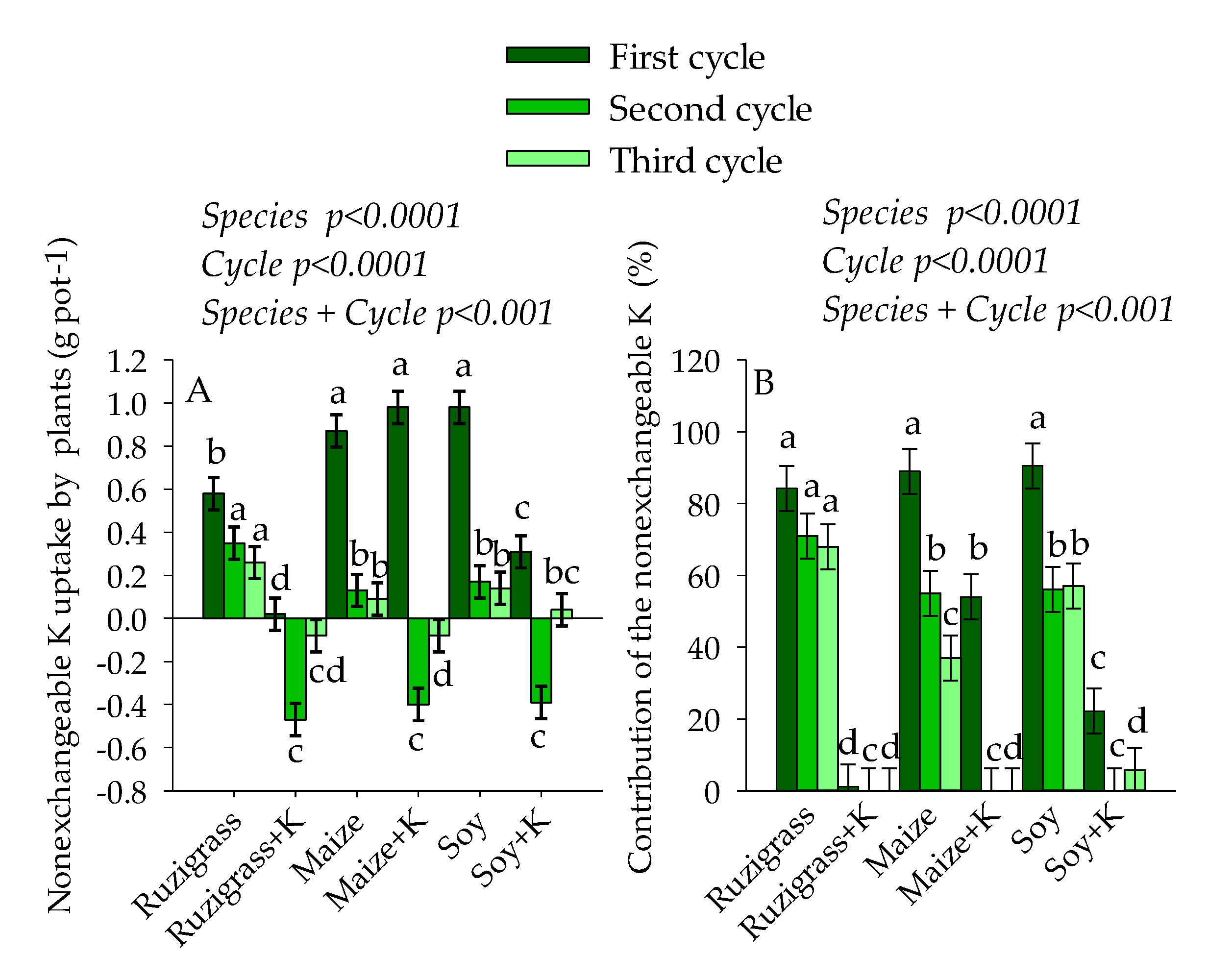
3. Results
3.1. Potassium in Soil and Plants
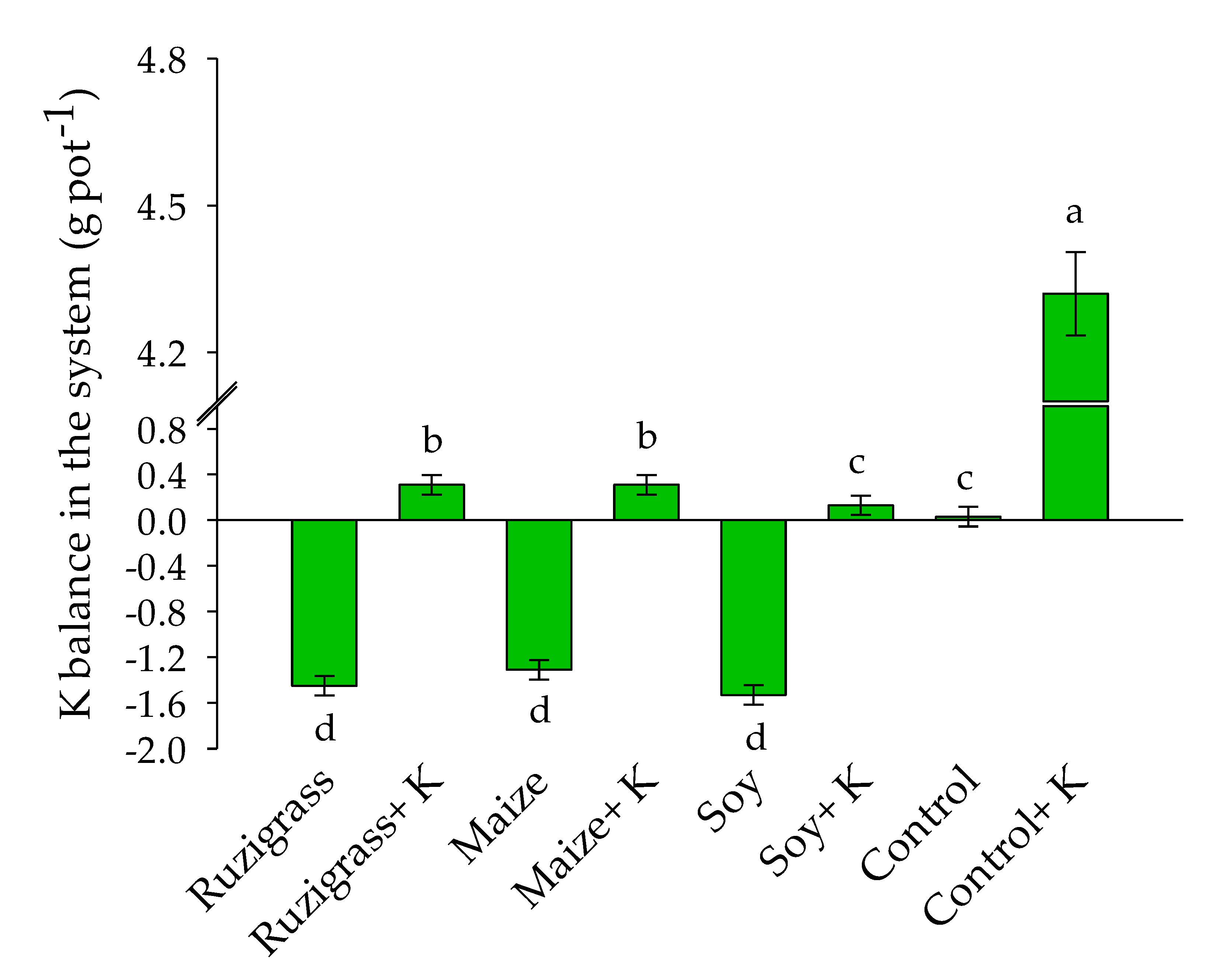
3.2. Potassium Balance in the System
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Inc.: London, UK, 2012; ISBN 9780123849052. [Google Scholar]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Rosolem, C.A.; Steiner, F. Effects of soil texture and rates of K input on potassium balance in tropical soil. Eur. J. Soil Sci. 2017, 68, 658–666. [Google Scholar] [CrossRef]
- Kaminski, J.; Brunetto, G.; Moterle, D.F.; Rheinheimer, D.S. Depletion of soil potassium as affected by successive cultivation. Rev. Bras. De Cienc. Do Solo 2007, 31, 1003–1010. [Google Scholar] [CrossRef] [Green Version]
- Bortoluzzi, E.C.; Santos, D.R.d.; Kaminski, J.; Gatiboni, L.C.; Tessier, D. Alterações na mineralogia de um argissolo do rio grande do sul submetido à fertilização potássica. Rev. Bras. De Ciência Do Solo 2005, 29, 327–335. [Google Scholar] [CrossRef]
- Singh, K.; Bansal, S.K. Long-term potassium contribution of potassium forms of udic haplustept and sustainability of growing crops. Commun. Soil Sci. Plant Anal. 2013, 44, 1282–1292. [Google Scholar] [CrossRef]
- Barré, P.; Velde, B.; Catel, N.; Abbadie, L. Soil-plant potassium transfer: Impact of plant activity on clay minerals as seen from X-Ray diffraction. Plant Soil 2007, 292, 137–146. [Google Scholar] [CrossRef]
- Bilias, F.; Barbayiannis, N. Evaluation of sodium tetraphenylboron (NaBPh 4 ) as a soil test of potassium availability. Arch. Agron. Soil Sci. 2017, 63, 468–476. [Google Scholar] [CrossRef]
- Kitagawa, Y.; Yanai, J.; Nakao, A. Evaluation of nonexchangeable potassium content of agricultural soils in Japan by the boiling HNO3extraction method in comparison with exchangeable potassium. Soil Sci. Plant Nutr. 2018, 64, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Barbagelata, P.A.; Mallarino, A.P. Field correlation of potassium soil test methods based on dried and field-moist soil samples for corn and soybean. Soil Sci. Soc. Am. J. 2013, 77, 318–327. [Google Scholar] [CrossRef]
- Antonangelo, J.A.; Firmano, R.F.; Alleoni, L.R.F.; Oliveira, A.; Zhang, H. Soybean production under continuous potassium fertilization in a long-term no-till oxisol. Agron. J. 2019, 111, 2462–2471. [Google Scholar] [CrossRef]
- Raij, V.B.; Quaggio, J.A.; da Silva, N.M. Extraction of phosphorus, potassium, calcium, and magnesium from soils by an ion-exchange resin procedure. Commun. Soil Sci. Plant Anal. 1986, 17, 547–566. [Google Scholar] [CrossRef]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Kerbauy, G.B. Fisiologia Vegetal, 2nd ed.; Guanabara Koogan São Paulo: São Paulo, Brazil, 2008. [Google Scholar]
- Barré, P.; Berger, G.; Velde, B. how element translocation by plants may stabilize illitic clays in the surface of temperate soils. Geoderma 2009, 151, 22–30. [Google Scholar] [CrossRef]
- Volf, M.R.; Guimarães, T.M.; Scudeletti, D.; Cruz, I.V.; Rosolem, C.A. Potassium dynamics in ruzigrass rhizosphere. Rev. Bras. De Ciência Do Solo 2018, 42, 170370. [Google Scholar] [CrossRef] [Green Version]
- Hinsinger, P.; Jaillard, B. Root-induced release of interlayer potassium and vermiculitization of phlogopite as related to potassium depletion in the rhizosphere of ryegrass. J. Soil Sci. 1993, 44, 525–534. [Google Scholar] [CrossRef]
- Benites, V.M.; Carvalho, M.C.S.; Resende, A.V.; Polidoro, J.C.; Bernardi, A.C.C.; Oliveira, F.A. Potássium, Calcium and Magnesium, 2nd ed.; Prochnon, L.I., Casarin, V., Stipp, S.R., Eds.; IPNI: Piracicaba, Brazil, 2010; ISBN 9788598519050. [Google Scholar]
- Darunsontaya, T.; Suddhiprakarn, A.; Kheoruenromne, I.; Gilkes, R.J. The kinetics of potassium release to sodium tetraphenylboron solution from the clay fraction of highly weathered soils. Appl. Clay Sci. 2010, 50, 376–385. [Google Scholar] [CrossRef]
- Oliveira Junior, A.; Castro, C.; Oliveira, F.A.; Foloni, J.S.S. Marcha de absorção e acúmulo de macronutrientes em soja com tipo de crescimento indeterminado. Comissão De Nutr. Veg. Fertil. E Biol. Dos Solos 2014, 1, 133–136. [Google Scholar]
- Castilhos, R.M.; Meurer, E.J.; Kämpf, N.; Pinto, L.F.S. Mineralogia e fontes de potássio em solos no rio grande do sul cultivados com arroz irrigado. Rev. Bras. De Ciência Do Solo 2002, 26, 579–587. [Google Scholar] [CrossRef]
- Melo, V.F.; Maschio, P.A.; Ribeiro, A.N.; Lima, V.C. Importância das espécies minerais no potássio total da fração argila de solos do triângulo mineiro. Rev. Bras. De Ciências Do Solo 2003, 27, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Mulvaney, R.L.; Ellsworth, T.R. The potassium paradox: Implications for soil fertility, crop production and human health. Renew. Agric. Food Syst. 2014, 29, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Essel, B.; Abaidoo, R.C.; Opoku, A.; Ewusi-Mensah, N. Economically optimal rate for nutrient application to maize in the semi-deciduous forest zone of Ghana. J. Soil Sci. Plant Nutr. 2020, 20, 1703–1713. [Google Scholar] [CrossRef]
- Villa, M.R.; Fernandes, L.A.; Faquin, V. Formas de potássio em solos de várzea e sua disponibilidade para o feijoeiro. Rev. Bras. De Cienc. Do Solo 2004, 28, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Hosseinpur, A.R.; Motaghian, H.R. Application of kinetic models in describing soil potassium release characteristics and their correlations with potassium extracted by chemical methods. Pedosphere 2013, 23, 482–492. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Calonego, J.C.; Foloni, J.S.S. Potassium leaching from green cover crop residues as affected by rainfall amount. Rev. Bras. De Ciência Do Solo 2003, 27, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Rosolem, C.A.; Bessa, A.M.; Marques, H.F. Potassium dynamics in soil and soybean potassic nutrition. Pesqui. Agropecu. Bras. 1993, 28, 1045–1054. [Google Scholar]
- Garcia, R.A.; Crusciol, C.A.C.; Calonego, J.C.; Rosolem, C.A. Potassium cycling in a corn-brachiaria cropping system. European J. Agron. 2008, 28, 579–585. [Google Scholar] [CrossRef]
- Volf, M.R. Rotation Soybean-Forage and the Contribution of Non- Exchangeable K in Plant Nutrition in a Cerrado Soil. Ph.D. Thesis, São Paulo State University, Botucatu, São Paulo, Brazil, 2019. [Google Scholar]
- Walkley, A.J.; Black, I.A. Estimation of soil organic carbon by the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Mateus, G.P.; Godoy, L.J.G.; Feltran, J.C.; Brancalião, S.R. Morfologia radicular e suprimento de potássio às raízes de milheto de acordo com a disponibilidade de água e potássio. Rev. Bras. De Ciência Do Solo 2003, 27, 875–884. [Google Scholar] [CrossRef] [Green Version]
- Galdos, M.V.; Brown, E.; Rosolem, C.A.; Pires, L.F.; Hallett, P.D.; Mooney, S.J. Brachiaria species influence nitrate transport in soil by modifying soil structure with their root system. Sci. Rep. 2020, 10, 5072. [Google Scholar] [CrossRef] [Green Version]
- Rosolem, C.A.; Vicentini, J.P.T.M.M.; Steiner, F. Potassium supply as affected by residual potassium fertilization in a cerrado oxisol. Rev. Bras. De Cienc. Do Solo 2012, 36, 1507–1515. [Google Scholar] [CrossRef] [Green Version]
- Andreotti, M.; de Souza, E.C.A.; Crusciol, C.A.C. Componentes morfológicos e produção de matéria seca de milho em função da aplicação de calcário e zinco. Sci. Agric. 2001, 58, 321–327. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Vicentini, J.P.T.M.M.; Steiner, F. Suprimento de potássio em função da adubação potássica residual em um latossolo vermelho do cerrado. Rev. Bras. De Ciência Do Solo 2012, 36, 1507–1515. [Google Scholar] [CrossRef] [Green Version]
- Bender, R.R.; Haegele, J.W.; Below, F.E. Nutrient uptake, partitioning, and remobilization in modern soybean varieties. Agron. J. 2015, 107, 563–573. [Google Scholar] [CrossRef] [Green Version]
- Vilela, E.F.; Büll, L.T. Avaliação do crescimento de plantas de milho em função de doses de potássio e estresse hídrico. Revista Bras. De Ciência Do Solo 2014, 23, 281–289. [Google Scholar] [CrossRef]
- Malavolta, E.; Vitti, G.C.; Oliveira, S.A. Avaliaçãodo Estado Nutricional Das Plantas: Princípios e Aplicações; IPNI: São Paulo, Brazil, 1997. [Google Scholar]
- Knudsen, D.; Peterson, G.A.; Pratt, P.F. Lithium, Sodium, and Potassium; American Society of Agronomy: Madison, NJ, USA, 1982. [Google Scholar]
- Ferreira, D.F. SISVAR: A computer statistical analysis system. Sisvar: Um sistema computacional de análise estatística. Ciênc. Agrotec 2011, 35, 1039–1042. [Google Scholar] [CrossRef] [Green Version]
- Moterle, D.F.; Kaminski, J.; dos Santos Rheinheimer, D.; Caner, L.; Bortoluzzi, E.C. Impact of potassium fertilization and potassium uptake by plants on soil clay mineral assemblage in South Brazil. Plant Soil 2016, 406, 157–172. [Google Scholar] [CrossRef]
- Li, T.; Wang, H.; Wang, J.; Zhou, Z.; Zhou, J. Exploring the potential of phyllosilicate minerals as potassium fertilizers using sodium tetraphenylboron and intensive cropping with perennial ryegrass. Sci. Rep. 2015, 5, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Wang, H.; Chen, X.; Zhou, J. Soil reserves of potassium: Release and availability to lolium perenne in relation to clay minerals in six cropland soils from Eastern China. Land Degrad. Dev. 2017, 28, 1696–1703. [Google Scholar] [CrossRef]
- Islam, S.; Timsina, J.; Salim, M.; Majumdar, K.; Gathala, M. Potassium supplying capacity of diverse soils and K-use efficiency of maize in South Asia. Agronomy 2018, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Firmano, R.F.; de Oliveira, A.; de Castro, C.; Alleoni, L.R.F. Potassium rates on the cationic balance of an oxisol and soybean nutritional status after 8 years of K deprivation. Exp. Agric. 2019, 1–19. [Google Scholar] [CrossRef]
- Raij, V.B.; Quaggio, J.A. Disponibilidade de potássio em solos para capim-braquiária cultivado em vasos. Bragantia 1984, 43, 531–539. [Google Scholar] [CrossRef]
- Hinsinger, P.; Elsass, F.; Jaillard, B.; Robert, M. Root-induced irreversible transformation of a trioctahedral mica in the rhizosphere of rape. J. Soil Sci. 1993, 44, 535–545. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Singh, V.; Farooq, M.; Singh, R.; Rahman, M. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2018, 1–14. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Kaur, T.; Yadav, N.; Halder, S.K.; Yadav, A.N.; Sachan, S.G.; Saxena, A.K. Potassium Solubilizing and Mobilizing Microbes: Biodiversity, Mechanisms of Solubilization, and Biotechnological Implication for Alleviations of Abiotic Stress; Elsevier Inc.: Amsterdam, The Netherlands, 2020; ISBN 9780128205266. [Google Scholar]
- Sun, Z. Interpretation of action potential generation mechanism in cardiomyocytes by potassium channel “origami windmill” model. J. US-China Med. Sci. 2020, 17. [Google Scholar] [CrossRef]
- Almeida, D.S.; Delai, L.B.; Sawaya, A.C.H.F.; Rosolem, C.A. Exudation of organic acid anions by tropical grasses in response to low phosphorus availability. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- White, P.J. Improving potassium acquisition and utilisation by crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 305–316. [Google Scholar] [CrossRef]
- Sousa, D.M.G.; Lobato, E. Correção Do Solo e Adubação Da Cultura Da Soja; Embrapa Cerrados: Planaltina, Brazil, 1996. [Google Scholar]
- Rosolem, C.A.; Neto, L.O.; Costa, V.E.; da Silva Grassmann, C. Ruzigrass root persistence and soybean root growth. Plant Soil 2019, 442, 333–341. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Damodar Reddy, D. Potassium balance and release kinetics under continuous rice—Wheat cropping system in vertisol. Field Crop. Res. 2002, 77, 81–91. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Dos Santos, F.P.; Foloni, J.S.S.; Calonego, J.C. Soil potassium as affected by fertilization over the millet straw and simulated rain|potássio no solo em conseqüência da adubação sobre a palha de milheto e chuva simulada. Pesqui. Agropecu. Bras. 2006, 41, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Harrell, D.L.; Bell, P.F. Potassium buffering characteristics of three soils low in exchangeable potassium. Soil Sci. Soc. Am. J. 2004, 68, 654. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properts | Value | Units |
|---|---|---|
| Clay | 275 | g·kg−1 |
| Silt | 175 | g·kg−1 |
| Sand | 550 | g·kg−1 |
| Initial pH 1 (CaCl2) | 3.8 | – |
| Soil organic carbon 5 | 7.0 | g·kg−1 |
| Available phosphorus (P resin) | 7.2 | mg·kg−1 |
| Calcium (Ca2+ resin) | 120 | mg·kg−1 |
| Mg (Mg2+ resin) | 30 | mg·kg−1 |
| Potassium 2 (K+ resin) | 72 | mg·kg−1 |
| Non-Exchangeable K 4 | 230 | mg·kg−1 |
| NAE K 3 (HNO3) | 302 | mg·kg−1 |
| Cation exchange capacity (CEC pH 7.0) | 8.3 | cmolc kg−1 |
| Treatments | Rates (mg K kg−1 Soil) * | ||
|---|---|---|---|
| First Crop | Second Crop | Third Crop | |
| Ruzigrass + K | 70 | 115 | 70 |
| Ruzigrass | 0 | 0 | 0 |
| Soy + K | 70 | 130 | 70 |
| Soy | 0 | 0 | 0 |
| Maize+ K | 70 | 200 | 140 |
| Maize | 0 | 0 | 0 |
| Control Soil + K | 140 | 140 | 140 |
| Control Soil | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volf, M.R.; Crusciol, C.A.C.; de Azevedo, A.C.; Thompson, M.L.; Kovar, J.L.; Rosolem, C.A. Potassium Bioavailability in a Tropical Kaolinitic Soil. Agronomy 2021, 11, 2016. https://doi.org/10.3390/agronomy11102016
Volf MR, Crusciol CAC, de Azevedo AC, Thompson ML, Kovar JL, Rosolem CA. Potassium Bioavailability in a Tropical Kaolinitic Soil. Agronomy. 2021; 11(10):2016. https://doi.org/10.3390/agronomy11102016
Chicago/Turabian StyleVolf, Marcelo Raphael, Carlos Alexandre Costa Crusciol, Antônio Carlos de Azevedo, Michael L. Thompson, John L. Kovar, and Ciro Antonio Rosolem. 2021. "Potassium Bioavailability in a Tropical Kaolinitic Soil" Agronomy 11, no. 10: 2016. https://doi.org/10.3390/agronomy11102016