
Effects of Phosphorus Supply on the Leaf Photosynthesis, and Biomass and Phosphorus Accumulation and Partitioning of Canola (Brassica napus L.) in Saline Environment

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Site, Soil Properties and Experimental Set
2.2. Assessment of Seed Yield Performance and Seed Oil Content
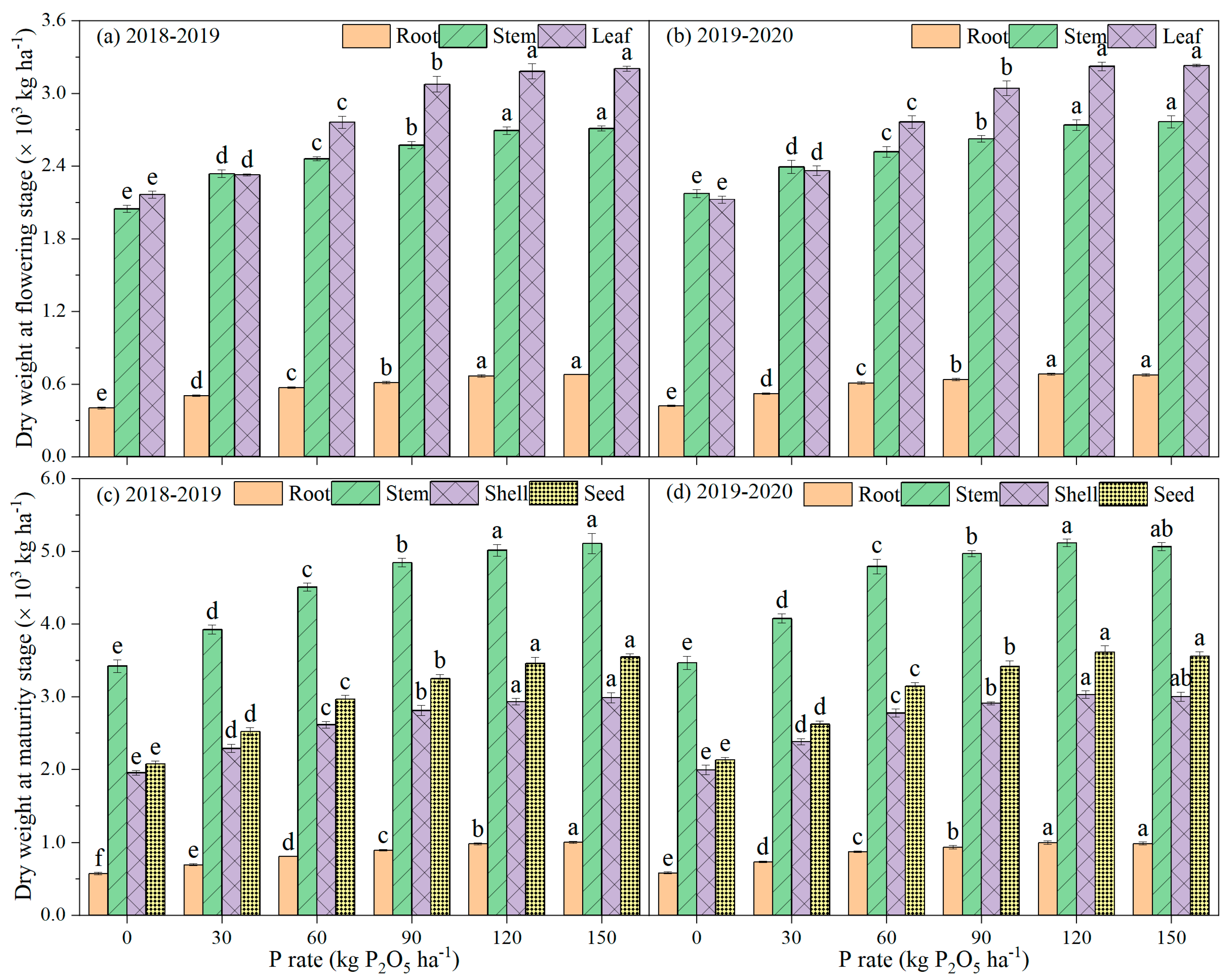
2.3. Assessment of Biomass and P Accumulation
2.4. Assessment of Biomass and P Partitioning
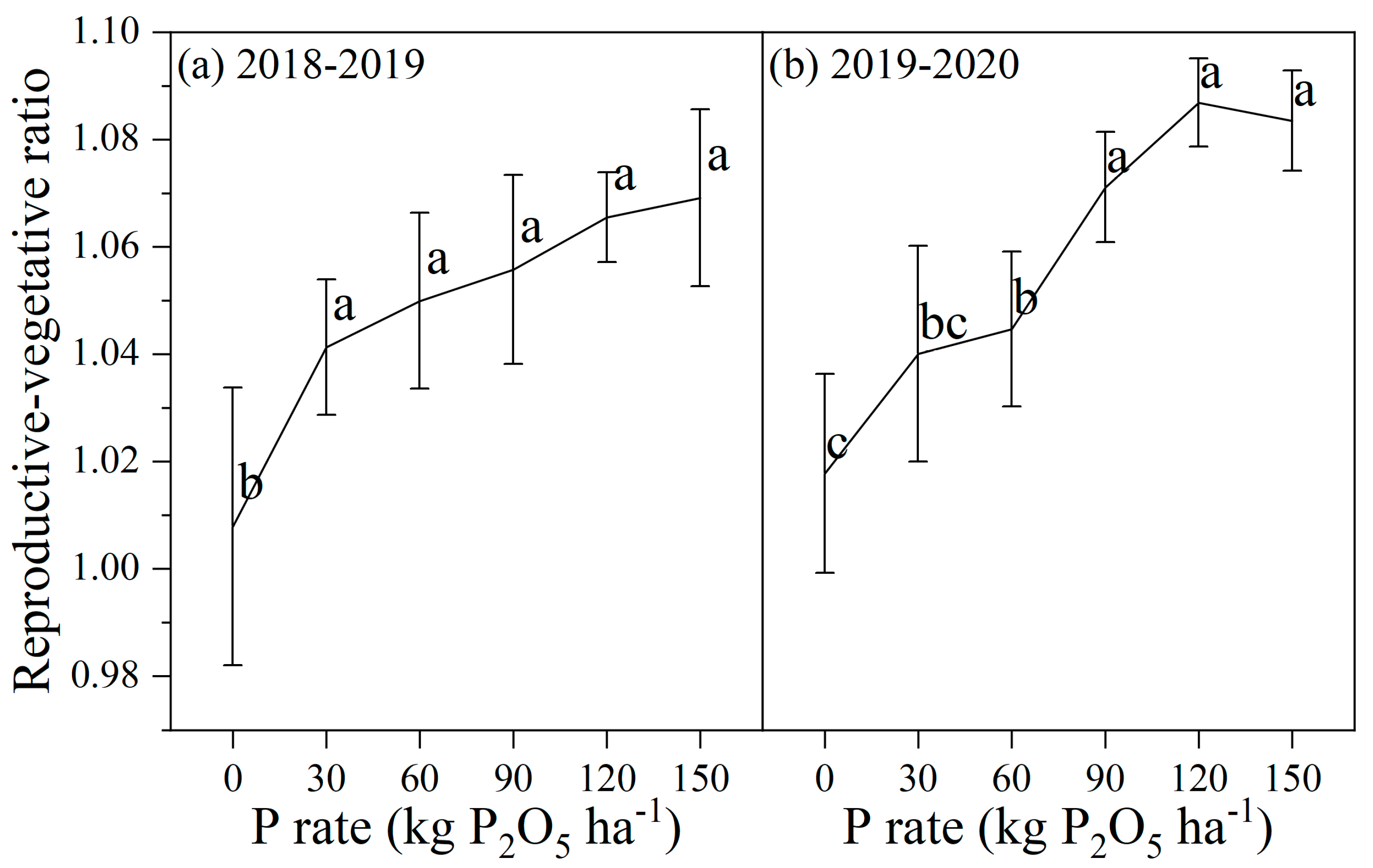
2.5. Assessment of the Reproductive-Vegetative Ratio
2.6. Assessment of Leaf Area Index and Photosynthetic Rate
2.7. Statistical Analysis
3. Results
3.1. Seed and Oil Yield Performance
3.2. Biomass and P Accumulation
3.3. Biomass and P Partitioning, and the Reproductive-Vegetative Ratio
3.4. Leaf Area Index and Leaf Photosynthesis
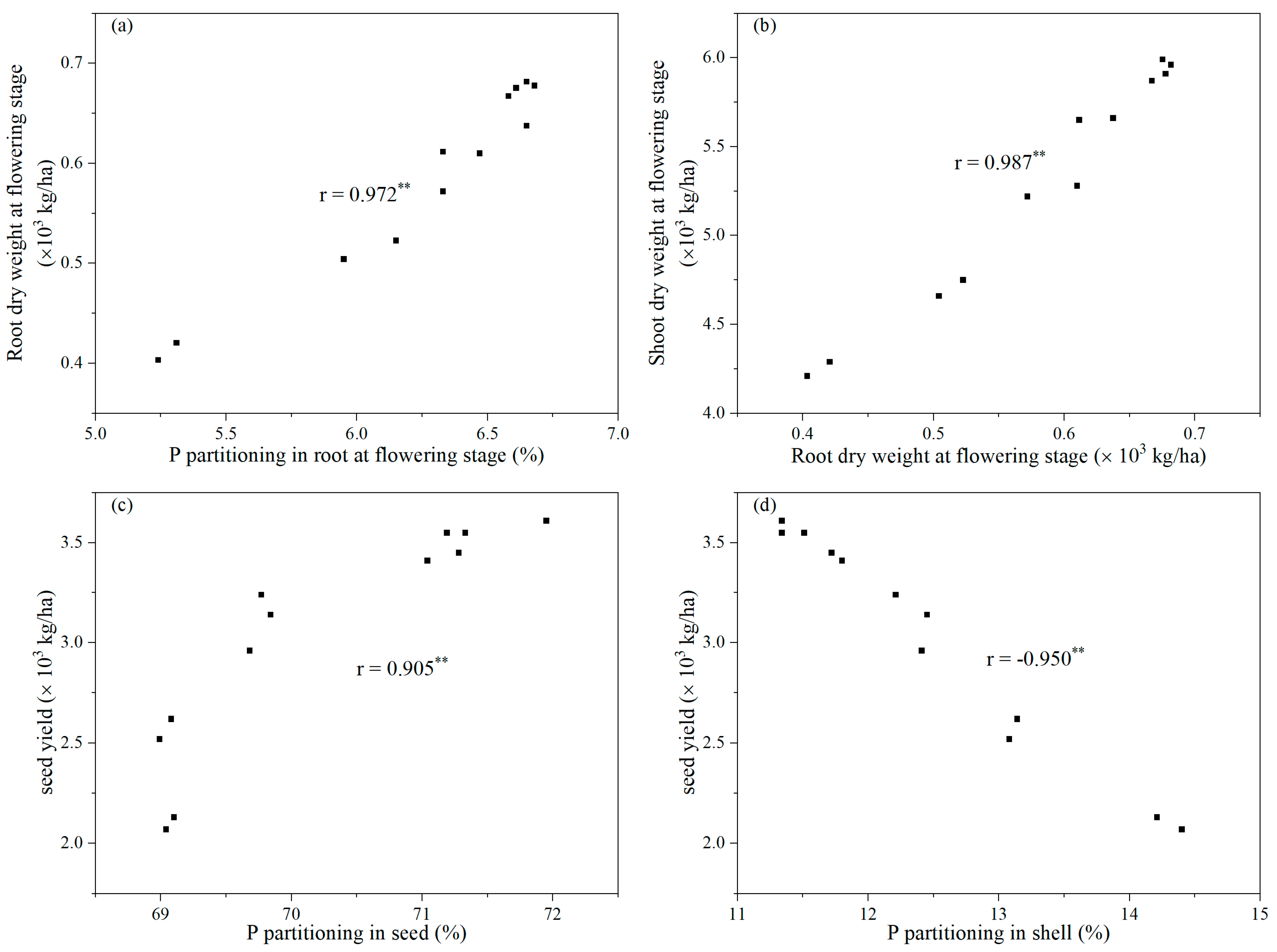
3.5. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, Q.; Hua, W.; Yin, Y.; Zhang, X.; Liu, L.; Shi, J.; Zhao, Y.; Qin, L.; Chen, C.; Wang, H. Rapeseed research and production in China. Crop J. 2017, 5, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Gunstone, F.D. Production and consumption of rapeseed oil on a global scale. Eur. J. Lipid Sci. Technol. 2001, 103, 447–449. [Google Scholar] [CrossRef]
- Bonjean, A.P.; Dequidt, C.; Sang, T. Rapeseed in China. OCL 2016, 23, D605. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Prasad, M.N.V.; Ozturk, M.; Fujita, M. Potential Use of Halophytes to Remediate Saline Soils. Biomed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Mittal, S.; Kumari, N.; Sharma, V. Differential response of salt stress on Brassica juncea: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes. Plant Physiol. Biochem. 2012, 54, 17–26. [Google Scholar] [CrossRef]
- Tanveer, M.; Shah, A.N. An insight into salt stress tolerance mechanisms of Chenopodium album. Environ. Sci. Pollut. Res. 2017, 24, 16531–16535. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Bybordi, A. Effect of ammonium/nitrate nitrogen ratio on photosynthesis, respiration and some vegetative traits of canola grown under salinity stress. J. Food Agric. Environ. 2012, 10, 372–375. [Google Scholar]
- Gao, L.; Liu, M.; Wang, M.; Shen, Q.; Guo, S. Enhanced Salt Tolerance under Nitrate Nutrition is Associated with Apoplast Na+ Content in Canola (Brassica napus L.) and Rice (Oryza sativa L.) Plants. Plant Cell Physiol 2016, 57, 2323–2333. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.L.; Niu, L.; Wei, J.; Chen, X.Y.; Chen, Y.L. Phosphorus Limitation Improved Salt Tolerance in Maize Through Tissue Mass Density Increase, Osmolytes Accumulation, and Na+ Uptake Inhibition. Front. Plant Sci. 2019, 10, 856. [Google Scholar] [CrossRef] [Green Version]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [Green Version]
- Zribi, O.T.; Hessini, K.; Trabelsi, N.; Zribi, F.; Hamdi, A.; Ksouri, R.; Abdelly, C. Aeluropus littoralis maintains adequate gas exchange, pigment composition and phenolic contents under combined effects of salinity and phosphorus deficiency. Aust. J. Bot. 2017, 65, 453–462. [Google Scholar] [CrossRef]
- Sabir, M.; Sultan, T.; Yaseen, M.; Ahmad, R.; Tarar, Z.H. Comparative Effect of Different Phosphatic Fertilizers on Growth of Rapeseed (Brassica napus L.) in Alkaline Calcareous Saline-Sodic Soils. Commun. Soil Sci. Plant Anal. 2021. [Google Scholar] [CrossRef]
- Sharif, M.; Sarir, M.S. Biological and chemical transformation of phosphorus in some important soil series of NWFP. Sarhad J. Agric. 2000, 16, 587–592. [Google Scholar]
- Zhang, K.; Shi, Y.; Cui, X.; Yue, P.; Li, K.; Liu, X.; Tripathi, B.M.; Chu, H. Salinity Is a Key Determinant for Soil Microbial Communities in a Desert Ecosystem. Msystems 2019, 4, e00225-18. [Google Scholar] [CrossRef] [Green Version]
- Li, J.Z.; Xie, Y.; Dai, A.Y.; Liu, L.F.; Li, Z.C. Root and shoot traits responses to phosphorus deficiency and QTL analysis at seedling stage using introgression lines of rice. J. Genet. Genom. 2009, 36, 173–183. [Google Scholar] [CrossRef]
- Liu, H.; Hu, C.; Hu, X.; Nie, Z.; Sun, X.; Tan, Q.; Hu, H. Interaction of molybdenum and phosphorus supply on uptake and translocation of phosphorus and molybdenum by Brassica napus. J. Plant Nutr. 2010, 33, 1751–1760. [Google Scholar] [CrossRef] [Green Version]
- Yaryura, P.; Cordon, G.; Leon, M.; Kerber, N.; Pucheu, N.; Rubio, G.; Garcia, A.; Lagorio, M.G. Effect of Phosphorus Deficiency on Reflectance and Chlorophyll Fluorescence of Cotyledons of Oilseed Rape (Brassica napus L.). J. Agron. Crop. Sci. 2009, 195, 186–196. [Google Scholar] [CrossRef]
- Ding, G.; Zhao, Z.; Liao, Y.; Hu, Y.; Shi, L.; Long, Y.; Xu, F. Quantitative trait loci for seed yield and yield-related traits, and their responses to reduced phosphorus supply in Brassica napus. Ann. Bot. 2012, 109, 747–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouras, H.; Bouaziz, A.; Bouazzama, B.; Hirich, A.; Choukr-Allah, R. How Phosphorus Fertilization Alleviates the Effect of Salinity on Sugar Beet (Beta vulgaris L.) Productivity and Quality. Agronomy 2021, 11, 1491. [Google Scholar] [CrossRef]
- Zribi, O.T.; Slama, I.; Trabelsi, N.; Hamdi, A.; Smaoui, A.; Abdelly, C. Combined effects of salinity and phosphorus availability on growth, gas exchange, and nutrient status of Catapodium rigidum. Arid Land Res. Manag. 2018, 32, 277–290. [Google Scholar] [CrossRef]
- Lee, H.; Zaman, R.; Lee, B.-R.; Kim, T.-H. Effects of Nitrogen Level on Nitrogen Partitioning and Harvest Index in Brassica napus L. J. Korean Soc. Grassl. Sci. 2018, 38, 140–144. [Google Scholar] [CrossRef]
- Zuo, Q.; Liu, J.; Shan, J.; Zhou, J.; Wang, L.; Yang, G.; Leng, S.; Liu, H. Carbon and Nitrogen Assimilation and Partitioning in Canola (Brassica napus L.) In Saline Environment. Commun. Soil Sci. Plant Anal. 2019, 50, 1700–1709. [Google Scholar] [CrossRef]
- Zhao, Z.; Wang, S.; White, P.J.; Wang, Y.; Shi, L.; Xu, F. Boron and Phosphorus Act Synergistically to Modulate Absorption and Distribution of Phosphorus and Growth of Brassica napus. J. Agric. Food Chem. 2020, 68, 7830–7838. [Google Scholar] [CrossRef]
- Liu, L.; Long, X.; Shao, H.; Liu, Z.-P.; Tao, Y.; Zhou, Q.; Zong, J. Ameliorants improve saline-alkaline soils on a large scale in northern Jiangsu Province, China. Ecol. Eng. 2015, 81, 328–334. [Google Scholar] [CrossRef]
- Bybordi, A. The Influence of Salt Stress on Seed Germination, Growth and Yield of Canola Cultivars. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2010, 38, 128–133. [Google Scholar]
- Yousaf, M.; Li, J.F.; Lu, J.W.; Ren, T.; Cong, R.H.; Fahad, S.; Li, X.K. Effects of fertilization on crop production and nutrient-supplying capacity under rice-oilseed rape rotation system. Sci. Rep. 2017, 7, 1270. [Google Scholar] [CrossRef] [PubMed]
- Brennan, R.F.; Bolland, M.D.A. Effect of fertiliser phosphorus and nitrogen on the concentrations of oil and protein in grain and the grain yield of canola (Brassica napus L.) grown in south-western Australia. Aust. J. Exp. Agric. 2007, 47, 984–991. [Google Scholar] [CrossRef]
- Lickfett, T.; Matthaus, B.; Velasco, L.; Mollers, C. Seed yield, oil and phytate concentration in the seeds of two oilseed rape cultivars as affected by different phosphorus supply. Eur. J. Agron. 1999, 11, 293–299. [Google Scholar] [CrossRef]
- Bybordi, A.; Ebrahimian, E. Growth, Yield and Quality Components of Canola Fertilized with Urea and Zeolite. Commun. Soil Sci. Plant Anal. 2013, 44, 2896–2915. [Google Scholar] [CrossRef]
- Wahid, M.A.; Cheema, M.A.; Saleem, M.F.; Nadeem, M.; Sattar, A.; Zaman, M. Canola growth and phosphorus amendments. i. yield and quality response of canola to different phosphorus amendments. Pak. J. Agric. Sci. 2014, 51, 847–854. [Google Scholar]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root structure and functioning for efficient acquisition of phosphorus: Matching morphological and physiological traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; George, T.S.; Dupuy, L.X.; Karley, A.J.; Valentine, T. Root traits for infertile soils. Front. Plant Sci. 2013, 4, 193. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Ding, G.-D.; White, P.J.; Wang, X.-H.; Jin, K.-M.; Xu, F.-S.; Shi, L. Mapping and cloning of quantitative trait loci for phosphorus efficiency in crops: Opportunities and challenges. Plant Soil 2019, 439, 91–112. [Google Scholar] [CrossRef]
- Nourgholipour, F.; Hosseini, H.M.; Tehrani, M.M.; Zadeh, B.M.; Moshiri, F. Phosphorus Efficiency of Winter Canola Cultivars and Rhizosphere Properties in Rhizobox Technique. Commun. Soil Sci. Plant Anal. 2019, 50, 35–48. [Google Scholar] [CrossRef]
- Elanchezhian, R.; Krishnapriya, V.; Pandey, R.; Rao, A.S.; Abrol, Y.P. Physiological and molecular approaches for improving phosphorus uptake efficiency of crops. Curr. Sci. 2015, 108, 1271–1279. [Google Scholar]
- Lyu, Y.; Tang, H.; Li, H.; Zhang, F.; Rengel, Z.; Whalley, W.R.; Shen, J. Major Crop Species Show Differential Balance between Root Morphological and Physiological Responses to Variable Phosphorus Supply. Front. Plant Sci. 2016, 7, 1939. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Jin, K.; Ding, G.; Wang, C.; Cai, H.; Wang, S.; White, P.J.; Xu, F.; Shi, L. The impact of different morphological and biochemical root traits on phosphorus acquisition and seed yield of Brassica napus. Field Crop. Res. 2020, 258, 107960. [Google Scholar] [CrossRef]
- Heyneke, E.; Fernie, A.R. Metabolic regulation of photosynthesis. Biochem. Soc. Trans. 2018, 46, 321–328. [Google Scholar] [CrossRef]
- Zangani, E.; Afsahi, K.; Shekari, F.; Mac Sweeney, E.; Mastinu, A. Nitrogen and Phosphorus Addition to Soil Improves Seed Yield, Foliar Stomatal Conductance, and the Photosynthetic Response of Rapeseed (Brassica napus L.). Agriculture 2021, 11, 483. [Google Scholar] [CrossRef]
- Chen, B.; Wang, Q.; Tang, X.; Ye, Z.; Stiles, S.; Feng, G. Growth and sink-source relationship of cotton (Gossypium hirsutum L.) under mulched drip irrigation in response to phosphorus fertilization and cultivar. J. Plant Nutr. 2021. [Google Scholar] [CrossRef]
- Wang, C.; Hai, J.; Tian, J.; Yang, J.; Zhao, X. Influence of Silique and Leaf Photosynthesis on Yield and Quality of Seed of Oilseed Rape (Brassica napus L.) after Flowering. Xibei Zhiwu Xuebao 2014, 34, 1620–1626. [Google Scholar] [CrossRef]
- Mahmood, M.; Tian, Y.; Ma, Q.; Ahmed, W.; Mehmood, S.; Hui, X.; Wang, Z. Changes in Phosphorus Fractions and Its Availability Status in Relation to Long Term P Fertilization in Loess Plateau of China. Agronomy 2020, 10, 1818. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Soil Properties | Value | |
|---|---|---|---|
| 2018 | 2019 | ||
| Soil nutrients content | Organic matter (g/kg) | 15.2 | 16.3 |
| Total nitrogen (g/kg) | 0.81 | 0.83 | |
| Alkali-hydrolysable nitrogen (mg/kg) | 61.7 | 64.2 | |
| Available phosphorus (mg/kg) | 10.8 | 11.6 | |
| Available potassium (mg/kg) | 182.5 | 179.6 | |
| Soil salt ion concentration | Na+ (g/kg) | 0.523 | 0.507 |
| K+ (g/kg) | 0.063 | 0.061 | |
| Ca2+ (g/kg) | 0.276 | 0.265 | |
| Mg2+ (g/kg) | 0.061 | 0.059 | |
| Cl− (g/kg) | 1.112 | 1.041 | |
| HCO3− (g/kg) | 0.345 | 0.346 | |
| SO42− (g/kg) | 0.312 | 0.318 | |
| Total salt ion (g/kg) | 2.692 | 2.597 | |
| Soil pH | pH | 8.13 | 8.06 |
| Year | P Rate (kg P2O5 ha−1) | Number of Primary Branches per Plant | Seed Yield (×103 kg ha−1) | Seed Oil Concentration (%) | Oil Yield (kg ha−1) | Number of Pods per Plant | Number of Seeds per Pod | 1000-Seed Weight |
|---|---|---|---|---|---|---|---|---|
| 2018–2019 | 0 | 6.1 e | 1.94 e | 43.73 c | 847.2 e | 76.8 e | 15.9 d | 3.77 a |
| 30 | 6.8 d | 2.42 d | 44.35 bc | 1075.3 d | 88.5 d | 16.8 c | 3.77 a | |
| 60 | 7.4 c | 2.83 c | 45.26 ab | 1282.3 c | 101.4 c | 17.0 bc | 3.82 a | |
| 90 | 8.5 b | 3.09 b | 46.25 a | 1430.5 b | 111.1 b | 17.2 ab | 3.77 a | |
| 120 | 9.2 a | 3.33 a | 46.44 a | 1545.1 a | 116.1 a | 17.2 ab | 3.84 a | |
| 150 | 9.0 a | 3.35 a | 46.49 a | 1558.7 a | 118.6 a | 17.5 a | 3.80 a | |
| 2019–2020 | 0 | 5.9 e | 2.04 e | 44.02 c | 898.9 e | 77.7 e | 16.3 c | 3.74 a |
| 30 | 6.9 d | 2.51 d | 44.53 bc | 1116.8 d | 91.2 d | 16.8 b | 3.80 a | |
| 60 | 7.5 c | 2.91 c | 45.86 ab | 1335.4 c | 106.4 c | 17.2 ab | 3.82 a | |
| 90 | 8.4 b | 3.22 b | 46.26 a | 1490.0 b | 113.8 b | 17.7 a | 3.77 a | |
| 120 | 9.3 a | 3.42 a | 46.81 a | 1601.7 a | 120.2 a | 17.6 a | 3.79 a | |
| 150 | 9.3 a | 3.40 a | 46.70 a | 1589.4 a | 118.2 a | 17.7 a | 3.78 a | |
| ANOVA Results | ||||||||
| P Rate | ** | ** | ** | ** | ** | ** | NS | |
| Year | NS | ** | * | ** | * | ** | NS | |
| P Rate * Year | NS | NS | NS | NS | NS | NS | NS | |
| Flowering Stage | Maturity Stage | ||||||
|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Shell | Seed | |
| Biomass Accumulation | |||||||
| P Rate | ** | ** | ** | ** | ** | ** | ** |
| Year | * | ** | NS | NS | NS | ** | ** |
| P Rate * Year | * | NS | NS | ** | * | NS | NS |
| P Accumulation | |||||||
| P Rate | ** | ** | ** | ** | ** | ** | ** |
| Year | * | * | NS | * | NS | * | * |
| P Rate * Year | * | NS | NS | ** | * | * | * |
| Biomass Partitioning | |||||||
| P Rate | ** | ** | ** | ** | ** | ** | ** |
| Year | NS | NS | NS | NS | * | NS | NS |
| P Rate * Year | NS | NS | NS | NS | NS | NS | NS |
| P Partitioning | |||||||
| P Rate | ** | ** | ** | NS | * | ** | ** |
| Year | NS | NS | NS | NS | NS | NS | NS |
| P Rate * Year | * | * | * | NS | NS | NS | NS |
| Year | P Rate (kg P2O5 ha−1) | Flowering Stage | Maturity Stage | |||||
|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Shell | Seed | ||
| 2018–2019 | 0 | 8.74 c | 44.34 a | 46.92 c | 7.14 c | 42.67 a | 24.33 a | 25.86 c |
| 30 | 9.76 b | 45.21 a | 45.03 c | 7.35 bc | 41.64 b | 24.28 a | 26.73 b | |
| 60 | 9.87 b | 42.47 b | 47.66 b | 7.40 b | 41.38 b | 23.99 ab | 27.22 ab | |
| 90 | 9.77 b | 41.10 c | 49.13 a | 7.57 b | 41.08 bc | 23.82 b | 27.54 ab | |
| 120 | 10.20 a | 41.17 c | 48.63 a | 7.92 a | 40.50 c | 23.66 b | 27.92 a | |
| 150 | 10.28 a | 41.12 c | 48.60 a | 7.94 a | 40.40 c | 23.61 b | 28.06 a | |
| 2019–2020 | 0 | 8.92 c | 46.07 a | 45.01 c | 7.13 c | 42.43 a | 24.38 a | 26.06 c |
| 30 | 9.91 b | 45.35 a | 44.74 c | 7.46 b | 41.56 b | 24.25 a | 26.73 b | |
| 60 | 10.36 a | 42.74 b | 46.91 b | 7.55 b | 41.36 b | 23.95 a | 27.14 b | |
| 90 | 10.12 ab | 41.64 c | 48.25 a | 7.64 ab | 40.65 c | 23.80 a | 27.92 a | |
| 120 | 10.26 ab | 41.22 c | 48.51 a | 7.81 a | 40.11 c | 23.75 a | 28.33 a | |
| 150 | 10.13 ab | 41.46 c | 48.42 a | 7.82 a | 40.18 c | 23.79 a | 28.21 a | |
| Year | P Rate (kg P2O5 ha−1) | Flowering Stage | Maturity Stage | |||||
|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Shell | Seed | ||
| 2018–2019 | 0 | 5.24 d | 45.14 a | 49.62 b | 3.05 a | 13.52 c | 14.40 a | 69.04 b |
| 30 | 5.95 c | 45.80 a | 48.25 c | 2.96 a | 14.98 a | 13.08 b | 68.99 b | |
| 60 | 6.33 b | 42.40 b | 51.27 a | 2.96 a | 14.94 a | 12.41 c | 69.68 b | |
| 90 | 6.33 b | 41.48 b | 52.19 a | 3.03 a | 14.99 a | 12.21 c | 69.77 b | |
| 120 | 6.58 a | 42.04 b | 51.38 a | 3.00 a | 13.99 b | 11.72 d | 71.28 a | |
| 150 | 6.68 a | 41.62 b | 51.70 a | 3.05 a | 14.28 b | 11.34 d | 71.33 a | |
| 2019–2020 | 0 | 5.31 d | 46.39 a | 48.30 c | 3.02 a | 13.66 d | 14.21 a | 69.10 c |
| 30 | 6.15 c | 45.49 b | 48.37 c | 2.98 ab | 14.80 a | 13.14 b | 69.08 c | |
| 60 | 6.47 b | 43.42 c | 50.10 b | 2.99 ab | 14.71 ab | 12.45 c | 69.84 c | |
| 90 | 6.65 a | 42.00 d | 51.35 a | 2.99 ab | 14.17 bcd | 11.80 d | 71.04 b | |
| 120 | 6.65 a | 41.47 d | 51.88 a | 2.89 b | 13.82 cd | 11.34 e | 71.95 a | |
| 150 | 6.61 a | 41.86 d | 51.53 a | 3.05 a | 14.26 abc | 11.51 de | 71.19 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zheng, J.; You, J.; Li, J.; Qian, C.; Leng, S.; Yang, G.; Zuo, Q. Effects of Phosphorus Supply on the Leaf Photosynthesis, and Biomass and Phosphorus Accumulation and Partitioning of Canola (Brassica napus L.) in Saline Environment. Agronomy 2021, 11, 1918. https://doi.org/10.3390/agronomy11101918
Wang L, Zheng J, You J, Li J, Qian C, Leng S, Yang G, Zuo Q. Effects of Phosphorus Supply on the Leaf Photosynthesis, and Biomass and Phosphorus Accumulation and Partitioning of Canola (Brassica napus L.) in Saline Environment. Agronomy. 2021; 11(10):1918. https://doi.org/10.3390/agronomy11101918
Chicago/Turabian StyleWang, Long, Jingdong Zheng, Jingjing You, Jing Li, Chen Qian, Suohu Leng, Guang Yang, and Qingsong Zuo. 2021. "Effects of Phosphorus Supply on the Leaf Photosynthesis, and Biomass and Phosphorus Accumulation and Partitioning of Canola (Brassica napus L.) in Saline Environment" Agronomy 11, no. 10: 1918. https://doi.org/10.3390/agronomy11101918