
The Addition of Selenium to the Nutrient Solution Decreases Cadmium Toxicity in Pepper Plants Grown under Hydroponic Conditions

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growing Conditions and Plant Material
2.2. Treatment with Cadmium and Selenium
2.3. Parameters Analysed
2.3.1. Gas Exchange, Chlorophyll Fluorescence, and Chlorophyll Content Parameters
2.3.2. Growth Parameters
2.3.3. Mineral Analysis in Leaves and Root
2.3.4. Determination of Proline in Leaves and Roots
2.3.5. Determination of Oxidative Damage in Leaves
2.3.6. Determination of Antioxidant Activity
2.3.7. Determination of Antioxidant Enzymes
2.4. Experimental Design and Statistical Analysis
3. Results
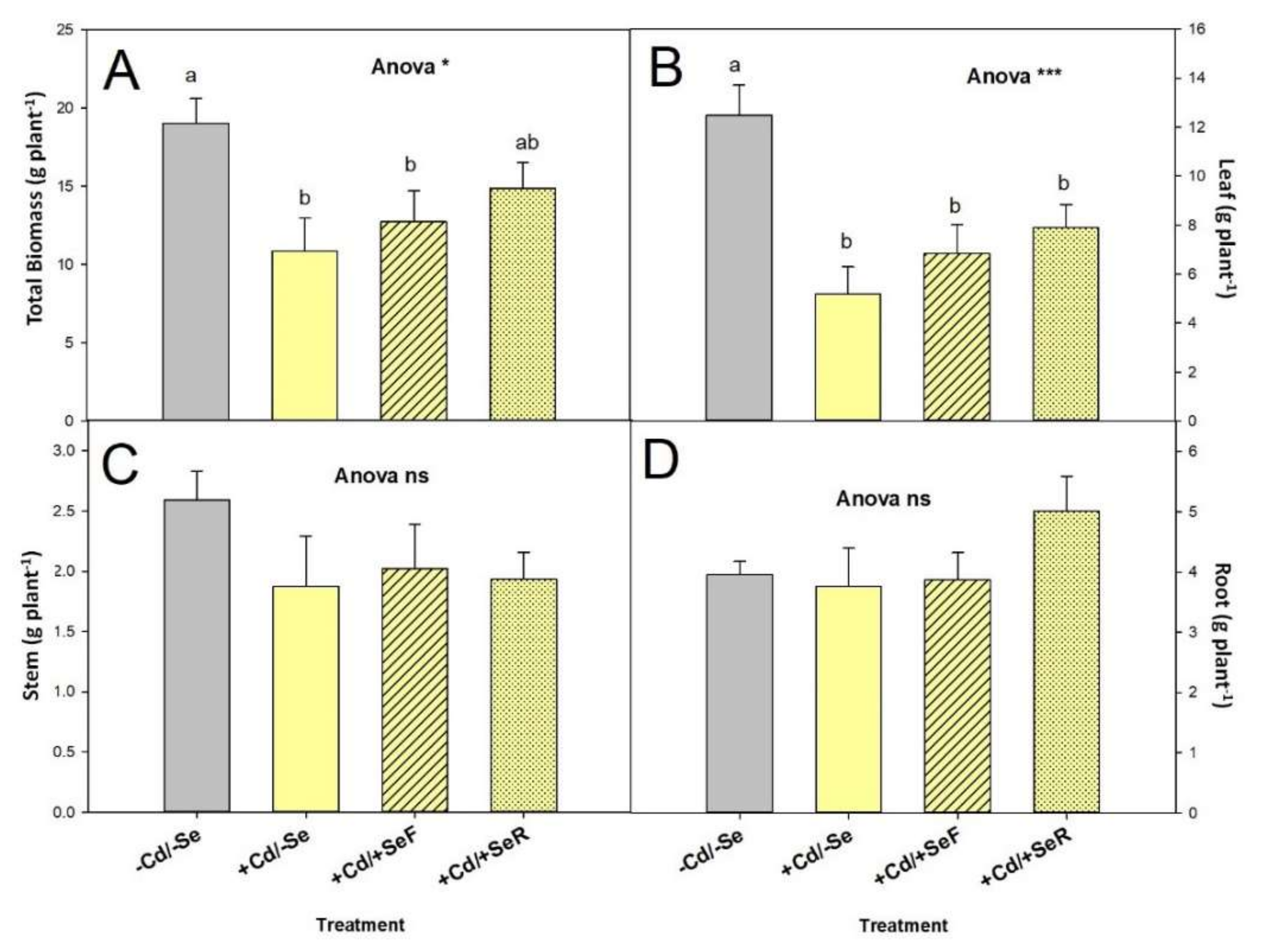
3.1. Growth Parameters
3.2. Concentration of Cd and Se in the Different Tissues of the Plant
3.3. Parameters of Gas Exchange, Chlorophyll Fluorescence and Chlorophyll Content
3.4. Proline in Leaves and Roots
3.5. Mineral Nutrition: Concentration of Macro and Micronutrients in Leaf and Roots
3.6. Oxidative Damage and Antioxidant Activity
3.7. Visual Symptoms of Cd
4. Discussion
4.1. Response of the Cristal Variety Pepper Plants to the Excess of Cd in the Nutrient Solution
4.2. The Application of Se Reduces the Negative Effects of Cd in Pepper Plants of the Variety Cristal
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rai, P.K.; Lee, S.S.; Zhang, M.; Tsang, Y.F.; Kim, K.H. Heavy metals in food crops: Health risks, fate, mechanisms, and management. Environ. Int. 2019, 125, 365–385. [Google Scholar] [CrossRef]
- United States Envionmental Protection Agency (EPA). Available online: https://www.epa.gov/sites/default/files/2016-09/documents/cadmium-compounds.pdf (accessed on 26 July 2021).
- Franic, M.; Galic, V. As, Cd, Cr, Cu, Hg: Physiological Implications and Toxicity in Plants. In Plant Metallomics and Functional Omics; Springer: Cham, The Netherlands, 2019; pp. 209–251. [Google Scholar]
- Kumar, S.; Sharma, A. Cadmium toxicity: Effects on human reproduction and fertility. Rev. Environ. Health 2019, 34, 327–338. [Google Scholar] [CrossRef]
- Shiyu, N.; Hongen, U.; Zhaojun, E.; Rengel, Z.; Wei, O.; Chang, I.; Peng, Z.H.A. Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: A review. Pedosphere 2020, 30, 168–180. [Google Scholar] [CrossRef]
- Solís-Domínguez, F.A.; González-Chávez, M.C.; Carrillo-González, R.; Rodríguez-Vázquez, R. Accumulation and localization of cadmium in Echinochloa polystachya grown within a hydroponic system. J. Hazard Mater. 2007, 141, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Duan, S.; Wu, Q.; Yu, M.; Shabala, S. Reducing Cadmium Accumulation in Plants: Structure–Function Relations and Tissue-Specific Operation of Transporters in the Spotlight. Plants 2020, 9, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astolfi, S.; Zuchi, S.; Passera, C. Effect of cadmium on H+/ATPase activity of plasma membrane vesicles isolated from roots of different S-supplied maize (Zea mays L.) plants. Plant Sci. 2005, 169, 361–368. [Google Scholar] [CrossRef]
- Parmar, P.; Kumari, N.; Sharma, V. Structural and functional alterations in photosynthetic apparatus of plants under cadmium stress. Bot. Stud. 2013, 54, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Pardo, B.; Carpena, R.O.; Zornoza, P. Cadmium in white lupin nodules: Impact on nitrogen and carbon metabolism. J. Plant Physiol. 2013, 170, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Irfan, M.; Hayat, S.; Ahmad, A.; Alyemeni, M.N. Soil cadmium enrichment: Allocation and plant physiological manifestations. Saudi J. Biol. Sci. 2013, 20, 1–10. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Rueda-Puente, E.O.; Troyo-Diéguez, E.; Córdoba-Matson, M.V.; Hernández-Montiel, L.G.; Nieto-Garibay, A. Baseline study of morphometric traits of wild Capsicum annuum growing near two biosphere reserves in the Peninsula of Baja California for future conservation management. BMC Plant Biol. 2015, 15, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizwan, M.; Ali, S.; Rehman, M.; Rinklebe, J.; Tsang, D.; Tack, F.; Abbasi, G.; Hussain, A.; Igalavithana, A.; Lee, B.; et al. Effects of selenium on the uptake of toxic trace elements by crop plants: A review. Crit. Rev. Environ. Sci. Technol. 2020, 1–36. [Google Scholar] [CrossRef]
- Chauhan, R.; Awasthi, S.; Srivastava, S.; Dwivedi, S.; Pilon-Smits, E.A.; Dhankher, O.P.; Tripathi, R.D. Understanding selenium metabolism in plants and its role as a beneficial element. Crit. Rev. Environ. Sci. Technol. 2019, 49, 1937–1958. [Google Scholar] [CrossRef]
- Abd_Allahef, A.; Abeer, H.; Alqarawi, A. Mitigation of cadmium induced stress in tomato (Solanum lycopersicum L.) by selenium. Pak. J. Bot. 2016, 48, 953–961. [Google Scholar]
- Yin, H.; Qi, Z.; Li, M.; Ahammed, G.; Chu, X.; Zhou, J. Selenium forms and methods of application differentially modulate plant growth, photosynthesis, stress tolerance, selenium content and speciation in Oryza sativa L. Ecotoxicol. Environ. Saf. 2019, 169, 911–917. [Google Scholar] [CrossRef]
- Alfosea-Simón, M.; Simón-Grao, S.; Zavala-Gonzalez, E.A.; Cámara-Zapata, J.M.; Simón, I.; Martínez-Nicolás, J.J.; Lidón, V.; García-Sánchez, F. Physiological, nutritional and metabolomic responses of tomato plants after the foliar application of amino acids Aspartic acid, Glutamic Acid and Alanine. Front. Plant Sci. 2020, 11, 581234. [Google Scholar] [CrossRef]
- Bates, L.; Waldren, R.; Teare, I. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yang, H.P.; Yan, R.; Chen, H.; Lee, D.H.; Zheng, C. Characteristics of hemicellulose, cellulose and lignin pyrolysis. Fuel 2007, 86, 1781–1788. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactivesubstances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Koleva, I.I.; Vanbeek, T.A.; Linssen, J.P.H.; Degroot, A.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity. A comparative study on 3 testing methods. Int. J. Plant Chem. Biochem. Tech. 2002, 13, 8–17. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, S.K. Assay of catalase and peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Chandlee, J.M.; Scandalios, J.G. Analysis of variants affecting the catalase developmental program in maize scutellum. Theor. Appl. Genet. 1984, 69, 71–77. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts; its inactivation in Ascorbate depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol. 1987, 28, 131–140. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Haynes, W. Tukey’s Test. In Encyclopedia of Systems Biology; Dubitzky, W., Wolkenhauer, O., Cho, K.H., Yokota, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2303–2304. [Google Scholar] [CrossRef]
- Huang, B.; Xin, J.; Dai, H.; Liu, A.; Zhou, W.; Yi, Y.; Liao, K. Root morphological responses of three hot pepper cultivars to Cd exposure and their correlations with Cd accumulation. Environ. Sci. Pollut. Res. 2015, 22, 1151–1159. [Google Scholar] [CrossRef] [PubMed]
- Latef, A. Growth and some physiological activities of pepper (Capsicum annuum L.) in response to cadmium stress and mycorrhizal symbiosis. J. Agric. Sci. Technol. 2013, 15, 1437–1448. [Google Scholar]
- Uzal, O.; Yasar, F. Efects of GA3 hormone treatments on ion uptake and growth of pepper plants under cadmium stress. Appl. Ecol. Environ. Res. 2017, 15, 1347–1357. [Google Scholar] [CrossRef]
- León, A.M.; Palma, J.M.; Corpas, F.J.; Gómez, M.; Romero-Puertas, M.C.; Chatterjee, D.; Mateos, R.M.; Del Río, L.A.; Sandalio, L.M. Antioxidative enzymes in cultivars of pepper plants with different sensitivity to cadmium. Plant Physiol. Biochem. 2002, 40, 813–820. [Google Scholar] [CrossRef]
- Sigel, A.; Sigel, H.; Sigel, R. Metallothioneins and related chelators. In Metal Ions in Life Sciences; Royal Society of Chemistry Publishing: Cambridge, UK, 2009; Volume 5. [Google Scholar] [CrossRef]
- Shanker, K.; Mishra, S.; Srivastava, S.; Srivastava, R.; Dass, S.; Prakash, S.; Srivastava, M.M. Effect of Selenite and Selenate on Plant Uptake of Cadmium by Maice (Zea mays). Bull. Environ. Contam. Toxicol. 1996, 56, 419–424. [Google Scholar] [CrossRef]
- Huang, B.; Xin, J.; Dai, H.; Zhou, W. Effects of interaction between cadmium (Cd) and selenium (Se) on grain yield and Cd and Se accumulation in a hybrid rice (Oryza sativa) system. J. Agric. Food Chem. 2017, 65, 9537–9546. [Google Scholar] [CrossRef]
- Guignardi, Z.; Schiavon, M. Biochemistry of plant selenium uptake and metabolism. In Selenium in Plants; Plant Ecophysiology; Pilon-Smits, E., Winkel, L., Lin, Z.Q., Eds.; Springer: Cham, The Netherlands, 2017; Volume 12, pp. 21–34. [Google Scholar] [CrossRef]
- Drahonovsky, J.; Szakova, J.; Mestek, O.; Tremlova, J.; Kana, A.; Najmanova, J.; Tlustos, P. Selenium uptake, transformation and inter-element interactions by selected wildlife plant species after foliar selenate application. Environ. Exp. Bot. 2016, 125, 12–19. [Google Scholar] [CrossRef]
- Bertoli, A.C.; Cannata, M.G.; Carvalho, R.; Bastos, A.R.R.; Freitas, M.P.; Dos Santos, A.A. Lycopersicon esculentum submitted to Cd-stressful conditions in nutrition solution: Nutrient contents and translocation. Ecotoxicol. Environ. Saf. 2012, 86, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef] [PubMed]
- Bachir, L.D.M.; Wu, F.; Zhang, G.; Wu, H. Genotypic difference in effect of cadmium on development and mineral concentrations of cotton. Commun. Soil Sci. Plant Anal. 2004, 35, 285–299. [Google Scholar] [CrossRef]
- Wu, H.; Wu, F.B.; Zhang, G.; Bachir, D.M. Effect of cadmium on uptake and translo-cation of three microelements in cotton. J. Plant Nutr. 2005, 27, 2019–2032. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatm. | Product | Apical Chl (SPAD) | Medium Chl (SPAD) | Leaf Proline (mg g−1 dw) | Root Proline (mg g−1 dw) |
|---|---|---|---|---|---|
| −Cd | −Se | 28.8 ± 2.9 a | 64.4 ± 3.9 a | 1.54 ± 0.14 b | 0.75 ± 0.05 b |
| +Cd | −Se | 18.9 ± 2.2 b | 29.2 ± 4.0 b | 2.27 ± 0.09 a | 0.80 ± 0.04 b |
| +SeF | 23.5 ± 4.3 ab | 41.9 ± 8.7 b | 1.58 ± 0.09 b | 1.27 ± 0.03 a | |
| +SeR | 26.9 ± 2.1 ab | 30.5 ± 5.0 b | 1.70 ± 0.12 b | 0.72 ± 0.04 b | |
| ANOVA | * | *** | ** | *** |
| Macronutrients Leaf (g 100 g−1 dw) | |||||||
|---|---|---|---|---|---|---|---|
| Cd Treat. | Se Treat | Ca | K | Mg | Na | P | S |
| −Cd | −Se | 2.17 ±0.24 | 5.14 ±0.57 | 0.42 ± 0.05 | 0.03 ± 0.0001 | 0.50 ± 0.08 b | 0.38 ± 0.05 |
| −Se | 2.67 ± 0.49 | 6.94 ± 0.57 | 0.58 ± 0.10 | 0.04 ± 0.0002 | 0.98 ± 0.09 a | 0.67 ± 0.12 | |
| +Cd | +SeFoliar | 2.85 ± 0.18 | 5.94 ± 0.17 | 0.51 ± 0.04 | 0.03 ± 0.0003 | 0.59 ± 0.06 b | 0.65 ± 0.13 |
| +SeRoot | 2.85 ± 0.10 | 5.80 ± 0.19 | 0.53 ± 0.01 | 0.03 ± 0.0004 | 0.53 ± 0.08 b | 0.81 ± 0.09 | |
| ANOVA | ns | ns | ns | ns | *** | ns | |
| Macronutrients Root (g 100 g−1 dw) | |||||||
| Cd Treat. | Se Treat | Ca | K | Mg | Na | P | S |
| −Cd | −Se | 3.70 ± 0.09 b | 4.49 ± 0.17 a | 0.47 ± 0.02 | 0.07 ± 0.002 a | 1.82 ± 0.13 bc | 0.37 ± 0.01 c |
| −Se | 4.00 ± 0.31 ab | 4.32 ± 0.21 a | 0.48 ± 0.02 | 0.05 ± 0.001 b | 2.20 ± 0.25 b | 0.60 ± 0.03 a | |
| +Cd | +SeFoliar | 1.32 ± 0.27 c | 3.83 ± 0.16 ab | 0.50 ± 0.07 | 0.04 ± 0.002 b | 1.13 ± 0.11 c | 0.50 ± 0.02 ab |
| +SeRoot | 5.45 ± 0.72 a | 3.52 ± 0.16 b | 0.46 ± 0.02 | 0.06 ± 0.002 ab | 3.19 ± 0.33 a | 0.46 ± 0.03 bc | |
| ANOVA | *** | ** | ns | *** | *** | *** | |
| Micronutrients Leaf (mg kg−1 dw) | |||||||
| Cd Treat. | Se Treat | B | Fe | Mn | Zn | Cu | |
| −Cd | −Se | 25.7 ± 2.6 | 139.9 ± 8.1 ab | 50.6 ± 5.7 | 21.9 ± 2.9 | 2.62 ± 0.28 a | |
| −Se | 33.5 ± 3.2 | 119.3 ± 13.6 ab | 66.8 ± 10.5 | 15.1 ± 1.7 | 2.45 ± 0.25 ab | ||
| +Cd | +SeFoliar | 29.6 ± 3.3 | 144.3 ± 5.2 a | 60.4 ± 3.4 | 17.9 ± 2.7 | 1.65 ± 0.22 b | |
| +SeRoot | 27.9 ± 2.0 | 107.8 ± 14.6 b | 53.5 ± 2.1 | 21.2 ± 2.2 | 1.97 ± 0.13 ab | ||
| ANOVA | ns | * | ns | ns | * | ||
| Micronutrients Roots (mg kg−1 dw) | |||||||
| Cd Treat. | Se Treat | B | Fe | Mn | Zn | Cu | |
| −Cd | −Se | 15.1 ± 1.3 | 723.9 ± 52.6 b | 134.8 ± 4.5 b | 120.8 ± 15.7 b | 20.6 ± 1.4 a | |
| −Se | 17.3 ± 0.6 | 856.1 ± 78.5 a | 128.1 ± 12.2 b | 193.8 ± 7.8 a | 19.8 ± 0.6 a | ||
| +Cd | +SeFoliar | 13.7 ± 0.7 | 667.1 ± 87.1 b | 118.3 ± 20.6 b | 120.6 ± 10.1 b | 16.9 ± 0.7 ab | |
| +SeRoot | 15.7 ± 1.3 | 891.7 ± 61.2 a | 218.9 ± 15.8 a | 200.2 ± 17.9 a | 15.7 ± 0.7 b | ||
| ANOVA | ns | * | *** | *** | *** | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Millan, R.; Alfosea-Simon, M.; Zavala-Gonzalez, E.A.; Camara-Zapata, J.M.; Martinez-Nicolas, J.J.; Lidon, V.; Simon, I.; Shahid, M.A.; Garcia-Sanchez, F.; Simon-Grao, S. The Addition of Selenium to the Nutrient Solution Decreases Cadmium Toxicity in Pepper Plants Grown under Hydroponic Conditions. Agronomy 2021, 11, 1905. https://doi.org/10.3390/agronomy11101905
Perez-Millan R, Alfosea-Simon M, Zavala-Gonzalez EA, Camara-Zapata JM, Martinez-Nicolas JJ, Lidon V, Simon I, Shahid MA, Garcia-Sanchez F, Simon-Grao S. The Addition of Selenium to the Nutrient Solution Decreases Cadmium Toxicity in Pepper Plants Grown under Hydroponic Conditions. Agronomy. 2021; 11(10):1905. https://doi.org/10.3390/agronomy11101905
Chicago/Turabian StylePerez-Millan, Rafael, Marina Alfosea-Simon, Ernesto Alejandro Zavala-Gonzalez, Jose Maria Camara-Zapata, Juan Jose Martinez-Nicolas, Vicente Lidon, Inma Simon, Muhammad Adnan Shahid, Francisco Garcia-Sanchez, and Silvia Simon-Grao. 2021. "The Addition of Selenium to the Nutrient Solution Decreases Cadmium Toxicity in Pepper Plants Grown under Hydroponic Conditions" Agronomy 11, no. 10: 1905. https://doi.org/10.3390/agronomy11101905