The Strategy for Marker-Assisted Breeding of Anthocyanin-Rich Spring Bread Wheat (Triticum aestivum L.) Cultivars in Western Siberia

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
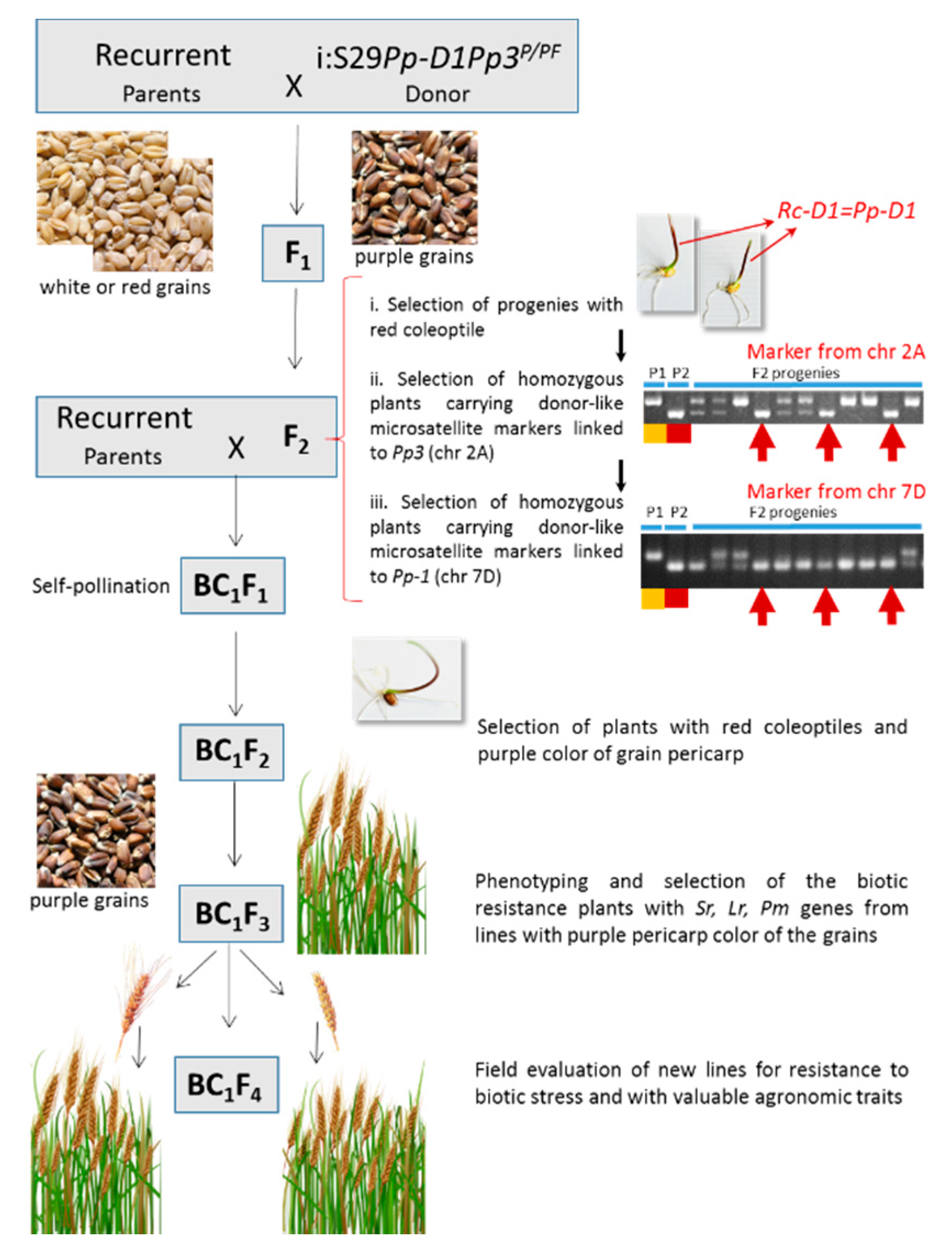
2.1. Plant Material and Breeding Strategy
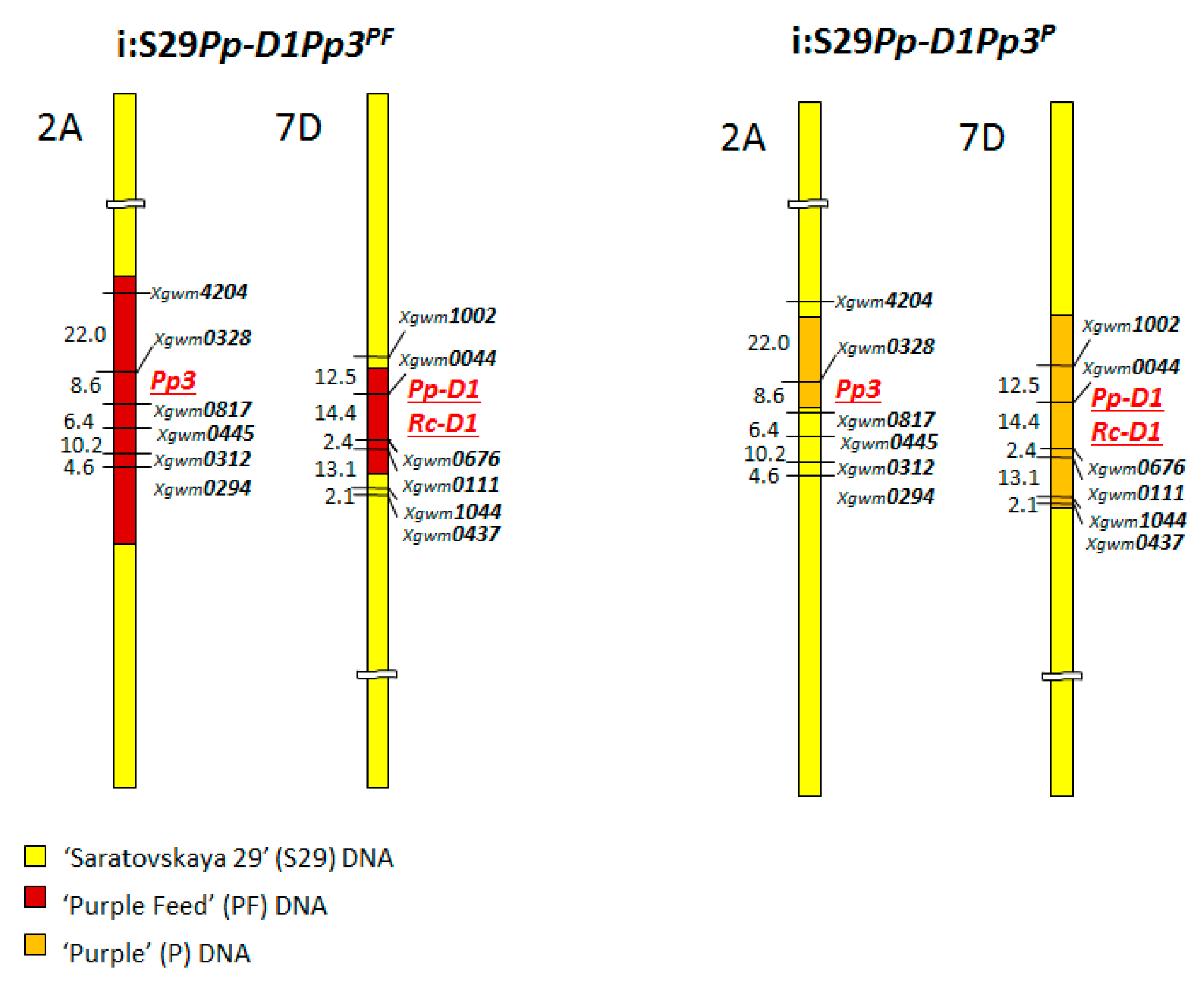
2.2. Molecular Markers
2.3. Morphological Markers
2.4. Sowing, Phenotyping, and Grain Quality Analysis
3. Results
3.1. Marker-Assisted Selection
3.2. Field Screening and Characterization of the Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeding Number | Cultivar/BC1F4 Line | Productivity, g/m2 | Protein,% | Gluten,% | Resistance * to | ||
|---|---|---|---|---|---|---|---|
| Powdery Mildew | Stem Rust | Leaf Rust | |||||
| cv Pamyati Aziyeva, check | 374 | 12.65 | 20.31 | MS | 20 MS | 20 MS | |
| cv Duet, check | 380 | 13.61 | 24.75 | MR | 50 S | 10 M | |
| cv Element22, check | 490 | 15.59 | 29.84 | MR | 5 M | 5 M | |
| 235 | (cv Element22 × i:S29PF) | 483 | 15.56 | 23.75 | MR | 30 MS | 10 MS |
| 236 | (cv Element22 × i:S29PF) | 452 | 16.19 | 24.61 | MR | 20 MS | 50 S |
| 237 | (cv Element22 × i:S29PF) | 490 | 16.32 | 26.22 | MR | 40 S | 20 S |
| 238 | (cv Element22 × i:S29PF) | 510 | 16.02 | 25.18 | MR | 5 M | 5 M |
| 239 | (cv Element22 × i:S29PF) | 687 | 14.99 | 23.34 | MR | 5 M | 10 MS |
| 240 | (cv Element22 × i:S29PF) | 522 | 15.46 | 21.85 | MR | 20 MS | 5 M |
| 249 | (cv Element22 × i:S29PF) | 617 | 16.12 | 26.29 | MR | 10 M | 15 M |
| 250 | (cv Element22 × i:S29PF) | 539 | 16.71 | 24.77 | MR | 20 MS | 15 MS |
| 251 | (cv Element22 × i:S29PF) 4 | 574 | 15.13 | 20.27 | MR | 40 S | 10 M |
| 253 | (cv Element22 × i:S29PF) | 593 | 15.54 | 23.95 | MR | 20 MS | 10 MS |
| 254 | (cv Element22 × i:S29PF) | 647 | 15.30 | 23.24 | MR | 10 MS | 10 MS |
| 255 | (cv Element22 × i:S29PF) | 597 | 16.39 | 26.63 | MR | 5 M | 10 MS |
| 256 | (cv Element22 × i:S29PF) | 468 | 14.03 | 32.84 | MR | 30 MS | 5 M |
| 257 | (cv Element22 × i:S29PF) | 304 | 15.62 | 22.24 | MR | 90 S | 0 R |
| 258 | (cv Element22 × i:S29PF) | 367 | 15.75 | 25.54 | MR | 80 S | 40 S |
| 259 | (cv Element22 × i:S29PF) | 409 | 13.18 | 22.17 | MR | 70 S | 5 M |
| 260 | (cv Element22 × i:S29PF) | 650 | 14.87 | 23.81 | MR | 10 M | 10 M |
| 261 | (cv Element22 × i:S29PF) | 513 | 16.32 | 26.22 | MS | 60 S | 5 M |
| 262 | (cv Element22 × i:S29PF) | 457 | 15.85 | 24.17 | MR | 40 S | 5 M |
| 263 | (cv Element22 × i:S29PF) | 499 | 15.26 | 22.38 | MR | 30 MS | 10 MS |
| 264 | (cv Element22 × i:S29PF) | 546 | 19.18 | 22.17 | MS | 50 S | 10 MS |
| 265 | (cv Element22 × i:S29PF) | 515 | 15.40 | 24.03 | MS | 30 S | 5 M |
| 266 | (cv Element22 × i:S29PF) | 570 | 15.34 | 24.45 | MR | 5 M | 5 M |
| 267 | (cv Element22 × i:S29PF) | 548 | 14.21 | 23.20 | MR | 60 S | 5 M |
| 268 | (STRU 0732 × i:S29PF) | 391 | 12.96 | 21.48 | MS | 80 S | 0 R |
| 269 | (STRU 0732 × i:S29PF) | 238 | 12.55 | 18.63 | R | 90 S | 30 MS |
| 270 | (STRU 0732 × i:S29PF) | 304 | 12.49 | 19.41 | MR | 90 S | 0 R |
| 271 | (STRU 0732 × i:S29PF) | 209 | 14.87 | 25.46 | MR | 90 S | 10 M |
| 272 | (STRU 0732 × i:S29PF) | 852 | 16.73 | 23.44 | R | 90 S | 5 M |
| 273 | (STRU 0732 × i:S29PF) | 162 | 14.20 | 26.15 | MR | 90 S | 5 M |
| 274 | (BW49880 × i:S29PF) | 370 | 15.97 | 26.13 | MR | 20 MS | 10 MS |
| 275 | (BW49880 × i:S29PF) | 440 | 16.19 | 26.62 | MR | 10 MS | 0 R |
| 279 | (BW49880 × i:S29P) | 357 | 17.46 | 28.98 | 5 | 10 M | 0 R |
| 281 | (BW49880 × i:S29PF) | 417 | 18.05 | 29.29 | MR | 5 M | 5 M |
| 294 | (cv Aina × i:S29PF) | 400 | 13.26 | 20.14 | MR | 80 S | 0 R |
| 295 | (cv Aina × i:S29PF) | 391 | 14.29 | 21.16 | MR | 90 S | 0 R |
| 296 | (cv Aina × i:S29PF) | 379 | 14.57 | 22.39 | MR | 90 S | 0 R |
| 297 | (cv Aina × i:S29PF) | 522 | 14.38 | 21.57 | MS | 60 S | 5 MS |
| 298 | (cv Aina × i:S29PF) | 461 | 14.03 | 21.33 | MS | 40 S | 15 S |
| 299 | (cv Aina × i:S29P) | 400 | 13.24 | 19.99 | MS | 90 S | 5 MS |
| 300 | (cv Aina × i:S29P) | 261 | 13.72 | 21.17 | MS | 80 S | 10 MS |
| 301 | (cv Aina × i:S29P) | 417 | 13.75 | 20.85 | MS | 80 S | 5 MS |
| 302 | (cv Aina × i:S29P) | 235 | 14.26 | 21.18 | MS | 90 S | 5 MS |
| 303 | (cv Aina × i:S29P) | 327 | 13.90 | 20.61 | MS | 90 S | 0 R |
| 304 | (cv Aina × i:S29P) | 391 | 14.26 | 23.20 | MR | 90 S | 0 R |
| 305 | (cv Aina × i:S29P) | 348 | 13.71 | 20.98 | MS | 90 S | 0 R |
| 306 | (cv Tobol’skaya × i:S29PF) | 346 | 13.08 | 19.73 | MR | 90 S | 0 R |
| 307 | (cv Tobol’skaya × i:S29PF) | 318 | 12.22 | 20.48 | MS | 90 S | 0 R |
| 310 | (cv Tobol’skaya × i:S29PF) | 330 | 13.26 | 19.68 | MR | 90 S | 0 R |
| 311 | (cv Tobol’skaya × i:S29PF) | 357 | 14.07 | 21.42 | MR | 90 S | 40 S |
| 312 | (cv Tobol’skaya × i:S29PF) | 330 | 12.88 | 20.45 | MS | 90 S | 40 S |
| 313 | (cv Tobol’skaya × i:S29PF) | 228 | 13.60 | 19.92 | MS | 90 S | 0 R |
| 314 | (cv Tobol’skaya × i:S29PF) | 323 | 13.25 | 20.11 | MR | 90 S | 0 R |
| 315 | (cv Tobol’skaya × i:S29PF) | 209 | 14.10 | 24.37 | MS | 90 S | 0 R |
| 316 | (cv Tobol’skaya × i:S29PF) | 296 | 12.89 | 20.96 | MS | 90 S | 0 R |
| 317 | (cv Tobol’skaya × i:S29 PF) | 174 | 14.48 | 23.86 | S | 90 S | 0 R |
| 318 | (cv Tobol’skaya × i:S29PF) | 278 | 13.60 | 21.04 | MS | 90 S | 0 R |
| 319 | (cv Tobol’skaya × i:S29PF) | 287 | 14.38 | 24.89 | MS | 30 MS | 0 R |


References
- Weststrate, J.A.; van Poppe, G.; Verschuren, P.M. Functional foods, trends and future. Br. J. Nutr. 2002, 88, S233–S235. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aal, E.S.M.; Young, J.C.; Rabalski, I. Anthocyanin composition in black, blue, pink, purple, and red cereal grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Liu, F.; Tong, L.; Chen, Z.; Chen, J.; Huang, C. Neuroprotective effects of anthocyanins and its major component cyanidin-3-O-glucoside (C3G) in the central nervous system: An outlined review. Eur. J. Pharmacol. 2019, 858, 172500. [Google Scholar] [CrossRef]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Poutanen, K.; Mykkänen, H.; Niskanen, L. Berries reduce postprandial insulin responses to wheat and rye breads in healthy women. J. Nutr. 2013, 143, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Tikhonova, M.A.; Amstislavskaya, T.G.; Akopyan, A.A.; Ovsyukova, M.V.; Tenditnik, M.V.; Khlestkina, E.K. Effects of diets rich in plant polyphenols in mouse models of neurodegenerative disorders. In Proceedings of the 12th International Multiconference: Bioinformatics of Genome Regulation and Structure/Systems Biology (BGRS/SB-2020), Novosibirsk, Russia, 6–10 July 2020; p. 292. [Google Scholar] [CrossRef]
- Tenditnik, M.V.; Tikhonova, M.A.; Litvinova, E.A.; Popova, N.A.; Amstislavskaya, T.G.; Khlestkina, E.K. Effects of anthocyanin-rich grain diet on growth and metastasis of Lewis lung carcinoma in mice. In Proceedings of the 12th International Multiconference: Bioinformatics of Genome Regulation and Structure/Systems Biology (BGRS/SB-2020), Novosibirsk, Russia, 6–10 July 2020; p. 359. [Google Scholar] [CrossRef]
- Gamel, T.H.; Abdel-Aal, E.S.M.; Tucker, A.J.; Pare, S.M.; Faughnan, K.; O’Brien, C.D.; Wright, A.J. Consumption of whole purple and regular wheat modestly improves metabolic markers in adults with elevated high-sensitivity C-reactive protein: A randomised, single-blind parallel-arm study. Br. J. Nutr. 2020, 1–11. [Google Scholar] [CrossRef]
- Zeven, A.C. Wheats with purple and blue grains: A review. Euphytica 1991, 56, 243–258. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Shishkina, A.A.; Goncharov, N.P.; Zuev, E.V.; Lysenko, N.S.; Mitrofanova, O.P.; Dragovich, A.Y.; Kudriavtsev, A.M. Evolution of Triticum aethiopicum Jakubz. from the position of chromosome analysis. Russ. J. Genet. 2018, 54, 629–642. [Google Scholar] [CrossRef]
- Belay, G.; Tesemma, T.; Bechere, E.; Mitiku, D. Natural and human selection for purple-grain tetraploid wheats in the Ethiopian highlands. Gen. Res. Crop Evol. 1995, 42, 387–391. [Google Scholar] [CrossRef]
- Copp, L.G.L. Purple grain in hexaploid wheat. Wheat Inf. Serv. 1965, 20, 18. [Google Scholar]
- Jensen, N.F. Registration of Charcoal wheat germplasm. Crop Sci. 1977, 17, 983. [Google Scholar] [CrossRef]
- Morin, C. Crop Development Breeds Patience for USask Wheat Researcher. 2019. Available online: https://agbio.usask.ca/news/2019/12/news-crop-development-breeds-patience-for-usask-wheat-researcher.php (accessed on 20 August 2020).
- Griffin, W.B. Outcrossing in New Zealand wheats measured by occurrence of purple grain. New Zealand J. Agric. Res. 1987, 30, 287–290. [Google Scholar] [CrossRef]
- Syed Jaafar, S.N.; Baron, J.; Siebenhandl-Ehn, S.; Rosenau, T.; Böhmdorfer, S.; Grausgruber, H. Increased anthocyanin content in purple pericarp× blue aleurone wheat crosses. Plant Breed. 2013, 132, 546–552. [Google Scholar] [CrossRef]
- King, C. New Possibilities with Purple Wheat. 5 May 2017. Available online: https://www.topcropmanager.com/new-possibilities-with-purple-wheat-20050/ (accessed on 20 September 2020).
- Li, W.; Pickard, M.D.; Beta, T. Evaluation of antioxidant activity and electronic taste and aroma properties of antho-beers from purple wheat grain. J. Agric. Food Chem. 2007, 55, 8958–8966. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Garcia, J.A.A.; Correa, V.G.; Vieira, T.F.; Bracht, A.; Peralta, R.M. Pigments and vitamins from plants as functional ingredients: Current trends and perspectives. Adv. Food Nutr. Res. 2019, 90, 259–303. [Google Scholar] [CrossRef]
- Bartl, P.; Albreht, A.; Skrt, M.; Tremlová, B.; Ošťádalová, M.; Šmejkal, K.; Ulrih, N.P. Anthocyanins in purple and blue wheat grains and in resulting bread: Quantity, composition, and thermal stability. Int. J. Food Sci. Nutr. 2015, 66, 514–519. [Google Scholar] [CrossRef]
- Khlestkina, E.K.; Usenko, N.I.; Gordeeva, E.I.; Stabrovskaya, O.I.; Sharfunova, I.B.; Otmakhova, Y.S. Evaluation of wheat products with high flavonoid content: Justification of importance of marker-assisted development and production of flavonoid-rich wheat cultivars. Vavilov. J. Genet. Breed. 2017, 21, 545–553. [Google Scholar] [CrossRef]
- Pasqualone, A.; Bianco, A.M.; Paradiso, V.M.; Summo, C.; Gabarcorta, G.; Caponio, F.; Blanco, A. Production and characterization of functional biscuits obtained from purple wheat. Food Chem. 2015, 180, 64–70. [Google Scholar] [CrossRef]
- Usenko, N.I.; Khlestkina, E.K.; Asavasanti, S.; Gordeeva, E.I.; Yudina, R.S.; Otmakhova, Y.S. Possibilities of enriching food products with anthocyanins by using new forms of cereals. Foods Raw Mater. 2018, 6, 128–135. [Google Scholar] [CrossRef]
- Ficco, D.B.M.; De Simone, V.; De Leonardis, A.M.; Giovanniello, V.; Del Nobile, M.A.; Padalino, L.; De Vita, P. Use of purple durum wheat to produce naturally functional fresh and dry pasta. Food Chem. 2016, 100, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamel, T.H.; Wright, A.J.; Pickard, M.; Abdel-Aal, E.S.M. Characterization of anthocyanin-containing purple wheat prototype products as functional foods with potential health benefits. Cereal Chem. 2020, 97, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Burešová, I.; Trojan, V.; Helis, M. Characteristics of flour and dough from purple and blue wheat grain. Potravin. Slovak J. Food Sci. 2019, 13, 163–166. [Google Scholar] [CrossRef]
- Morgounov, A.; Karaduman, Y.; Akin, B.; Aydogan, S.; Baenziger, P.S.; Bhatta, M.; Guzman, C. Yield and quality in purple-grained wheat isogenic lines. Agronomy 2020, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Khlestkina, E.K.; Shoeva, O.Y.; Gordeeva, E.I. Flavonoid biosynthesis genes in wheat. Russ. J. Genet. Appl. Res. 2015, 5, 268–278. [Google Scholar] [CrossRef]
- Shoeva, O.; Gordeeva, E.; Khlestkina, E. The regulation of anthocyanin synthesis in the wheat pericarp. Molecules 2014, 19, 20266–20279. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Liu, T.; Nan, W.; Jeewani, D.C.; Niu, Y.; Li, C.; Wang, Y.; Shi, X.; Wang, C.; Wang, J.; et al. Two transcription factors TaPpm1 and TaPpb1 co-regulate anthocyanin biosynthesis in purple pericarps of wheat. J. Exp. Botany 2018, 69, 2555–2567. [Google Scholar] [CrossRef] [Green Version]
- Himi, E.; Taketa, S. Isolation of candidate genes for the barley Ant1 and wheat Rc genes controlling anthocyanin pigmentation in different vegetative tissues. Mol. Genet. Genom. 2015, 290, 1287–1298. [Google Scholar] [CrossRef]
- Gordeeva, E.I.; Glagoleva, A.Y.; Kukoeva, T.V.; Khlestkina, E.K.; Shoeva, O.Y. Purple-grained barley (Hordeum vulgare L.): Marker-assisted development of NILs for investigating peculiarities of the anthocyanin biosynthesis regulatory network. BMC Plant. Biol. 2019, 19, 49–57. [Google Scholar] [CrossRef]
- Arbuzova, V.S.; Maystrenko, O.I.; Popova, O.M. Development of near-isogenic lines of the common wheat cultivar ‘Saratovskaya 29’. Cereal Res. Commun. 1998, 26, 39–46. [Google Scholar] [CrossRef]
- Tereshchenko, O.; Gordeeva, E.; Arbuzova, V.; Börner, A.; Khlestkina, E. The D genome carries a gene determining purple grain colour in wheat. Cereal Res. Com. 2012, 40, 334–341. [Google Scholar] [CrossRef]
- Chatrath, R.; Gupta, V.; Parkash, O.; Singh, G.P. Evaluation of biofortified spring wheat genotypes for yield and micronutrients. J. Appl. Nat. Sci. 2018, 10, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Ghimire, S.; Thapa, D.B.; Paudel, A.; Adhikari, N.R. Variability study of biofortified bread wheat genotypes for grain Zinc and Iron concentration, yield and yield associated traits at Khumaltar, Lalitpur, Nepal. Int. J. Appl. Sci. Biotechnol. 2019, 7, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar]
- Ganal, M.W.; Röder, M.S. Microsatellite and SNP markers in wheat breeding. In Genomics-Assisted Crop Improvement; Springer: Dordrecht, Germany, 2007; pp. 1–24. [Google Scholar]
- Gordeeva, E.I.; Shoeva, O.Y.; Khlestkina, E.K. Marker-assisted development of bread wheat near-isogenic lines carrying various combinations of purple pericarp (Pp) alleles. Euphytica 2015, 203, 469–476. [Google Scholar] [CrossRef]
- Plaschke, J.; Ganal, M.W.; Röder, M.S. Detection of genetic diversity in closely related bread wheat using microsatellite markers. Theor. Appl. Genet. 1995, 91, 1001–1007. [Google Scholar] [CrossRef]
- Koyshybaev, M.; Shamanin, V.P.; Morgunov, A.I. Screening of Wheat for Resistance to Major Diseases; FAO-SEK: Ankara, Turkey, 2014. (In Russian) [Google Scholar]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Martinek, P.; Jirsa, O.; Vaculova, K.; Chrpova, J.; Watanabe, N.; Buresova, V.; Kopecky, D.; Stiasna, K.; Vyhnanek, T.; Trojan, V. Use of wheat gene resources with different colour in breeding. Proc. Tag. Pflanz. 2014, 64, 1–4. [Google Scholar]
- Zykin, V.A.; Shamanin, V.P.; Belan, I.A. Ecology of Wheat: Monograph; Publishing house Omsk state Agrarian University: Omsk, Russia, 2000; p. 124. (In Russian) [Google Scholar]
- Likhenko, I.E.; Sovetov, V.V.; Anosov, S.I.; Likhenko, N.N. Formation of grain yield of Siberian cultivars of spring soft wheat under conditions of continental climate of Western Siberia. Res. Tech. Adv. Agribus. Sector 2014, 1, 27–30. (In Russian) [Google Scholar]
- Shamanin, V.P.; Pototskaya, I.V.; Shepelev, S.S.; Pozherukova, V.E.; Salina, Е.А.; Skolotneva, Е.S.; Morgounov, A.I. Stem rust in Western Siberia–race composition and effective resistance genes. Vavilov. J. Genet. Breed. 2020, 24, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Khlestkina, E.K. Molecular markers in genetic studies and breeding. Russ. J. Genet. Appl. Res. 2014, 4, 236–244. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Cortez, R.; Ongkowijoyo, P.; de Mejia, E.G. Protection of color and chemical degradation of anthocyanin from purple corn (Zea mays L.) by zinc ions and alginate through chemical interaction in a beverage model. Food Res. Int. 2018, 105, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhu, X.; Li, Y.; Wang, C. Analysis of the ph-dependent fe (iii) ion chelating activity of anthocyanin extracted from black soybean [Glycine max (L.) Merr.] coats. J. Agric. Food Chem. 2018, 66, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Shamanin, V.; Salina, E.; Wanyera, R.; Zelenskiy, Y.; Olivera, P.; Morgounov, A. Genetic diversity of spring wheat from Kazakhstan and Russia for resistance to stem rust Ug99. Euphytica 2016, 212, 287–296. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Njau, P.; Wanyera, R.; Ward, R.W. Will stem rust destroy the world’s wheat crop? Adv. Agron. 2008, 98, 271–309. [Google Scholar]
- Babiker, E.; Ibrahim, A.M.; Yen, Y.; Stein, J. Identification of a microsatellite marker associated with stem rust resistance gene ‘sr35’ in wheat. Aust. J. Crop. Sci. 2009, 3, 195–200. [Google Scholar]
- Gupta, S.K.; Charpe, A.; Koul, S.; Haque, Q.M.R.; Prabhu, K.V. Development and validation of SCAR markers co-segregating with an Agropyron elongatum derived leaf rust resistance gene Lr24 in wheat. Euphytica 2006, 150, 233–240. [Google Scholar] [CrossRef]
- Jin, Y.; Szabo, L.J.; Pretorius, Z.A.; Singh, R.P.; Ward, R.; Fetch, T. Detection of virulence to resistance gene Sr24 within race TTKS of Puccinia graminis f. sp tritici. Plant Dis. 2008, 92, 923–926. [Google Scholar] [CrossRef] [Green Version]
- Gultyaeva, E.I.; Shaydayuk, E.L.; Shamanin, V.P.; Akhmetova, A.K.; Tyunin, V.A.; Shreider, E.R.; Morgunov, A.I. Genetic structure of Russian and Kazakhstani leaf rust causative agent Puccinia triticina Erikss. populations as assessed by virulence profiles and SSR markers. Agric. Biol. 2018, 53, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Spielmeyer, W.; Sharp, P.J.; Lagudah, E.S. Identification and validation of markers linked to broad-spectrum stem rust resistance gene Sr2 in wheat (Triticum aestivum L.). Crop. Sci. 2003, 43, 333–336. [Google Scholar]
- Aktar-Uz-Zaman, M.; Tuhina-Khatun, M.; Hanafi, M.M.; Sahebi, M. Genetic analysis of rust resistance genes in global wheat cultivars: An overview. Biotechnol. Biotechnol. Equip. 2017, 31, 431–445. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Espino, J.; Singh, R.; Crespo-Herrera, L.A.; Villaseñor-Mir, H.E.; Rodriguez-Garcia, M.F.; Dreisigacker, S.; Lagudah, E. Adult plant slow rusting genes confer high levels of resistance to rusts in bread wheat cultivars from Mexico. Front. Plant Sci. 2020, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Huerta-Espino, J.; Bhavani, S.; Herrera-Foessel, S.A.; Singh, D.; Singh, P.K.; Crossa, J. Race non-specific resistance to rust diseases in CIMMYT spring wheats. Euphytica 2011, 179, 175–186. [Google Scholar] [CrossRef]

| Cultivar/Line | Seed Coat Color | Pedigree | Characterization | Origin |
|---|---|---|---|---|
| cv Element 22 | Red | GRANIT/SARATOVSKAYA 29/3/ERYTHROSPERMUM 59//TSELINNAYA 20/TERTSIYA | Mid-late ripening, resistance genes: Sr35 (3AL), Lr26, Sr31, Yr9, Pm8 (1BS) | P.A. Stolypin Omsk State Agrarian University, Russia |
| cv Aina | Red | TSELINNAYA YUBILIENAYA/2*PASTOR/3/BABAX/LR43//BABAX | Medium ripening, resistance genes: Lr24/Sr24 (3DL) | Karabalyk Agricultural Experimental Station, Kazakhstan |
| cv Tobol’skaya | White | LUTESCENS 123-S/OMSKAYA20 | Mid-late ripening, high drought tolerance, good grain quality, resistance to head smut | Altai Research Institute of Agriculture, and “Kurgansemena” Ltd. |
| BW 49880 | White | VILLA JUAREZ F2009/SOLALA//WBLL1*2/BRAMBLING | Medium early ripening, complex resistance to stem and leaf rust, zinc biofortified grain | Breeding line with high Zn concentration (CIMMYT, INT), successfully used in many breeding programs [37,38]. |
| STRU 0732-84k 071 | Red | - | Medium ripening, good grain quality, short-stemmed | Breeding line was kindly provided by Andreas Jacobi (Strube D&S GmbH, Söllingen, Germany) |
| Chr | GWM Markers |
|---|---|
| 2A | Xgwm 0294, 0312, 0445, 0817, 0328, 4204 |
| 7D | Xgwm 0437, 1044, 0111, 0676, 0044, 1002 |
| No. | Crossing Combinations | Total Number of the F2 Seeds Planted for Coleoptile Color Test | Number of F2 Seedlings with Dark-Red Coleoptile, Planted for SSR-Genotyping and Crossing | Number of F2 Plants Genotyped with SSR-Markers to Select Pp-D1Pp-D1Pp3Pp3 Genotypes | Number of F2 Genotypes Pp-D1Pp-D1Pp3Pp3 Used in Crosses with Recipient |
|---|---|---|---|---|---|
| 1 | cv Aina × i:S29PF | 14 | 10 | 10 | 3 |
| 2 | cv Aina × i:S29P | 8 | 4 | 4 | 1 |
| 3 | BW49880 × i:S29PF | 80 | 50 | 24 | 7 |
| 4 | BW49880 × i:S29P | 80 | 50 | 24 | 6 |
| 5 | cv Element22 × i:S29PF | 80 | 50 | 24 | 3 |
| 6 | cv Element22 × i:S29P | 80 | 50 | 24 | 3 |
| 7 | cv Tobol’skaya × i:S29PF | 18 | 13 | 13 | 4 |
| 8 | cv Tobol’skaya × i:S29P | 80 | 50 | 24 | 7 |
| 9 | STRU 0732-84k 071 × i:S29PF | 80 | 50 | 24 | 6 |
| 10 | STRU 0732-84k 071 × i:S29P | 80 | 50 | 24 | 5 |
| Breeding Number | Cultivar/BC1F4 Line | Grain Weight, g/m2 | Exceeding the Check ± g/m2 | Vegetation Period, Days | Heading Date | Resistance * to | ||
|---|---|---|---|---|---|---|---|---|
| Powdery Mildew | Stem Rust | Leaf Rust | ||||||
| Medium Early | ||||||||
| cv Pamyati Aziyeva, check | 374 | 72 | 10 July | MS | 20 MS | 20 MS | ||
| 275 | (BW49880 × i:S29PF) | 440 | +66 | 72 | 10 July | MR | 10 MS | 0 R |
| 281 | (BW49880 × i:S29PF) | 417 | +43 | 76 | 10 July | MR | 5 M | 5 M |
| Mid-Season | ||||||||
| cv Duet, check | 380 | 78 | 13 July | MR | 50 S | 10 M | ||
| 297 | (cv Aina × i:S29PF) | 522 | +142 | 79 | 10 July | MS | 60 S | 5 MS |
| 298 | (cv Aina × i:S29PF) | 461 | +81 | 79 | 10 July | MS | 40 S | 15 S |
| 301 | (cv Aina × i:S29P) | 417 | +37 | 78 | 12 July | MS | 80 S | 5 MS |
| Mid-Late | ||||||||
| cv Element22, check | 490 | 85 | 15 July | MR | 5 M | 5 M | ||
| 239 | (cv Element22 × i:S29PF) | 687 | +197 | 84 | 14 July | MR | 5 M | 10 MS |
| 240 | (cv Element22 × i:S29PF) | 522 | +32 | 84 | 12 July | MR | 20 MS | 5 M |
| 249 | (cv Element22 × i:S29PF) | 617 | +127 | 86 | 11 July | MR | 10 M | 15 M |
| 250 | (cv Element22 × i:S29PF) | 539 | +49 | 86 | 15 July | MR | 20 MS | 15 MS |
| 251 | (cv Element22 × i:S29PF) | 584 | +94 | 86 | 13 July | MR | 40 S | 10 M |
| 253 | (cv Element22 × i:S29PF) | 593 | +103 | 85 | 15 July | MR | 20 MS | 10 MS |
| 254 | (cv Element22 × i:S29PF) | 647 | +157 | 85 | 13 July | MR | 10 MS | 10 MS |
| 255 | (cv Element22 × i:S29PF) | 597 | +107 | 84 | 17 July | MR | 5 M | 10 MS |
| 260 | (cv Element22 × i:S29PF) | 650 | +160 | 84 | 12 July | MR | 10 M | 10 M |
| 264 | (cv Element22 × i:S29PF) | 546 | +56 | 84 | 11 July | MS | 50 S | 10 MS |
| 266 | (cv Element22 × i:S29PF) | 570 | +80 | 85 | 12 July | MR | 5 M | 5 M |
| 267 | (cv Element22 × i:S29PF) | 548 | +58 | 85 | 13 July | MR | 60 S | 5 M |
| HCP0,05 | 28.5 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordeeva, E.; Shamanin, V.; Shoeva, O.; Kukoeva, T.; Morgounov, A.; Khlestkina, E. The Strategy for Marker-Assisted Breeding of Anthocyanin-Rich Spring Bread Wheat (Triticum aestivum L.) Cultivars in Western Siberia. Agronomy 2020, 10, 1603. https://doi.org/10.3390/agronomy10101603
Gordeeva E, Shamanin V, Shoeva O, Kukoeva T, Morgounov A, Khlestkina E. The Strategy for Marker-Assisted Breeding of Anthocyanin-Rich Spring Bread Wheat (Triticum aestivum L.) Cultivars in Western Siberia. Agronomy. 2020; 10(10):1603. https://doi.org/10.3390/agronomy10101603
Chicago/Turabian StyleGordeeva, Elena, Vladimir Shamanin, Olesya Shoeva, Tatyana Kukoeva, Alexey Morgounov, and Elena Khlestkina. 2020. "The Strategy for Marker-Assisted Breeding of Anthocyanin-Rich Spring Bread Wheat (Triticum aestivum L.) Cultivars in Western Siberia" Agronomy 10, no. 10: 1603. https://doi.org/10.3390/agronomy10101603