
Protective Effect of Sargassum fusiforme Fucoidan against Ethanol-Induced Oxidative Damage in In Vitro and In Vivo Models

 , , , , and
, , , , and 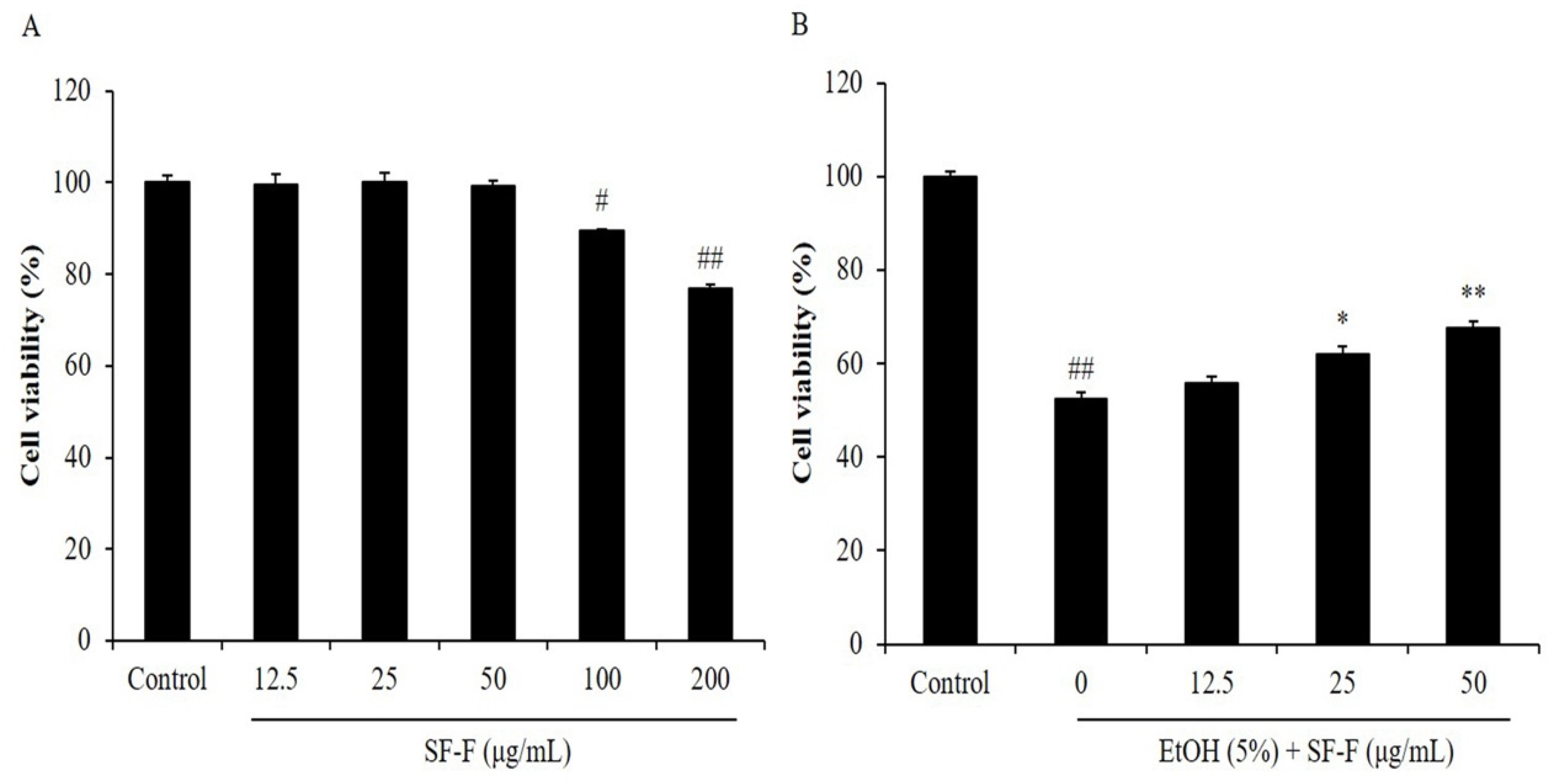{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation and Characterization of SF-F
2.3. Cell Culture
2.4. Measurement of Cytotoxicity and Cytoprotective Effect of SF-F
2.5. Nuclear Staining with Hoechst 33342
2.6. Application of EtOH to Zebrafish Embryos
2.7. Measurement of ROS Generation, Cell Death, and Lipid Peroxidation in Zebrafish
2.8. Statistical Analysis
3. Results and Discussion
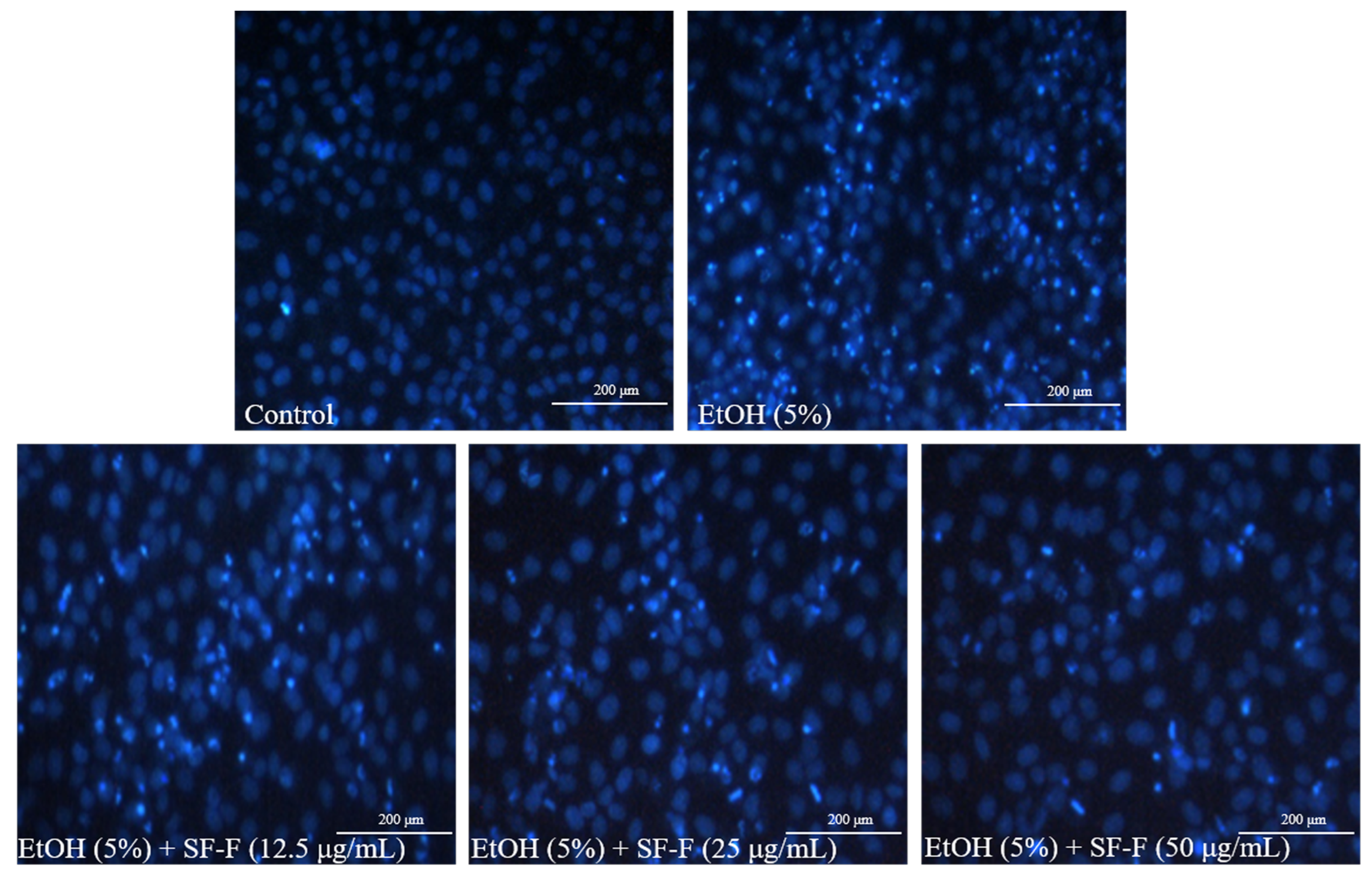
3.1. SF-F Protects Chang Liver Cells against Oxidative Stress Stimulated by EtOH
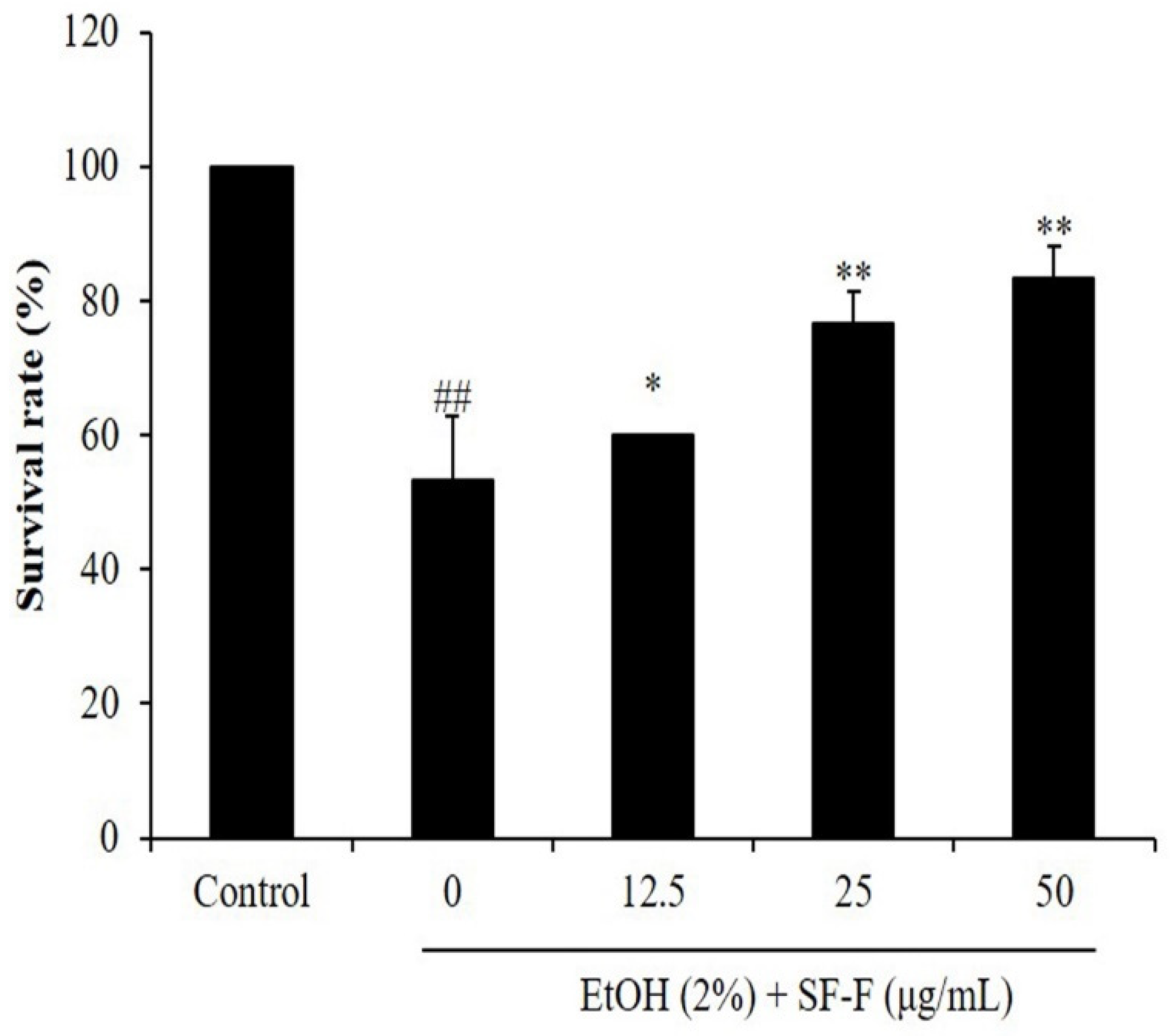
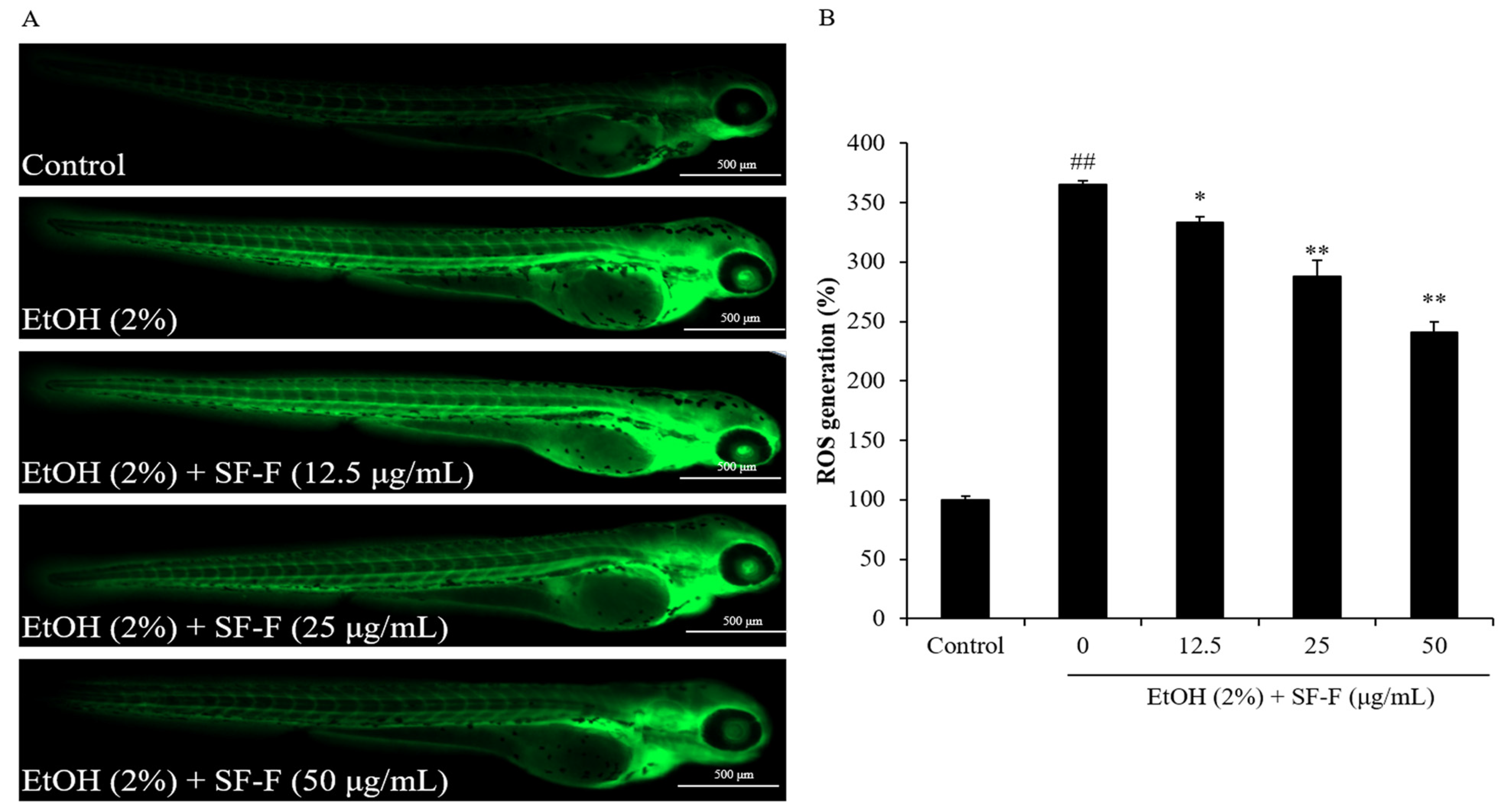
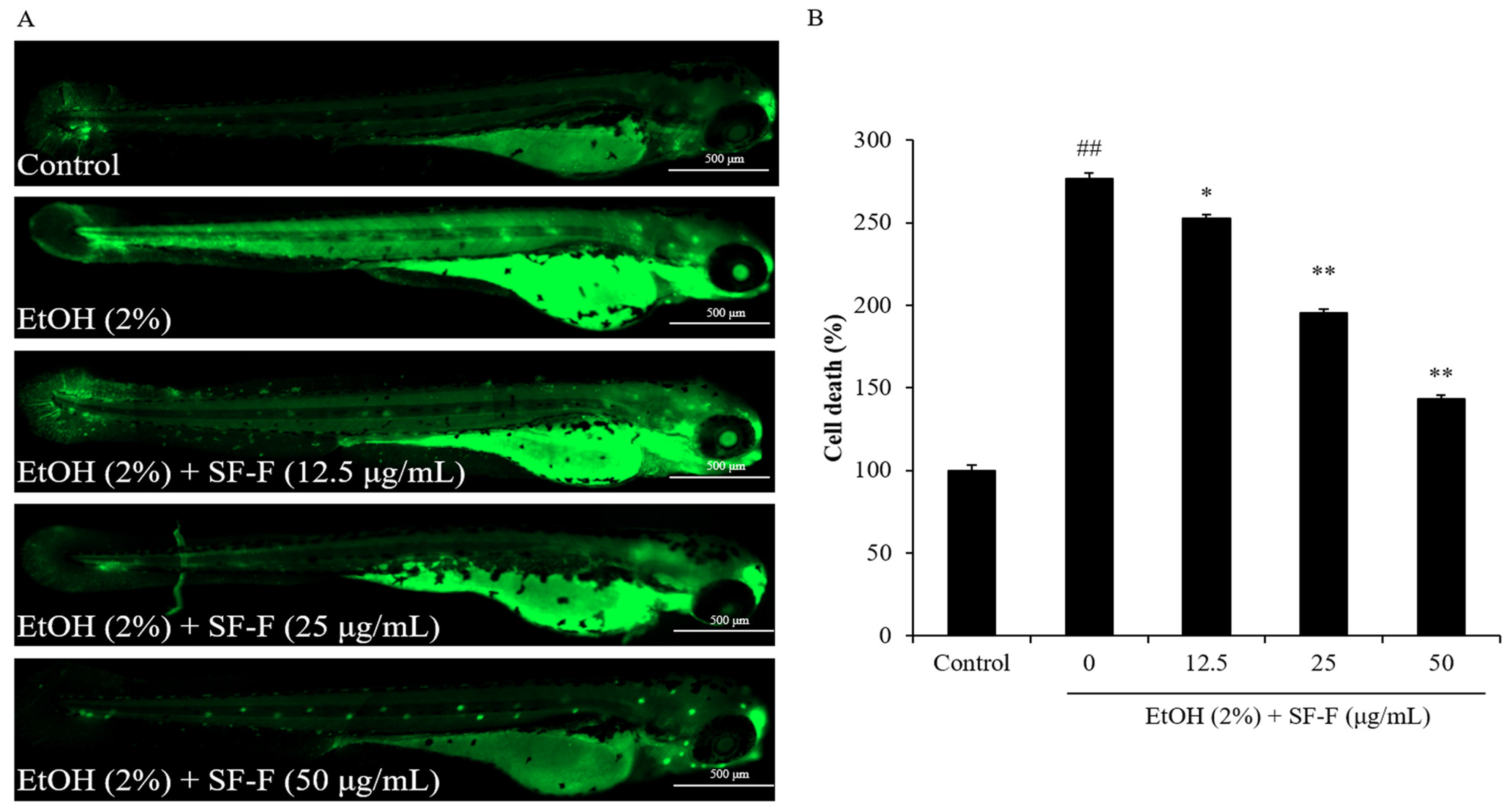
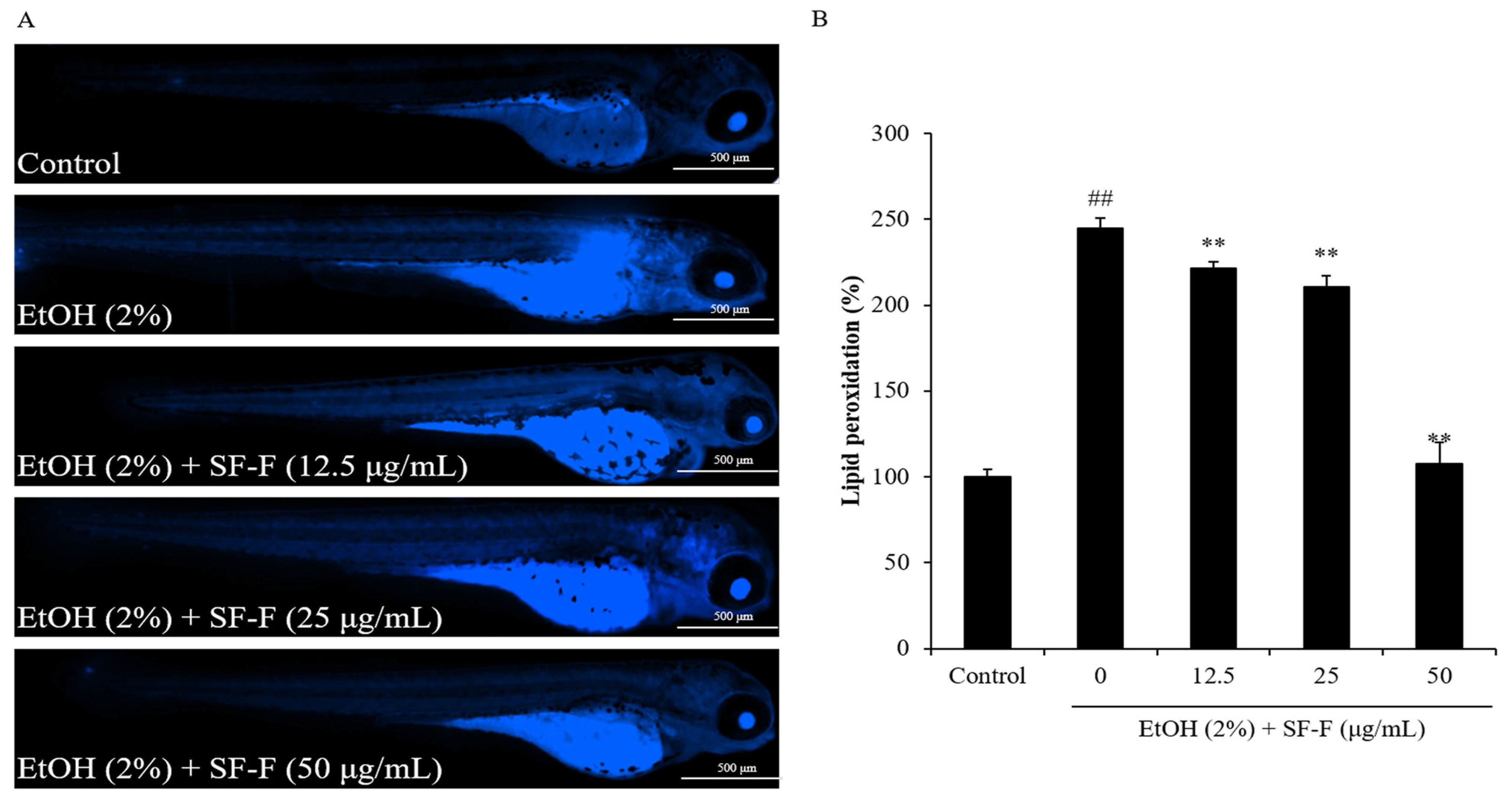
3.2. SF-F Protects Zebrafish against Oxidative Damage Stimulated by EtOH
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, F.; Xu, Y.; You, K.; Zhang, J.; Yang, F.; Li, Y.-X. Calcitriol alleviates ethanol-induced hepatotoxicity via AMPK/mTOR-mediated autophagy. Arch. Biochem. Biophys. 2021, 697, 108694. [Google Scholar] [CrossRef]
- Stephenson, M.; Heron, J.; Bountress, K.; Hickman, M.; Kendler, K.S.; Edwards, A.C. The effect of parental alcohol use on alcohol use disorder in young adulthood: Exploring the mediating roles of adolescent alcohol expectancies and consumption. J. Adolesc. 2023. [Google Scholar] [CrossRef] [PubMed]
- Lippard, E.T.C.; Kirsch, D.E.; Kosted, R.; Le, V.; Almeida, J.R.C.; Fromme, K.; Strakowski, S.M. Subjective response to alcohol in young adults with bipolar disorder and recent alcohol use: A within-subject randomized placebo-controlled alcohol administration study. Psychopharmacology 2023, 240, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Samuvel, D.J.; Li, L.; Krishnasamy, Y.; Gooz, M.; Takemoto, K.; Woster, P.M.; Lemasters, J.J.; Zhong, Z. Mitochondrial depolarization after acute ethanol treatment drives mitophagy in living mice. Autophagy 2022, 18, 2671–2685. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, R.; Wang, X.; Jiang, Y.; Xu, W.; Shao, Y.; Yue, C.; Shi, W.; Jin, H.; Ge, T.; et al. Activation of UQCRC2-dependent mitophagy by tetramethylpyrazine inhibits MLKL-mediated hepatocyte necroptosis in alcoholic liver disease. Free. Radic. Biol. Med. 2022, 179, 301–316. [Google Scholar] [CrossRef]
- Bayrak, B.B.; Arda-Pirincci, P.; Bolkent, S.; Yanardag, R. Zinc Prevents Ethanol-Induced Oxidative Damage in Lingual Tissues of Rats. Biol. Trace Elem. Res. 2022, 200, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Vecchione, G.; Grasselli, E.; Compalati, A.D.; Ragazzoni, M.; Cortese, K.; Gallo, G.; Voci, A.; Vergani, L. Ethanol and fatty acids impair lipid homeostasis in an in vitro model of hepatic steatosis. Food Chem. Toxicol. 2016, 90, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Ma, Y.; Shrivastava, S.K.; Srivastava, R.K.; Shankar, S. Chronic alcohol exposure induces hepatocyte damage by inducing oxidative stress, SATB2 and stem cell-like characteristics, and activating lipogenesis. J. Cell. Mol. Med. 2022, 26, 2119–2131. [Google Scholar] [CrossRef] [PubMed]
- Song, X.-Y.; Li, R.-H.; Liu, W.-W.; Hayashi, T.; Mizuno, K.; Hattori, S.; Fujisaki, H.; Ikejima, T. Effect of silibinin on ethanol- or acetaldehyde-induced damge of mouse primary hepatocytes in vitro. Toxicol. In Vitro 2021, 70, 105047. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chang, X.; Zhan, H.; Zhang, Q.; Li, C.; Gao, Q.; Yang, M.; Luo, Z.; Li, S.; Sun, Y. Curcumin and Baicalin ameliorate ethanol-induced liver oxidative damage via the Nrf2/HO-1 pathway. J. Food Biochem. 2020, 44, e13425. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, R.A.; Pérez, M.J.; Aranguiz, A.; Tapia-Monsalves, C.; Mendez, G. Activation of the Melanocortin-4 Receptor Prevents Oxidative Damage and Mitochondrial Dysfunction in Cultured Hippocampal Neurons Exposed to Ethanol. Neurotox. Res. 2020, 38, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Chen, Y.-H.; Pratama, S.A.; Chen, Y.-L.; Shirakawa, H.; Peng, H.-C.; Yang, S.-C. The Prophylactic Effects of Glutamine on Muscle Protein Synthesis and Degradation in Rats with Ethanol-Induced Liver Damage. Nutrients 2021, 13, 2788. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shang, H.; Li, J.; Ma, Y.; Xu, C.; Ma, J.; Hou, J.; Jiang, Z. Four Different Structural Dietary Polyphenols, Especially Dihydromyricetin, Possess Superior Protective Effect on Ethanol-Induced ICE-6 and AML-12 Cytotoxicity: The Role of CYP2E1 and Keap1-Nrf2 Pathways. J. Agric. Food Chem. 2023, 71, 1518–1530. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Chen, Z.; Wei, H.; Zhu, S.; Cai, K. Isolation and Identification of Antioxidative Peptide from Goose Liver Hydrolysate to Ameliorate Alcohol-Mediated Oxidative Stress Damage in HHL-5 Hepatocytes. Molecules 2022, 27, 7151. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Seaweed-Based Compounds and Products for Sustainable Protection against Plant Pathogens. Mar. Drugs 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Begum, R.; Howlader, S.; Mamun-Or-Rashid, A.N.M.; Rafiquzzaman, S.M.; Ashraf, G.M.; Albadrani, G.M.; Sayed, A.A.; Peluso, I.; Abdel-Daim, M.M.; Uddin, M.S. Antioxidant and Signal-Modulating Effects of Brown Seaweed-Derived Compounds against Oxidative Stress-Associated Pathology. Oxidative Med. Cell. Longev. 2021, 2021, 9974890. [Google Scholar] [CrossRef] [PubMed]
- Kumar, Y.; Tarafdar, A.; Badgujar, P.C. Seaweed as a Source of Natural Antioxidants: Therapeutic Activity and Food Applications. J. Food Qual. 2021, 2021, 5753391. [Google Scholar] [CrossRef]
- Reboleira, J.; Silva, S.; Chatzifragkou, A.; Niranjan, K.; Lemos, M.F.L. Seaweed fermentation within the fields of food and natural products. Trends Food Sci. Technol. 2021, 116, 1056–1073. [Google Scholar] [CrossRef]
- Lomartire, S.; Cotas, J.; Pacheco, D.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Environmental Impact on Seaweed Phenolic Production and Activity: An Important Step for Compound Exploitation. Mar. Drugs 2021, 19, 245. [Google Scholar] [CrossRef]
- Polat, S.; Trif, M.; Rusu, A.; Šimat, V.; Čagalj, M.; Alak, G.; Meral, R.; Özogul, Y.; Polat, A.; Özogul, F. Recent advances in industrial applications of seaweeds. Crit. Rev. Food Sci. Nutr. 2021, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, H.; Wang, X.; Wang, X.; Cong, P.; Xu, J.; Xue, C. Comparative Lipidomics Study of Four Edible Red Seaweeds Based on RPLC–Q-TOF. J. Agric. Food Chem. 2023, 71, 2183–2196. [Google Scholar] [CrossRef]
- Dinesh Kumar, S.; Satish, L.; Dhanya, N.; Malar Vizhi, J.; Nadukkattu Nayagi, N.; Gopala Krishnan, S.; Ganesan, M. Tank cultivation of edible seaweeds: An overview of the Indian perspective for opportunities and challenges. Biomass Convers. Biorefinery 2023. [Google Scholar] [CrossRef]
- Park, E.; Yu, H.; Lim, J.-H.; Hee Choi, J.; Park, K.-J.; Lee, J. Seaweed metabolomics: A review on its nutrients, bioactive compounds and changes in climate change. Food Res. Int. 2023, 163, 112221. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, H.; Waditee-Sirisattha, R. Antioxidative, Anti-Inflammatory, and Anti-Aging Properties of Mycosporine-Like Amino Acids: Molecular and Cellular Mechanisms in the Protection of Skin-Aging. Mar. Drugs 2019, 17, 222. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.-H.; Kim, S.-K. Sulfated polysaccharides as bioactive agents from marine algae. Int. J. Biol. Macromol. 2013, 62, 70–75. [Google Scholar] [CrossRef]
- Raguraman, V.; Jyotsna, J.; Palaniappan, S.; Gopal, S.; Thirugnanasambandam, R.; Kirubagaran, R. Sulfated polysaccharide from Sargassum tenerrimum attenuates oxidative stress induced reactive oxygen species production in in vitro and in zebrafish model. Carbohydr. Polym. 2019, 203, 441–449. [Google Scholar] [CrossRef]
- Jin, W.; Ren, L.; Liu, B.; Zhang, Q.; Zhong, W. Structural Features of Sulfated Glucuronomannan Oligosaccharides and Their Antioxidant Activity. Mar. Drugs 2018, 16, 291. [Google Scholar] [CrossRef]
- Dörschmann, P.; Bittkau, K.S.; Neupane, S.; Roider, J.; Alban, S.; Klettner, A. Effects of Fucoidans from Five Different Brown Algae on Oxidative Stress and VEGF Interference in Ocular Cells. Mar. Drugs 2019, 17, 258. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lu, J.-B.; Wang, C.; Wang, C.-S.; Zhang, H.-H.; Li, C.-Y.; Qian, G.-Y. Effects of Sargassum fusiforme polysaccharides on antioxidant activities and intestinal functions in mice. Int. J. Biol. Macromol. 2013, 58, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Wang, L.; Sanjeewa, K.K.A.; Kang, S.I.; Lee, J.-S.; Jeon, Y.-J. Antioxidant Potential of Sulfated Polysaccharides from Padina boryana; Protective Effect against Oxidative Stress in In Vitro and In Vivo Zebrafish Model. Mar. Drugs 2020, 18, 212. [Google Scholar] [CrossRef]
- Geun Lee, H.; Jayawardena, T.U.; Liyanage, N.M.; Song, K.-M.; Choi, Y.-S.; Jeon, Y.-J.; Kang, M.-C. Antioxidant potential of low molecular weight fucoidans from Sargassum autumnale against H2O2-induced oxidative stress in vitro and in zebrafish models based on molecular weight changes. Food Chem. 2022, 384, 132591. [Google Scholar] [CrossRef]
- Li, X.; Xin, S.; Zheng, X.; Lou, L.; Ye, S.; Li, S.; Wu, Q.; Ding, Q.; Ji, L.; Nan, C.; et al. Inhibition of the Occurrence and Development of Inflammation-Related Colorectal Cancer by Fucoidan Extracted from Sargassum fusiforme. J. Agric. Food Chem. 2022, 70, 9463–9476. [Google Scholar] [CrossRef]
- Zuo, J.; Zhang, Y.; Wu, Y.; Liu, J.; Wu, Q.; Shen, Y.; Jin, L.; Wu, M.; Ma, Z.; Tong, H. Sargassum fusiforme fucoidan ameliorates diet-induced obesity through enhancing thermogenesis of adipose tissues and modulating gut microbiota. Int. J. Biol. Macromol. 2022, 216, 728–740. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.-Y.; Yang, H.-W.; Fu, X.; Kim, J.-I.; Jeon, Y.-J. Fucoidan isolated from the popular edible brown seaweed Sargassum fusiforme suppresses lipopolysaccharide-induced inflammation by blocking NF-κB signal pathway. J. Appl. Phycol. 2021, 33, 1845–1852. [Google Scholar] [CrossRef]
- Wang, L.; Jayawardena, T.U.; Yang, H.-W.; Lee, H.G.; Kang, M.-C.; Sanjeewa, K.K.A.; Oh, J.Y.; Jeon, Y.-J. Isolation, Characterization, and Antioxidant Activity Evaluation of a Fucoidan from an Enzymatic Digest of the Edible Seaweed, Hizikia fusiforme. Antioxidants 2020, 9, 363. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-C.; Kim, K.-N.; Wijesinghe, W.A.J.P.; Yang, X.; Ahn, G.; Jeon, Y.-J. Protective effect of polyphenol extracted from Ecklonia cava against ethanol induced oxidative damage in vitro and in zebrafish model. J. Funct. Foods 2014, 6, 339–347. [Google Scholar] [CrossRef]
- Chen, J.; Wang, D.; Zong, Y.; Yang, X. DHA Protects Hepatocytes from Oxidative Injury through GPR120/ERK-Mediated Mitophagy. Int. J. Mol. Sci. 2021, 22, 5675. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, E.A.; Chen, C.Y.; Wu, H.N.; Chien, C.-C.; Chen, S.-C. Propolis alleviates 4-aminobiphenyl-induced oxidative DNA damage by inhibition of CYP2E1 expression in human liver cells. Environ. Toxicol. 2021, 36, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.-Y.; Chen, L.; Li, Q.-J.; Shen, Y.-Z.; Jin, L.; Zhang, X.; Chen, P.-C.; Wu, M.-J.; Choi, J.-I.; et al. Different extraction methods bring about distinct physicochemical properties and antioxidant activities of Sargassum fusiforme fucoidans. Int. J. Biol. Macromol. 2020, 155, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhang, X.; Tang, Y.; Mao, J. Composition, isolation, purification and biological activities of Sargassum fusiforme polysaccharides: A review. Carbohydr. Polym. 2020, 228, 115381. [Google Scholar] [CrossRef]
- Baggio, S.; Zenki, K.; Martins Silva, A.; dos Santos, T.G.; Rech, G.; Lazzarotto, G.; Dias, R.D.; Mussulini, B.H.; Rico, E.P.; de Oliveira, D.L. Fetal alcohol spectrum disorders model alters the functionality of glutamatergic neurotransmission in adult zebrafish. NeuroToxicology 2020, 78, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro-da-Silva, J.; Luchiari, A.C. Embryonic ethanol exposure on zebrafish early development. Brain Behav. 2021, 11, e02062. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Oh, J.-Y.; Yang, H.-W.; Hyun, J.; Ahn, G.; Fu, X.; Xu, J.; Gao, X.; Cha, S.-H.; Jeon, Y.-J. Protective Effect of Sargassum fusiforme Fucoidan against Ethanol-Induced Oxidative Damage in In Vitro and In Vivo Models. Polymers 2023, 15, 1912. https://doi.org/10.3390/polym15081912
Wang L, Oh J-Y, Yang H-W, Hyun J, Ahn G, Fu X, Xu J, Gao X, Cha S-H, Jeon Y-J. Protective Effect of Sargassum fusiforme Fucoidan against Ethanol-Induced Oxidative Damage in In Vitro and In Vivo Models. Polymers. 2023; 15(8):1912. https://doi.org/10.3390/polym15081912
Chicago/Turabian StyleWang, Lei, Jae-Young Oh, Hye-Won Yang, Jimin Hyun, Ginnae Ahn, Xiaoting Fu, Jiachao Xu, Xin Gao, Seon-Heui Cha, and You-Jin Jeon. 2023. "Protective Effect of Sargassum fusiforme Fucoidan against Ethanol-Induced Oxidative Damage in In Vitro and In Vivo Models" Polymers 15, no. 8: 1912. https://doi.org/10.3390/polym15081912