
Immobilization of Hyperthermostable Carboxylesterase EstD9 from Anoxybacillus geothermalis D9 onto Polymer Material and Its Physicochemical Properties
 , and
, and 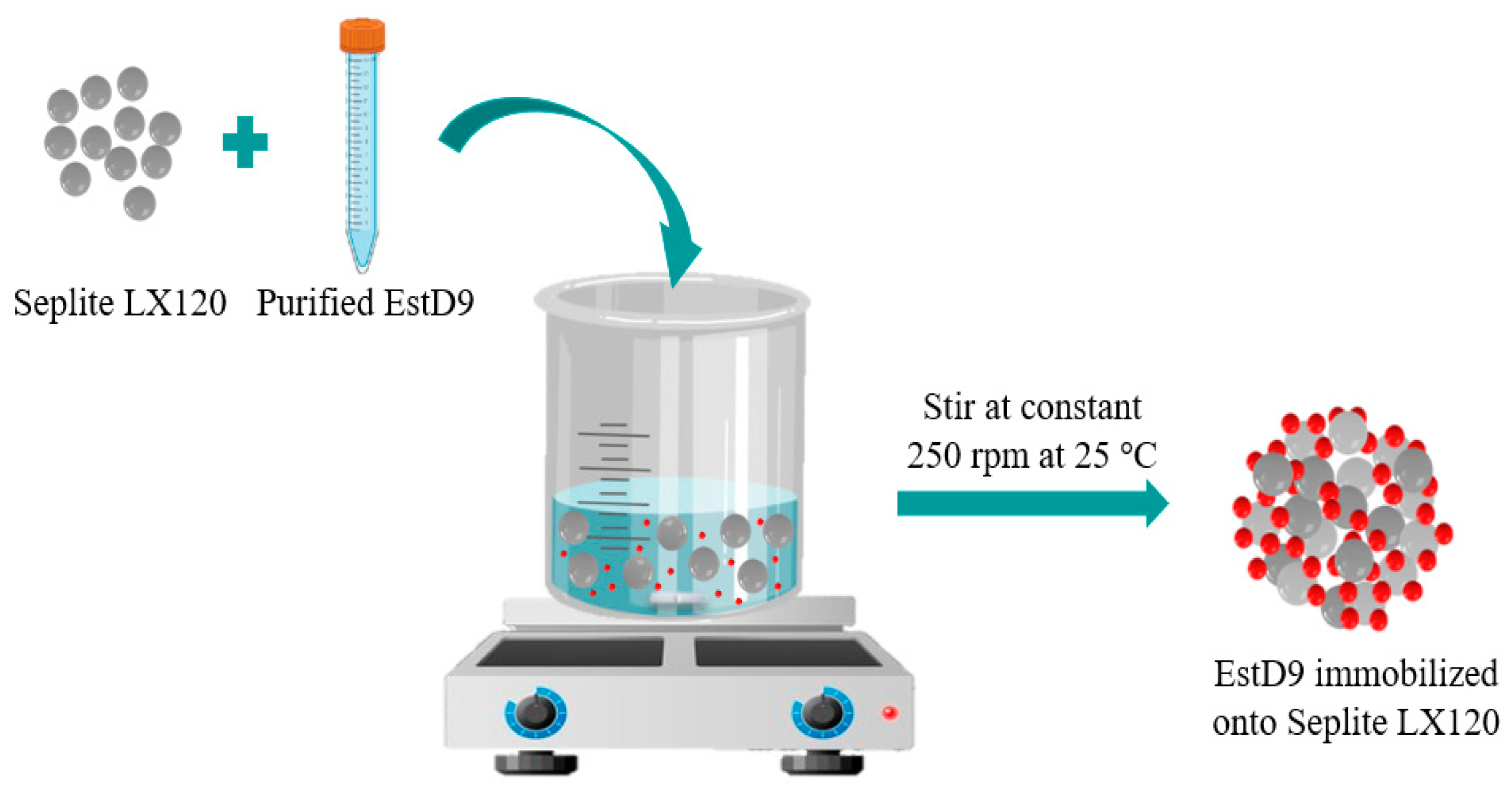{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Enzyme Preparation
2.3. Protein Content Determination
2.4. Immobilization of EstD9
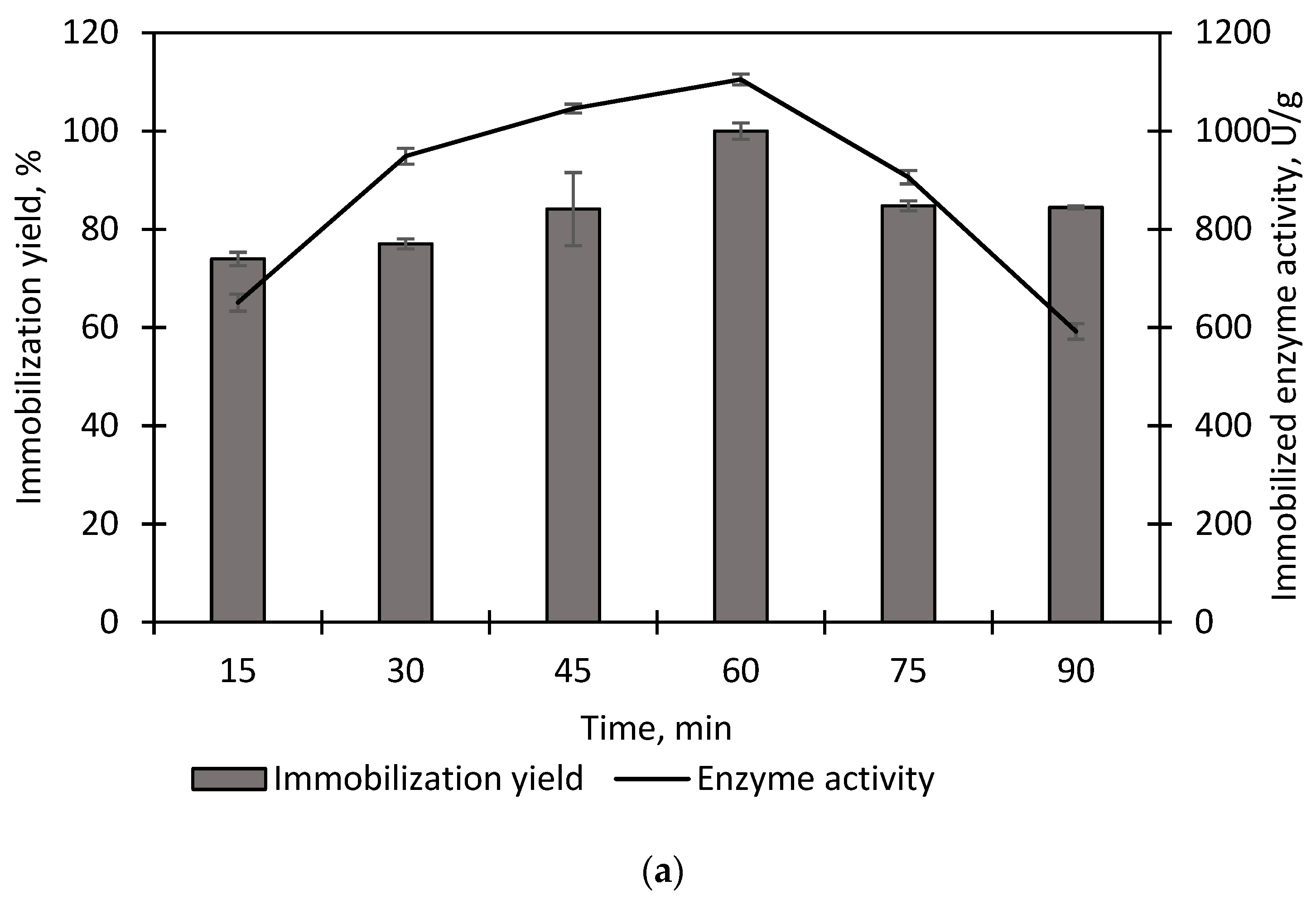
2.4.1. Effect of Adsorption Time
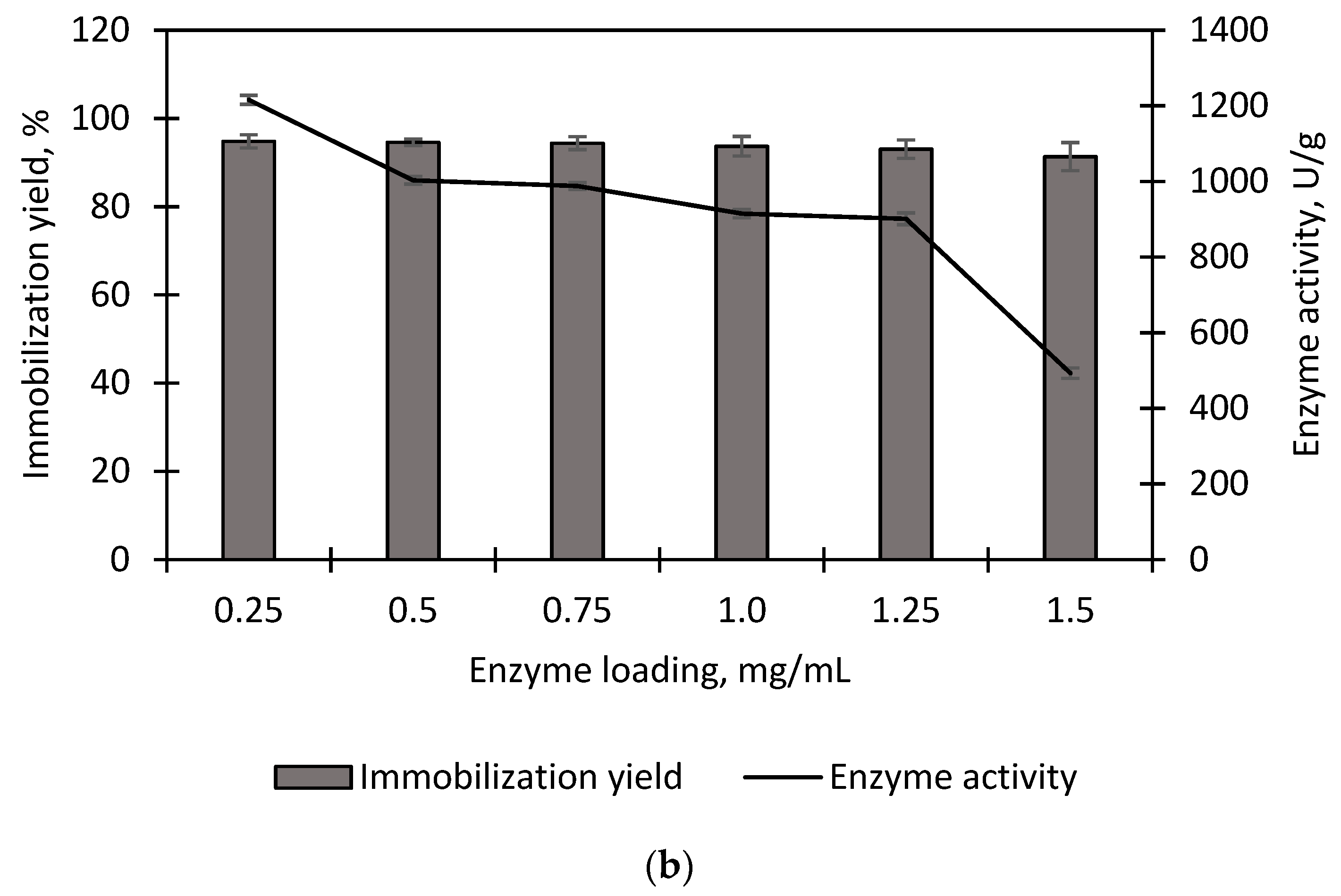
2.4.2. Effect of Enzyme Loading
2.5. Enzyme Assay
2.6. Structural and Morphological Characterization
2.7. Biochemical Characterization
2.7.1. Effect of Temperature on Activity and Stability
2.7.2. Effect of pH on Activity and Stability
2.7.3. Effect of Organic Solvents on Activity
2.7.4. Effect of Metal Ions on Activity
2.8. Storage Stability
2.9. Reusability
2.10. Statistical Analysis
3. Results and Discussion
3.1. Immobilization of Purified EstD9
3.2. Morphological Characterization of Immobilized EstD9
3.3. BET Analysis of Seplite LX120 and Immobilized EstD9
3.4. Structural Analysis of Immobilized EstD9
3.5. Effect of Temperature on Immobilized EstD9 Activity and Stability
3.6. Effect of pH on Activity and Stability of Immobilized EstD9
3.7. Effect of Organic Solvents on Activity of Immobilized EstD9
3.8. Effect of Metal Ions on Activity of Immobilized EstD9
3.9. Storage Stability and Reusability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bonzom, C.; Schild, L.; Gustafsson, H.; Olsson, L. Feruloyl esterase immobilization in mesoporous silica particles and characterization in hydrolysis and transesterification. BMC Biochem. 2018, 19, 1. [Google Scholar] [CrossRef] [Green Version]
- Lan, L.; Ren, X.; Yang, J.; Liu, D.; Zhang, C. Detection techniques of carboxylesterase activity: An update review. Bioorg. Chem. 2020, 94, 103388. [Google Scholar] [CrossRef] [PubMed]
- Abeijón Mukdsi, M.C.; Maillard, M.B.; Medina, R.B.; Thierry, A. Ethyl butanoate is synthesised both by alcoholysis and esterification by dairy lactobacilli and propionibacteria. LWT 2018, 89, 38–43. [Google Scholar] [CrossRef]
- Johan, U.U.M.; Rahman, R.N.Z.R.A.; Kamarudin, N.H.A.; Ali, M.S.M. An integrated overview of bacterial carboxylesterase: Structure, function and biocatalytic applications. Colloids Surf. B Biointerfaces 2021, 205, 111882. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Rahman, Z.; Mubeen, H.; Makhdoom, J.; Tariq, J.; Mahjabeen, N.; Ali, Z.; Hamid, A.; Shafique, E.; Aftab, M.N. Heterologous expression, molecular studies and biochemical characterization of a novel alkaline esterase gene from Bacillus thuringiensis for detergent industry. RSC Adv. 2022, 12, 34482–34495. [Google Scholar] [CrossRef]
- Kumar, D.; Bhardwaj, R.; Jassal, S.; Goyal, T.; Khullar, A.; Gupta, N. Application of enzymes for an eco-friendly approach to textile processing. Environ. Sci. Pollut. Res. 2021; Online ahead of print. [Google Scholar] [CrossRef]
- Barzkar, N.; Sohail, M.; Tamadoni Jahromi, S.; Gozari, M.; Poormozaffar, S.; Nahavandi, R.; Hafezieh, M. Marine Bacterial Esterases: Emerging Biocatalysts for Industrial Applications. Appl. Biochem. Biotechnol. 2021, 193, 1187–1214. [Google Scholar] [CrossRef]
- Dahiya, D.; Nigam, P.S. Sustainable Biosynthesis of Esterase Enzymes of Desired Characteristics of Catalysis for Pharmaceutical and Food Industry Employing Specific Strains of Microorganisms. Sustainability 2022, 14, 8673. [Google Scholar] [CrossRef]
- Beladiya, C.; Tripathy, R.K.; Bajaj, P.; Aggarwal, G.; Pande, A.H. Expression, purification and immobilization of recombinant AiiA enzyme onto magnetic nanoparticles. Protein Expr. Purif. 2015, 113, 56–62. [Google Scholar] [CrossRef]
- Basso, A.; Serban, S. Industrial applications of immobilized enzymes—A review. Mol. Catal. 2019, 479, 110607. [Google Scholar] [CrossRef]
- Liang, S.; Wu, X.L.; Xiong, J.; Zong, M.H.; Lou, W.Y. Metal-organic frameworks as novel matrices for efficient enzyme immobilization: An update review. Coord. Chem. Rev. 2020, 406, 213149. [Google Scholar] [CrossRef]
- Bayazidi, P.; Almasi, H.; Asl, A.K. Immobilization of lysozyme on bacterial cellulose nanofibers: Characteristics, antimicrobial activity and morphological properties. Int. J. Biol. Macromol. 2018, 107, 2544–2551. [Google Scholar] [CrossRef] [PubMed]
- Bilal, M.; Zhao, Y.; Rasheed, T.; Iqbal, H.M.N. Magnetic nanoparticles as versatile carriers for enzymes immobilization: A review. Int. J. Biol. Macromol. 2018, 120, 2530–2544. [Google Scholar] [CrossRef] [PubMed]
- Remonatto, D.; Miotti, R.H.; Monti, R.; Bassan, J.C.; de Paula, A.V. Applications of immobilized lipases in enzymatic reactors: A review. Process Biochem. 2022, 114, 1–20. [Google Scholar] [CrossRef]
- Liu, D.M.; Chen, J.; Shi, Y.P. Advances on methods and easy separated support materials for enzymes immobilization. TrAC Trends Anal. Chem. 2018, 102, 332–342. [Google Scholar] [CrossRef]
- Darwesh, O.M.; Ali, S.S.; Matter, I.A.; Elsamahy, T.; Mahmoud, Y.A. Enzymes Immobilization onto Magnetic Nanoparticles to Improve Industrial and Environmental Applications, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 630, ISBN 9780128201435. [Google Scholar]
- Le, L.T.H.L.; Yoo, W.; Jeon, S.; Kim, K.K.; Kim, T.D. Characterization and Immobilization of a Novel SGNH Family Esterase (LaSGNH1) from Lactobacillus acidophilus NCFM. Int. J. Mol. Sci. 2019, 21, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana-Peña, S.; Carballares, D.; Morellon-Sterling, R.; Rocha-Martin, J.; Fernandez-Lafuente, R. The combination of covalent and ionic exchange immobilizations enables the coimmobilization on vinyl sulfone activated supports and the reuse of the most stable immobilized enzyme. Int. J. Biol. Macromol. 2022, 199, 51–60. [Google Scholar] [CrossRef]
- Yushkova, E.D.; Nazarova, E.A.; Matyuhina, A.V.; Noskova, A.O.; Shavronskaya, D.O.; Vinogradov, V.V.; Skvortsova, N.N.; Krivoshapkina, E.F. Application of Immobilized Enzymes in Food Industry. J. Agric. Food Chem. 2019, 67, 11553–11567. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, V.; Tonetto, G.; Ferreira, M.L. Optimization of the enzymatic synthesis of pentyl oleate with lipase immobilized onto novel structured support. Fermentation 2019, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Feng, Y.; Wang, G.; Wang, Z.; Bilal, M.; Lv, H.; Jia, S.; Cui, J. Production and use of immobilized lipases in/on nanomaterials: A review from the waste to biodiesel production. Int. J. Biol. Macromol. 2020, 152, 207–222. [Google Scholar] [CrossRef]
- Somu, P.; Narayanasamy, S.; Gomez, L.A.; Rajendran, S.; Lee, Y.R.; Balakrishnan, D. Immobilization of enzymes for bioremediation: A future remedial and mitigating strategy. Environ. Res. 2022, 212, 113411. [Google Scholar] [CrossRef]
- Almeida, R.V.; Branco, R.V.; Peixoto, B.; da Silva Lima, C.; Alqueres, S.M.C.; Martins, O.B.; Antunes, O.A.C.; Freire, D.M.G. Immobilization of a recombinant thermostable esterase (Pf2001) from Pyrococcus furiosus on microporous polypropylene: Isotherms, hyperactivation and purification. Biochem. Eng. J. 2008, 39, 531–537. [Google Scholar] [CrossRef]
- Öz, Y.; Sürmeli, Y.; Sanli-Mohamed, G. Immobilization of Thermoalkalophilic Esterase Onto Magnetic-Cornstarch Nanoparticle. Res. Sq. 2021, 1–20. [Google Scholar] [CrossRef]
- Bezerra, C.S.; De Farias Lemos, C.M.G.; De Sousa, M.; Gonçalves, L.R.B. Enzyme immobilization onto renewable polymeric matrixes: Past, present, and future trends. J. Appl. Polym. Sci. 2015, 132, 1–15. [Google Scholar] [CrossRef]
- Lyu, X.; Gonzalez, R.; Horton, A.; Li, T. Immobilization of enzymes by polymeric materials. Catalysts 2021, 11, 1211. [Google Scholar] [CrossRef]
- Cantone, S.; Ferrario, V.; Corici, L.; Ebert, C.; Fattor, D.; Spizzo, P.; Gardossi, L. Efficient immobilisation of industrial biocatalysts: Criteria and constraints for the selection of organic polymeric carriers and immobilisation methods. Chem. Soc. Rev. 2013, 42, 6262–6276. [Google Scholar] [CrossRef] [Green Version]
- Johan, U.U.M.; Rahman, R.N.Z.R.A.; Kamarudin, N.H.A.; Latip, W.; Ali, M.S.M. A new hyper-thermostable carboxylesterase from Anoxybacillus geothermalis D9. Int. J. Biol. Macromol. 2022, 222, 2486–2497. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sheldon, R.A. Enzyme immobilization: The quest for optimum performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Liu, X.; Guan, Y.; Shen, R.; Liu, H. Immobilization of lipase onto micron-size magnetic beads. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 822, 91–97. [Google Scholar] [CrossRef]
- Tamayo-Cabezas, J.; Karboune, S. Optimizing Immobilization and Stabilization of Feruloyl Esterase from Humicola Insolens and its Application for the Feruloylation of Oligosaccharides. Process Biochem. 2020, 98, 11–20. [Google Scholar] [CrossRef]
- Yang, X.; Xudong, T.; Fengying, D.; Lin, L.; Wei, W.; Wei, D. Facile one-pot immobilization of a novel thermostable carboxylesterase from Geobacillus uzenensis for continuous pesticide degradation in a packed-bed column reactor. Catalysts 2020, 10, 518. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Peirce, S.; Torrestiana-Sanchez, B.; Yates, M.; Rosales-Quintero, A.; Virgen-Ortíz, J.J.; Fernandez-Lafuente, R. Evaluation of different commercial hydrophobic supports for the immobilization of lipases: Tuning their stability, activity and specificity. RSC Adv. 2016, 6, 100281–100294. [Google Scholar] [CrossRef]
- Tercan, Ç.; Sürmeli, Y.; Şanlı-Mohamed, G. Thermoalkalophilic recombinant esterase entrapment in chitosan/calcium/alginate-blended beads and its characterization. J. Chem. Technol. Biotechnol. 2021, 96, 2257–2264. [Google Scholar] [CrossRef]
- Bardestani, R.; Patience, G.S.; Kaliaguine, S. Experimental methods in chemical engineering: Specific surface area and pore size distribution measurements—BET, BJH, and DFT. Can. J. Chem. Eng. 2019, 97, 2781–2791. [Google Scholar] [CrossRef]
- Sadat, A.; Joye, I.J. Peak fitting applied to fourier transform infrared and raman spectroscopic analysis of proteins. Appl. Sci. 2020, 10, 5918. [Google Scholar] [CrossRef]
- Shao, P.; He, Z.; Hu, Y.; Shen, Y.; Zhang, S.; Yu, Y. Zeolitic imidazolate frameworks with different organic ligands as carriers for Carbonic anhydrase immobilization to promote the absorption of CO2 into tertiary amine solution. Chem. Eng. J. 2022, 435, 134957. [Google Scholar] [CrossRef]
- Pota, G.; Sapienza Salerno, A.; Costantini, A.; Silvestri, B.; Passaro, J.; Califano, V. Co-immobilization of Cellulase and β-Glucosidase into Mesoporous Silica Nanoparticles for the Hydrolysis of Cellulose Extracted from Eriobotrya japonica Leaves. Langmuir 2022, 38, 5481–5493. [Google Scholar] [CrossRef]
- Zahirinejad, S.; Hemmati, R.; Homaei, A.; Dinari, A.; Hosseinkhani, S.; Mohammadi, S.; Vianello, F. Nano-organic supports for enzyme immobilization: Scopes and perspectives. Colloids Surf. B Biointerfaces 2021, 204, 111774. [Google Scholar] [CrossRef]
- He, S.; Wu, X.; Ma, B.; Xu, Y. High specific immobilization of His-tagged recombinant Microbacterium esterase by Ni-NTA magnetic chitosan microspheres for efficient synthesis of key chiral intermediate of d-biotin. Bioprocess Biosyst. Eng. 2021, 44, 2193–2204. [Google Scholar] [CrossRef]
- Jun, L.Y.; Yon, L.S.; Mubarak, N.M.; Bing, C.H.; Pan, S.; Danquah, M.K.; Abdullah, E.C.; Khalid, M. An overview of immobilized enzyme technologies for dye and phenolic removal from wastewater. J. Environ. Chem. Eng. 2019, 7, 102961. [Google Scholar] [CrossRef]
- Kim, J.K.; Abdelhamid, M.A.A.; Pack, S.P. Direct immobilization and recovery of recombinant proteins from cell lysates by using EctP1-peptide as a short fusion tag for silica and titania supports. Int. J. Biol. Macromol. 2019, 135, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, K.K.; Kishen, S.; Mehta, A.; Sharma, A.; Gupta, R. Purification of high molecular weight thermotolerant esterase from Serratia sp. and its characterization. 3 Biotech 2021, 11, 308. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Hui-Ying, Y. Purification and characterization of an extracellular esterase with organic solvent tolerance from a halotolerant isolate, Salimicrobium sp. LY19. BMC Biotechnol. 2013, 13, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Dhar, K.; Kanwar, S.S.; Arora, P.K. Lipase catalysis in organic solvents: Advantages and applications. Biol. Proced. Online 2016, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divakar, K.; Suryia Prabha, M.; Pennathur, G. Purification, immobilization and kinetic characterization of G-x-S-x-G esterase with short chain fatty acid specificity from Lysinibacillus fusiformis AU01. Biocatal. Agric. Biotechnol. 2017, 12, 131–141. [Google Scholar] [CrossRef]
- Exterkate, F.A. Structural changes and interactions involved in the Ca2+-triggered stabilization of the cell-bound cell envelope proteinase in Lactococcus lactis subsp. cremoris SK11. Appl. Environ. Microbiol. 2000, 66, 2021–2028. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.X.; Zhang, Y.; Guo, Z.Q.; Zhao, B.; Guo, J.S. Role of Ca2+ and Mg2+ in changing biofilm structure and enhancing biofilm formation of P. stutzeri strain XL-2. Colloids Surf. B Biointerfaces 2022, 220, 112972. [Google Scholar] [CrossRef]
- Wu, P.; Luo, F.; Lu, Z.; Zhan, Z.; Zhang, G. Improving the Catalytic Performance of Pectate Lyase Through Pectate Lyase/Cu3(PO4)2 Hybrid Nanoflowers as an Immobilized Enzyme. Front. Bioeng. Biotechnol. 2020, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bilal, M.; Iqbal, H.M.N. Naturally-derived biopolymers: Potential platforms for enzyme immobilization. Int. J. Biol. Macromol. 2019, 130, 462–482. [Google Scholar] [CrossRef]
- Aghaei, H.; Mohammadbagheri, Z.; Hemasi, A.; Taghizadeh, A. Efficient hydrolysis of starch by α-amylase immobilized on cloisite 30B and modified forms of cloisite 30B by adsorption and covalent methods. Food Chem. 2022, 373, 131425. [Google Scholar] [CrossRef]
- Rahman, M.A.; Culsum, U.; Kumar, A.; Gao, H.; Hu, N. Immobilization of a novel cold active esterase onto Fe3O4~cellulose nano-composite enhances catalytic properties. Int. J. Biol. Macromol. 2016, 87, 488–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolling, D.J.; Suguino, W.A.; Angonesi Brod, F.C.; Arisi, A.C.M. Immobilization of a recombinant esterase from Lactobacillus plantarum on polypropylene Accurel MP1000. Appl. Biochem. Biotechnol. 2011, 163, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Lian, Z.X.; Ma, Z.S.; Wei, J.; Liu, H. Preparation and characterization of immobilized lysozyme and evaluation of its application in edible coatings. Process Biochem. 2012, 47, 201–208. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johan, U.U.M.; Rahman, R.N.Z.R.A.; Kamarudin, N.H.A.; Latip, W.; Ali, M.S.M. Immobilization of Hyperthermostable Carboxylesterase EstD9 from Anoxybacillus geothermalis D9 onto Polymer Material and Its Physicochemical Properties. Polymers 2023, 15, 1361. https://doi.org/10.3390/polym15061361
Johan UUM, Rahman RNZRA, Kamarudin NHA, Latip W, Ali MSM. Immobilization of Hyperthermostable Carboxylesterase EstD9 from Anoxybacillus geothermalis D9 onto Polymer Material and Its Physicochemical Properties. Polymers. 2023; 15(6):1361. https://doi.org/10.3390/polym15061361
Chicago/Turabian StyleJohan, Ummie Umaiera Mohd., Raja Noor Zaliha Raja Abd. Rahman, Nor Hafizah Ahmad Kamarudin, Wahhida Latip, and Mohd Shukuri Mohamad Ali. 2023. "Immobilization of Hyperthermostable Carboxylesterase EstD9 from Anoxybacillus geothermalis D9 onto Polymer Material and Its Physicochemical Properties" Polymers 15, no. 6: 1361. https://doi.org/10.3390/polym15061361