
Harvesting Mycelial Biomass of Selected Basidiomycetes for Chitosan Biopolymer Extraction
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Material
2.2. Inoculum Preparation
2.3. Biomass Cultivation
2.4. Chitosan Extraction
2.5. Nitrogen Content
2.6. Fourier Transform Infrared Spectrometry (FTIR)
2.7. Statistical Analysis
3. Results and Discussion
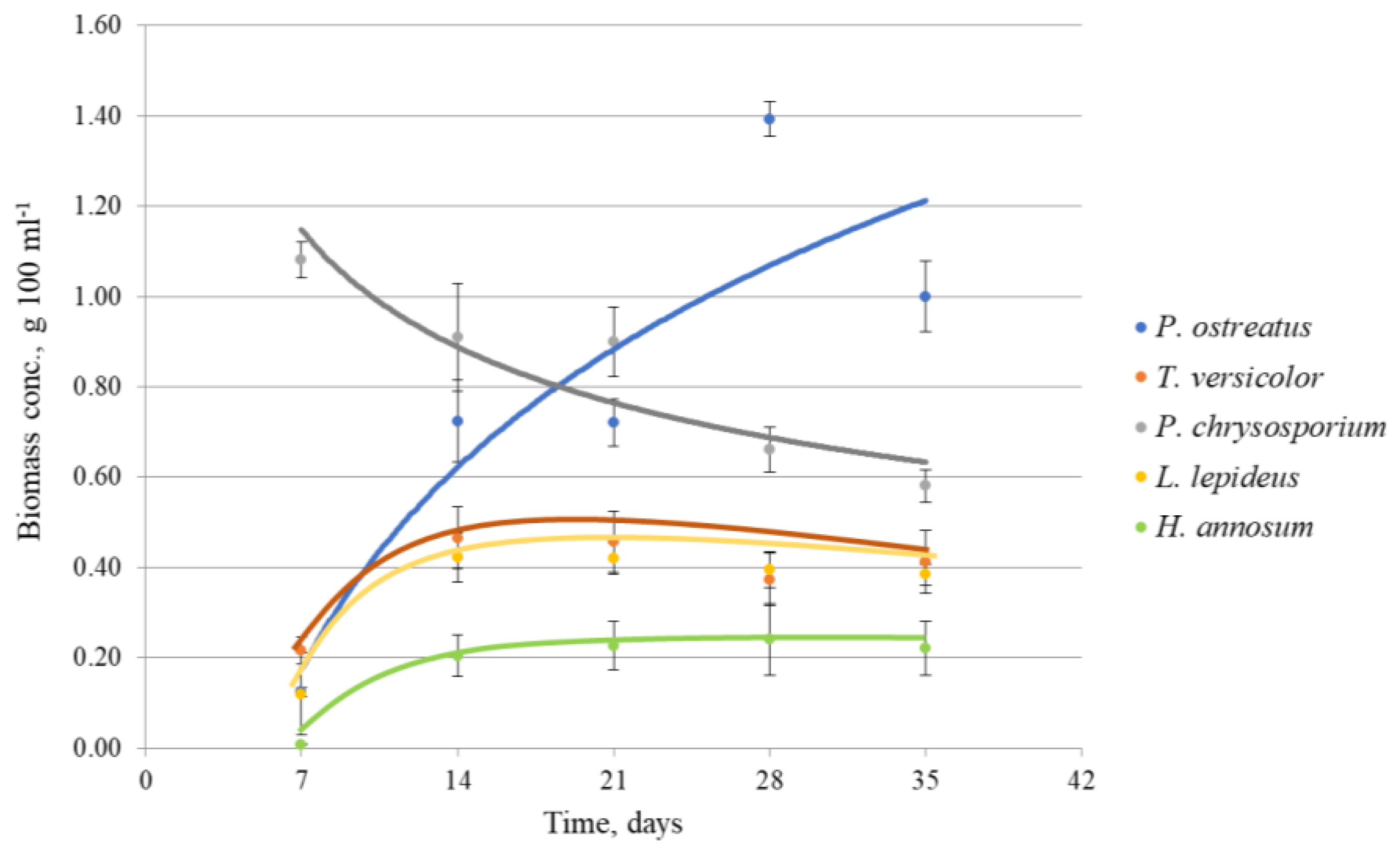
3.1. Growth Dynamics
3.2. Biomass Concentration
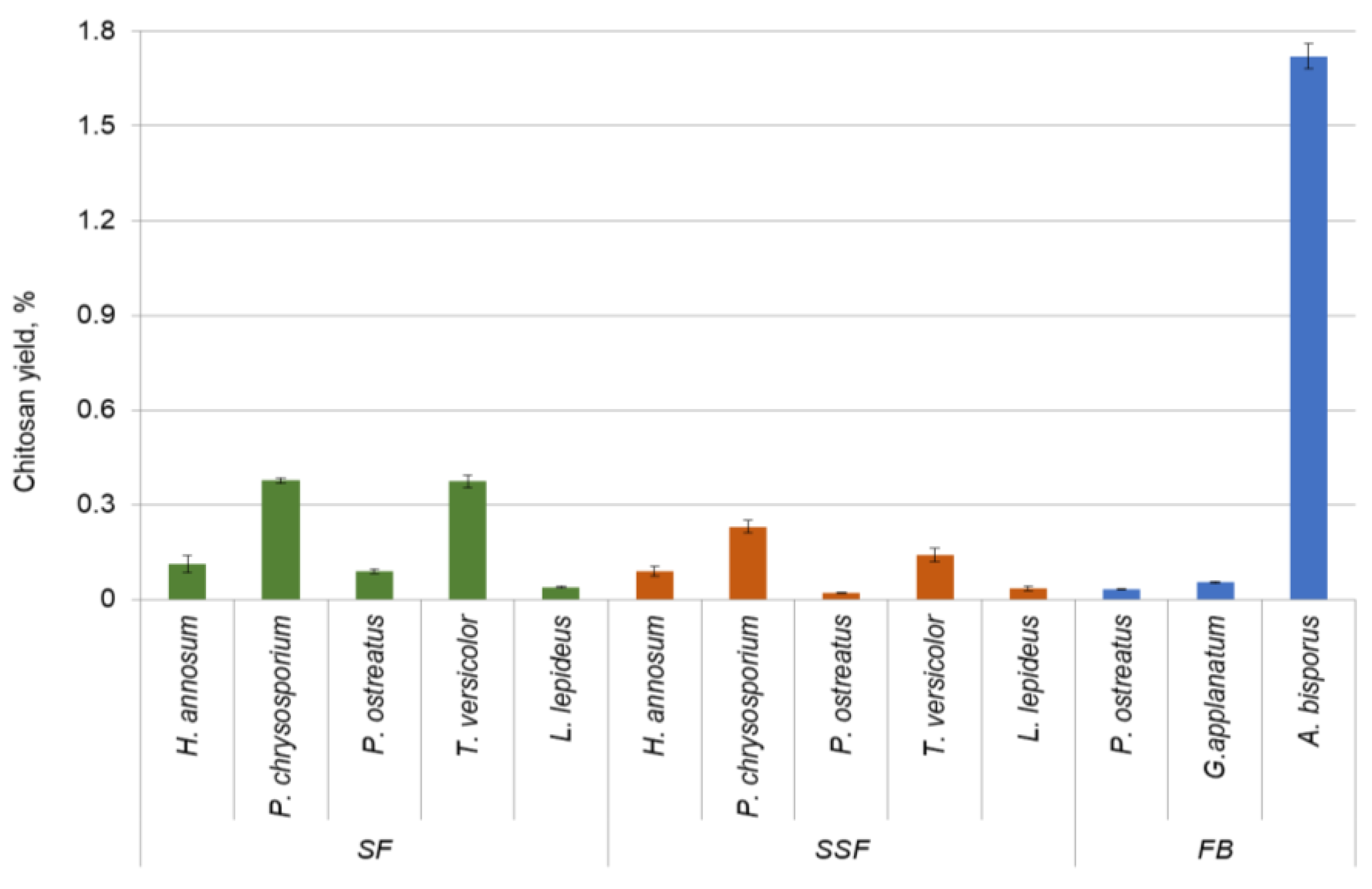
3.3. Chitin-Containing Part and Chitosan Yield
3.4. Nitrogen Content in Fungal Material
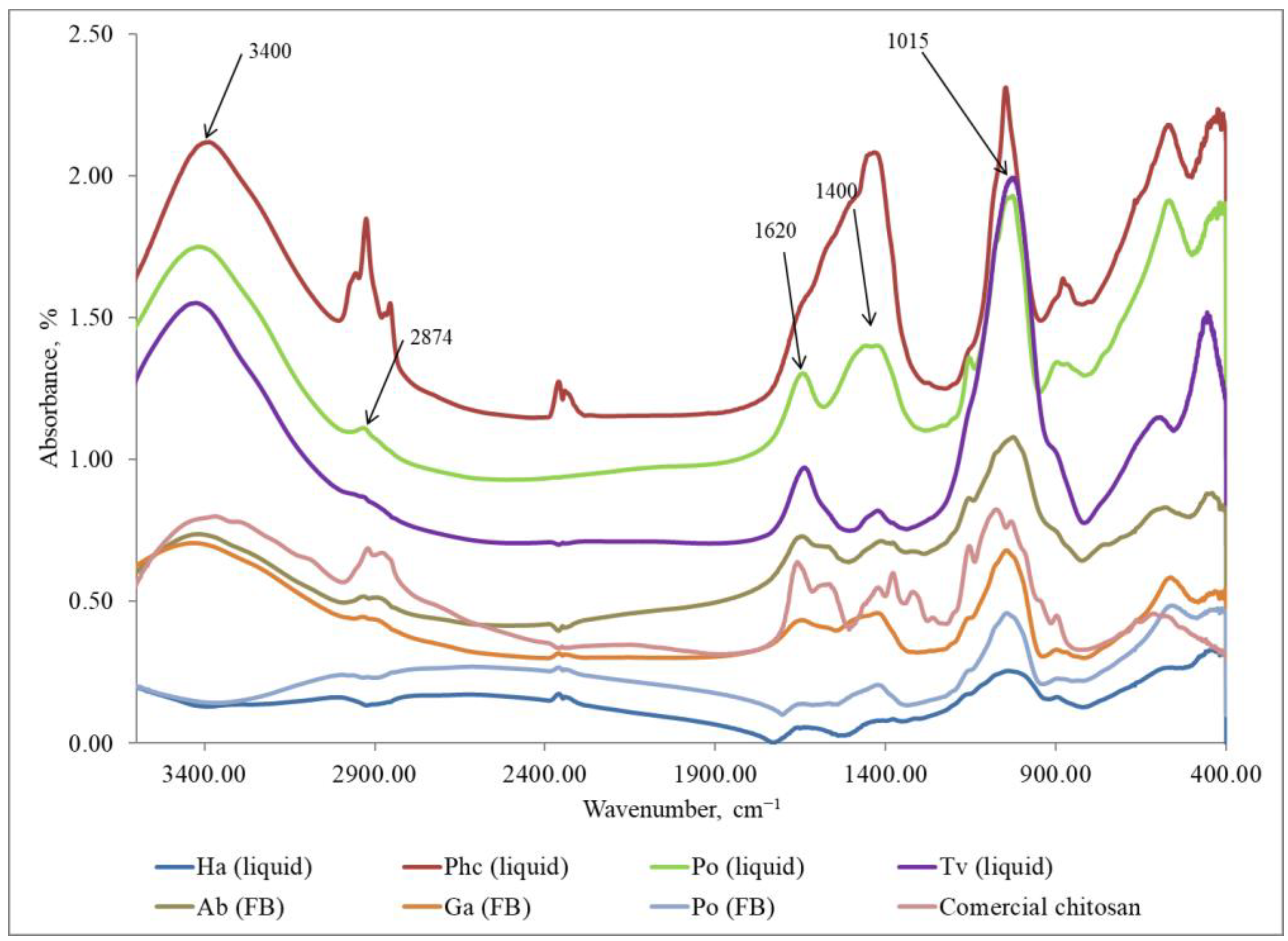
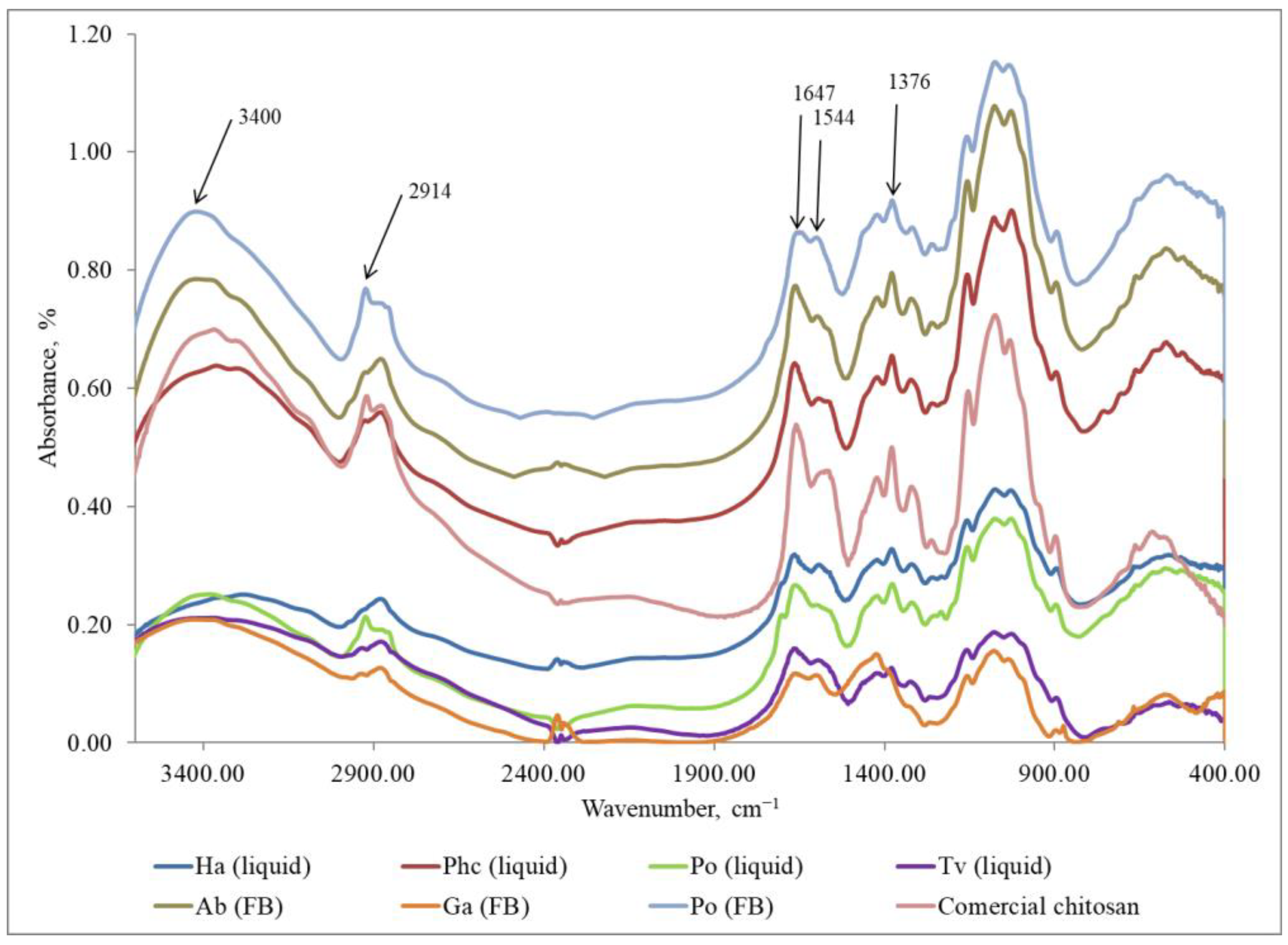
3.5. Characterization of Fungal Chitosan by Fourier Transform Infrared Spectra
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gow Neil, A.R.; Latge, J.-P.; Munro Carol, A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol. Spectr. 2017, 5, 10–1128. [Google Scholar] [CrossRef]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology: Decay and Its Prevention; Academic Press: Cambridge, MA, USA, 2020; 576p. [Google Scholar]
- Abo Elsoud, M.M.; El Kady, E.M. Current trends in fungal biosynthesis of chitin and chitosan. Bull. Natl. Res. Cent. 2019, 43, 59. [Google Scholar] [CrossRef]
- Brown, H.E.; Esher, S.K.; Alspaugh, J.A. Chitin: A “Hidden Figure” in the Fungal Cell Wall. In The Fungal Cell Wall: An Armour and a Weapon for Human Fungal Pathogens; Latgé, J.-P., Ed.; Springer International Publishing: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008. [Google Scholar]
- Pellis, A.; Guebitz, G.M.; Nyanhongo, G.S. Chitosan: Sources, Processing and Modification Techniques. Gels 2022, 8, 393. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, I.; Alcántara, A.R.; Civera, M.C.; Arias, C.; Elorza, B.; Heras Caballero, A.; Acosta, N. Chitosan: An Overview of Its Properties and Applications. Polymers 2021, 13, 3256. [Google Scholar] [CrossRef]
- Singh, R.; Upadhyay, S.K.; Singh, M.; Sharma, I.; Sharma, P.; Kamboj, P.; Saini, A.K.; Voraha, R.; Sharma, A.; Upadhyay, T.K.; et al. Chitin, Chitinases and Chitin Derivatives in Biopharmaceutical, Agricultural and Environmental Perspective. Biointerface Res. Appl. Chem. 2022, 11, 9985–10005. [Google Scholar] [CrossRef]
- Gaitnieks, T.; Bruna, L.; Zaluma, A.; Burnevica, N.; Klavina, D.; Legzdina, L.; Jansons, J.; Piri, T. Development of Heterobasidion spp. fruit bodies on decayed Piecea abies. For. Ecol. Manag. 2021, 482, 118835. [Google Scholar] [CrossRef]
- García-Torreiro, M.; López-Abelairas, M.; Lu-Chau, T.A.; Lema, J.M. Fungal pretreatment of agricultural residues for bioethanol production. Ind. Crops Prod. 2016, 89, 486–492. [Google Scholar] [CrossRef]
- Chowdhary, P.; More, N.; Yadav, A.; Bharagava, R.N. Chapter 12—Ligninolytic Enzymes: An Introduction and Applications in the Food Industry. In Enzymes in Food Biotechnology; Kuddus, M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 181–195. [Google Scholar] [CrossRef]
- Mittar, D.; Khanna, P.K.; Marwaha, S.S.; Kennedy, J.F. Biobleaching of Pulp and Paper Mill Effluents by Phanerochaete chrysosporium. J. Chem. Technol. Biotechnol. 1992, 53, 81–92. [Google Scholar] [CrossRef]
- Fulekar, M.H.; Pathak, B.; Fulekar, J.; Godambe, T. Bioremediation of Organic Pollutants Using Phanerochaete chrysosporium. In Fungi as Bioremediators; Goltapeh, E.M., Danesh, Y.R., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 135–157. [Google Scholar] [CrossRef]
- De Mattos-Shipley, K.M.J.; Ford, K.L.; Alberti, F.; Banks, A.M.; Bailey, A.M.; Foster, G.D. The good, the bad and the tasty: The many roles of mushrooms. Stud. Mycol. 2016, 85, 125–157. [Google Scholar] [CrossRef]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Skočaj, M.; Gregori, A.; Grundner, M.; Sepčić, K.; Sežun, M. Hydrolytic and oxidative enzyme production through cultivation of Pleurotus ostreatus on pulp and paper industry wastes. Holzforschung 2018, 72, 813–817. [Google Scholar] [CrossRef]
- Habtemariam, S. Trametes versicolor (Synn. Coriolus versicolor) Polysaccharides in Cancer Therapy: Targets and Efficacy. Biomedicines 2020, 8, 135. [Google Scholar] [CrossRef]
- Doskocil, I.; Havlik, J.; Verlotta, R.; Tauchen, J.; Vesela, L.; Macakova, K.; Opletal, L.; Kokoska, L.; Rada, V. In vitro immunomodulatory activity, cytotoxicity and chemistry of some central European polypores. Pharm. Biol. 2016, 54, 2369–2376. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Kanawaku, R.; Masumoto, M.; Yanase, H. Efficient xylose fermentation by the brown rot fungus Neolentinus lepideus. Enzym. Microb. Technol. 2012, 50, 96–100. [Google Scholar] [CrossRef]
- Usman, M.; Murtaza, G.; Ditta, A. Nutritional, Medicinal, and Cosmetic Value of Bioactive Compounds in Button Mushroom (Agaricus bisporus): A Review. Appl. Sci. 2021, 11, 5943. [Google Scholar] [CrossRef]
- Manasseh, A.T.; Godwin, J.T.A.; Emanghe, E.U.; Borisde, O.O. Phytochemical properties of Ganoderma applanatum as potential agents in the application of nanotechnology in modern day medical practice. Asian Pac. J. Trop. Biomed. 2012, 2, S580–S583. [Google Scholar] [CrossRef]
- Mesa Ospina, N.; Ospina Alvarez, S.P.; Escobar Sierra, D.M.; Rojas Vahos, D.F.; Zapata Ocampo, P.A.; Ossa Orozco, C.P. Isolation of chitosan from Ganoderma lucidum mushroom for biomedical applications. J. Mater. Sci. Mater. Med. 2015, 26, 135. [Google Scholar] [CrossRef]
- ISO 18134-3:2015; Solid Biofuels—Determination of Moisture Content—Oven Dry Method—Part 3: Moisture in General Analysis Sample. ISO: Genva, Switzerland, 2015.
- CEN/TS 15104:2011; Solid Biofuels—Determination of Total Content of Carbon, Hydrogen and Nitrogen—Instrumental Methods. CEN: Brussels, Belgium, 2011.
- Maziero, R.; Cavazzoni, V.; Bononi, V.L.R. Screening of basidiomycetes for the production of exopolysaccharide and biomass in submerged culture. Rev. Microbiol. 1999, 30, 77–84. [Google Scholar] [CrossRef]
- Walker, G.M.; White, N.A. Introduction to Fungal Physiology. In Fungi; Wiley: Hoboken, NJ, USA, 2017; pp. 1–35. [Google Scholar] [CrossRef]
- Krupodorova, T.A.; Barshteyn, V.Y.; Sekan, A.S. Review of the basic cultivation conditions influence on the growth of basidiomycetes. Curr. Res. Environ. Appl. Mycol. (J. Fungal Biol.) 2021, 11, 494–531. [Google Scholar] [CrossRef]
- El-Enshasy, H.A. Chapter 9—Filamentous Fungal—Process Characteristics, Products, and Applications. In Bioprocessing for Value-Added Products from Renewable Resources; Yang, S.-T., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 225–261. [Google Scholar] [CrossRef]
- Khamrai, M.; Banerjee, S.L.; Kundu, P.P. A sustainable production method of mycelium biomass using an isolated fungal strain Phanerochaete chrysosporium (accession no: KY593186): Its exploitation in wound healing patch formation. Biocatal. Agric. Biotechnol. 2018, 16, 548–557. [Google Scholar] [CrossRef]
- Hoa, H.T.; Wang, C.L. The Effects of Temperature and Nutritional Conditions on Mycelium Growth of Two Oyster Mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus). Mycobiology 2015, 43, 14–23. [Google Scholar] [CrossRef]
- Akila, R.M. Fermentative production of fungal Chitosan, a versatile biopolymer (perspectives and its applications). Adv. Appl. Sci. Res. 2014, 5, 157–170. [Google Scholar]
- Crestini, C.; Kovac, B.; Giovannozzi-Sermanni, G. Production and isolation of chitosan by submerged and solid-state fermentation from Lentinus edodes. Biotechnol. Bioeng. 1996, 50, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, J.F.C.W.S. Structure, Synthesis, and Assembly, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar] [CrossRef]
- Huq, T.; Khan, A.; Brown, D.; Dhayagude, N.; He, Z.; Ni, Y. Sources, production and commercial applications of fungal chitosan: A review. J. Bioresour. Bioprod. 2022, 7, 85–98. [Google Scholar] [CrossRef]
- Ssekatawa, K.; Byarugaba, D.K.; Wampande, E.M.; Moja, T.N.; Nxumalo, E.; Maaza, M.; Sackey, J.; Ejobi, F.; Kirabira, J.B. Isolation and characterization of chitosan from Ugandan edible mushrooms, Nile perch scales and banana weevils for biomedical applications. Sci. Rep. 2021, 11, 4116. [Google Scholar] [CrossRef] [PubMed]
- De Elson Santiago, A. Characterization and Properties of Chitosan. In Biotechnology of Biopolymers; Magdy, E., Ed.; IntechOpen: Rijeka, Croatia, 2011; pp. 1–20. [Google Scholar] [CrossRef]
- Araújo, D.; Ferreira, I.C.; Torres, C.A.; Neves, L.; Freitas, F. Chitinous polymers: Extraction from fungal sources, characterization and processing towards value-added applications. J. Chem. Technol. Biotechnol. 2020, 95, 1277–1289. [Google Scholar] [CrossRef]
- Nwe, N.; Stevens, W.F.; Tokura, S.; Tamura, H. Characterization of chitosan and chitosan–glucan complex extracted from the cell wall of fungus Gongronella butleri USDB 0201 by enzymatic method. Enzym. Microb. Technol. 2008, 42, 242–251. [Google Scholar] [CrossRef]
- Martín-López, H.; Pech-Cohuo, S.C.; Herrera-Pool, E.; Medina-Torres, N.; Cuevas-Bernardino, J.C.; Ayora-Talavera, T.; Espinosa-Andrews, H.; Ramos-Díaz, A.; Trombotto, S.; Pacheco, N. Structural and Physicochemical Characterization of Chitosan Obtained by UAE and Its Effect on the Growth Inhibition of Pythium ultimum. Agriculture 2020, 10, 464. [Google Scholar] [CrossRef]
- Atykyan, N.; Revin, V.; Shutova, V. Raman and FT-IR Spectroscopy investigation the cellulose structural differences from bacteria Gluconacetobacter sucrofermentans during the different regimes of cultivation on a molasses media. AMB Express 2020, 10, 84. [Google Scholar] [CrossRef]
- Hong, T.; Yin, J.-Y.; Nie, S.-P.; Xie, M.-Y. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem. X 2021, 12, 100168. [Google Scholar] [CrossRef]
- Manal, G.; Mahmoud, E.E.K.; Mohsen, S.A. Chitin, Chitosan and Glucan, Properties and Applications. World J. Agric. Soil Sci. 2019, 3, 1–19. [Google Scholar] [CrossRef]
- Fesel, P.H.; Zuccaro, A. β-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants. Fungal Genet. Biol. 2016, 90, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Novak, M. Structural analysis of glucans. Ann. Transl. Med. 2014, 2, 17. [Google Scholar] [CrossRef] [PubMed]
- Szlek, D.B.; Reynolds, A.M.; Hubbe, M.A. Hydrophobic molecular treatments of cellulose-based or other polysaccharide barrier layers for sustainable food packaging: A Review. BioResources 2022, 17, 3551–3673. [Google Scholar] [CrossRef]
- Vikele, L.; Laka, M.; Sable, I.; Rozenberga, L.; Grinfelds, U.; Zoldners, J.; Passas, R.; Mauret, E. Effect of chitosan on properties of paper for packaging. Cellul. Chem. Technol. 2017, 51, 67–73. [Google Scholar]
- Andze, L.; Zoldners, J.; Rozenberga, L.; Sable, I.; Skute, M.; Laka, M.; Vecbiskena, L.; Andzs, M.; Actins, A. Effect of molecular chitosan on recovered paper properties described by mathematic model. Cellul. Chem. Technol. 2018, 52, 873–881. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungus | Dry Biomass (g 100 mL–1) | ||
|---|---|---|---|
| SSF | Shaken SF | Stationary SF | |
| P. ostreatus | 3.04 ± 0.39 | 0.63 ± 0.05 | 0.09 ± 0.03 |
| T. versicolor | 3.65 ± 0.66 | 0.48 ± 0.04 | 0.39 ± 0.03 |
| P. chrysosporium | 0.79 ± 0.21 | 1.03 ± 0.05 | 0.19 ± 0.00 |
| L. lepideus | 1.85 ± 0.29 | 0.40 ± 0.02 | 0.02 ± 0.01 |
| H. annosum | 0.75 ± 0.04 | 0.20 ± 0.03 | 0.36 ± 0.14 |
| Fungus | Cultivation | Moisture | AIS | Chitin-Containing Part AIS/Acid IS) | Glucans | 40% AIS | 40% AIS/Acid IS | Chitosan |
|---|---|---|---|---|---|---|---|---|
| H. annosum | SF | 6.8 | 11.6 ± 0.2 | 10.0 ± 0.6 | 0.12 ± 0.01 | 6.3 ± 0.2 | 2.7 ± 0.2 | 0.11 ± 0.04 |
| SSF | 6.1 | 12.4 ± 0.3 | 9.4 ± 0.3 | 0.23 ± 0.04 | 5.0 ± 0.3 | 2.5 ± 0.3 | 0.09 ± 0.02 | |
| P. chrysosporium | SF | 8.2 | 18.5 ± 0.7 | 13.7 ± 0.4 | 0.54 ± 0.03 | 4.4 ± 0.7 | 2.1 ± 0.3 | 0.38 ± 0.01 |
| SSF | 9.1 | 15.5 ± 0.5 | 12.7 ± 0.6 | 0.26 ± 0.02 | 3.4 ± 0.4 | 1.9 ± 0.4 | 0.23 ± 0.02 | |
| P. ostreatus | SF | 7.3 | 15.7 ± 0.4 | 13.3 ± 0.4 | 0.82 ± 0.03 | 6.7 ± 0.6 | 4.2 ± 0.2 | 0.09 ± 0.01 |
| SSF | 6.8 | 18.4 ± 0.3 | 13.8 ± 0.3 | 0.28 ± 0.01 | 2.2 ± 0.3 | 1.3 ± 0.1 | 0.021 ± 0.002 | |
| T. versicolor | SF | 8.4 | 15.4 ± 0.6 | 12.8 ± 0.3 | 0.17 ± 0.0.7 | 5.5 ± 0.2 | 3.3 ± 0.6 | 0.37 ± 0.02 |
| SSF | 7.3 | 30.9 ± 0.3 | 26.8 ± 0.5 | 0.10 ± 0.05 | 17.5 ± 0.8 | 15.2 ± 0.4 | 0.14 ± 0.02 | |
| L. lepideus | SF | 8.9 | 11.3 ± 0.8 | 10.1 ± 0.7 | 0.07 ± 0.03 | 5.2 ± 0.2 | 2.5 ± 0.2 | 0.039 ± 0.003 |
| SSF | 9.8 | 9.9 ± 0.2 | 8.9 ± 0.4 | 0.06 ± 0.02 | 4.8 ± 0.4 | 4.5 ± 0.4 | 0.035 ± 0.006 | |
| P. ostreatus | FB | 5.0 | 21.42 ± 0.4 | – | 0.04 ± 0.01 | – | – | 0.032 ± 0.002 |
| G.applanatum | FB | 8.3 | 48.05 ± 0.5 | – | 0.05 ± 0.04 | – | – | 0.054 ± 0.003 |
| A. bisporus | FB | 6.8 | 15.6 ± 0.2 | 14.3 ± 0.2 | 0.11 ± 0.01 | 8.4 ± 0.4 | 2.5 ± 0.2 | 1.72 ± 0.04 |
| Fungus | Nitrogen Content (%) | |||||
|---|---|---|---|---|---|---|
| Cultivation | Raw Material | AIS | Chitin-Containing Part (AIS/acid IS) | Glucans | Chitosan | |
| H. annosum | SF | 6.41 ± 0.07 | 3.48 ± 0.05 | 4.15 ± 0.06 | 0.54 ± 0.01 | 4.02 ± 0.02 |
| SSF | 3.38 ± 0.07 | 2.30 ± 0.06 | 2.99 ± 0.06 | 0.30 | - | |
| P. chrysosporium | SF | 5.30 ± 0.03 | 2.58 ± 0.20 | 2.88 ± 0.25 | 0.31 ± 0.07 | 6.00 ± 0.04 |
| SSF | 1.59 ± 0.01 | 1.52 ± 0.01 | 1.96 ± 0.08 | 0.53 | 5.43 | |
| P. ostreatus | SF | 5.78 ± 0.02 | 2.93 ± 0.02 | 3.09 ± 0.02 | 0.38 ± 0.02 | 2.77 ± 0.03 |
| SSF | 1.40 ± 0.28 | 1.09 ± 0.05 | 1.36 ± 0.01 | 0.27 | - | |
| T. versicolor | SF | 6.15 ± 0.25 | 2.36 ± 0.02 | 3.04 ± 0.20 | 0.14 ± 0.02 | 6.16 |
| SSF | 1.62 ± 0.03 | 1.01 ± 0.01 | 1.39 | - | 5.29 | |
| L. lepideus | SF | 7.29 ± 0.07 | 2.63 ± 0.13 | - | 0.17 ± 0.02 | - |
| SSF | 1.39 ± 0.01 | 1.01 ± 0.15 | - | - | - | |
| P. ostreatus | FB | 4.55 ± 0.05 | 1.35 ± 0.02 | 1.37 ± 0.02 | 0.14 ± 0.01 | 3.80 ± 0.01 |
| G. applanatum | FB | 3.07 ± 0.01 | 3.32 ± 0.03 | 3.01 ± 0.01 | - | 4.82 ± 0.04 |
| A. bisporus | FB | 3.95 ± 0.02 | 4.02 ± 0.02 | 4.25 ± 0.01 | 0.43 | 6.24 ± 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irbe, I.; Andze, L.; Blumfelde, M.; Filipova, I.; Verovkins, A.; Zoldners, J. Harvesting Mycelial Biomass of Selected Basidiomycetes for Chitosan Biopolymer Extraction. Polymers 2023, 15, 3548. https://doi.org/10.3390/polym15173548
Irbe I, Andze L, Blumfelde M, Filipova I, Verovkins A, Zoldners J. Harvesting Mycelial Biomass of Selected Basidiomycetes for Chitosan Biopolymer Extraction. Polymers. 2023; 15(17):3548. https://doi.org/10.3390/polym15173548
Chicago/Turabian StyleIrbe, Ilze, Laura Andze, Mara Blumfelde, Inese Filipova, Anrijs Verovkins, and Juris Zoldners. 2023. "Harvesting Mycelial Biomass of Selected Basidiomycetes for Chitosan Biopolymer Extraction" Polymers 15, no. 17: 3548. https://doi.org/10.3390/polym15173548