
DNA–Lysozyme Nanoarchitectonics: Quantitative Investigation on Charge Inversion and Compaction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
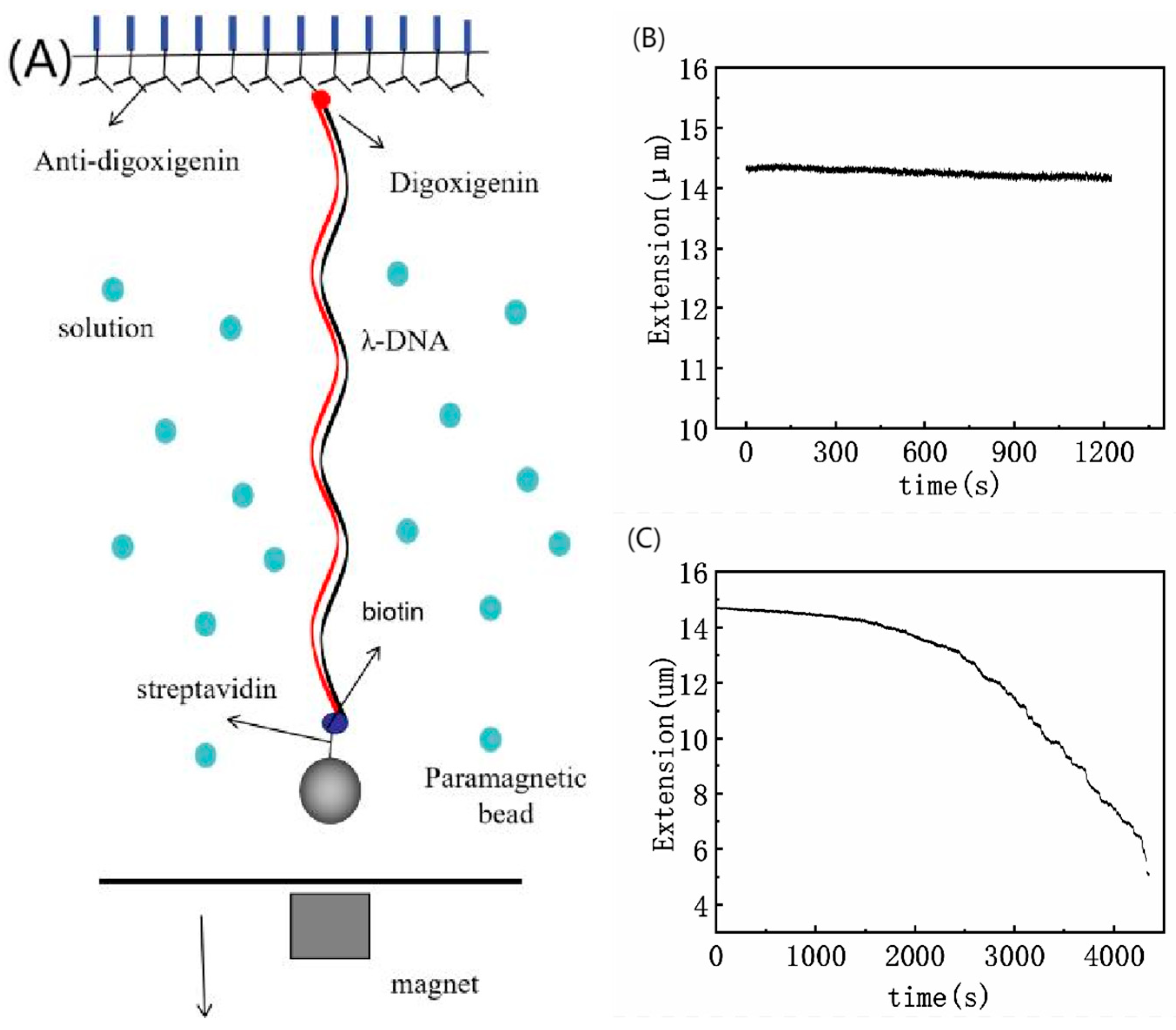
2.2. Magnetic Tweezers Experiment
2.3. Electrophoretic Mobility Measurement (EM) and Size Measurement by DLS
2.4. Atomic Force Microscopy (AFM)
3. Results
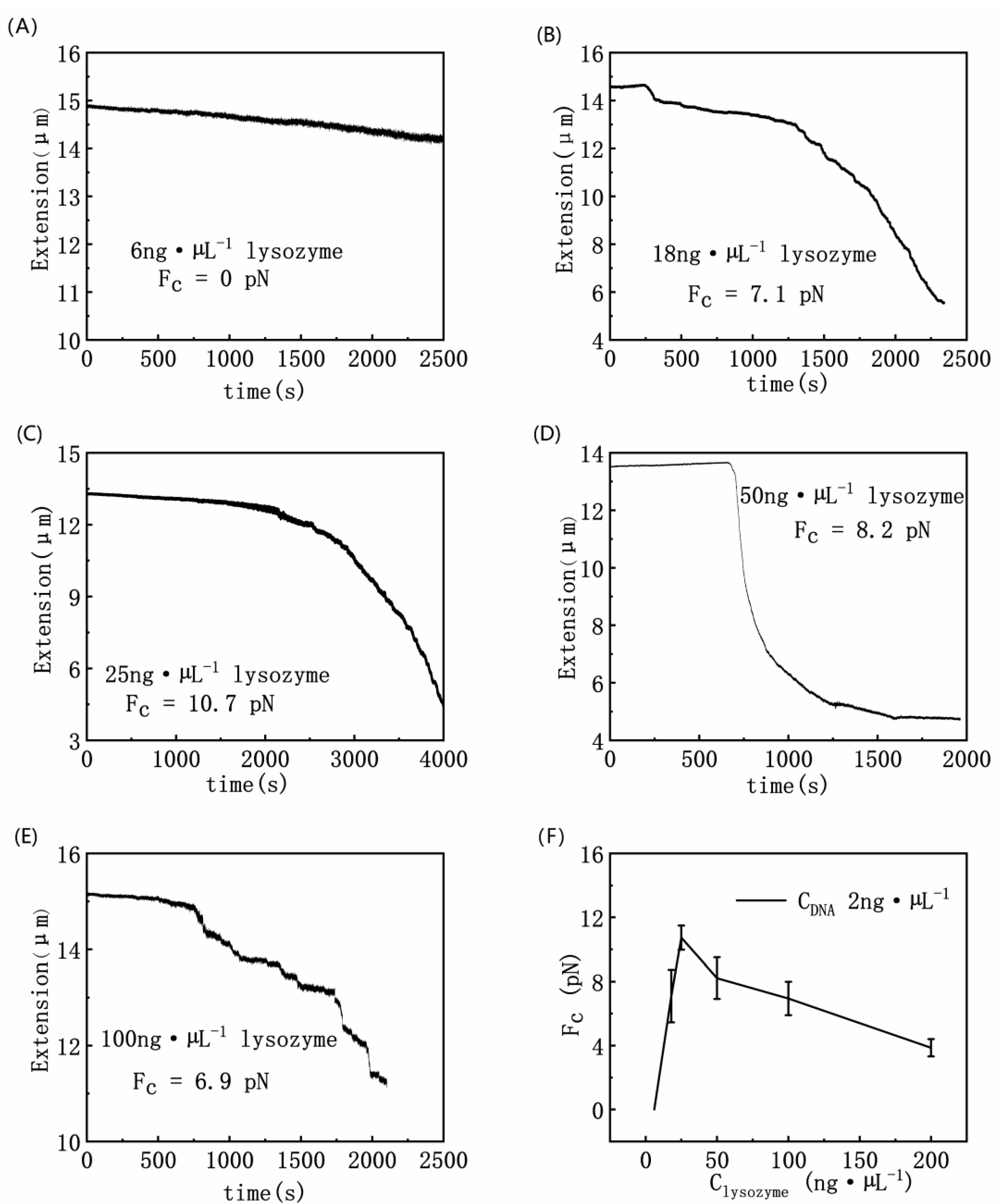
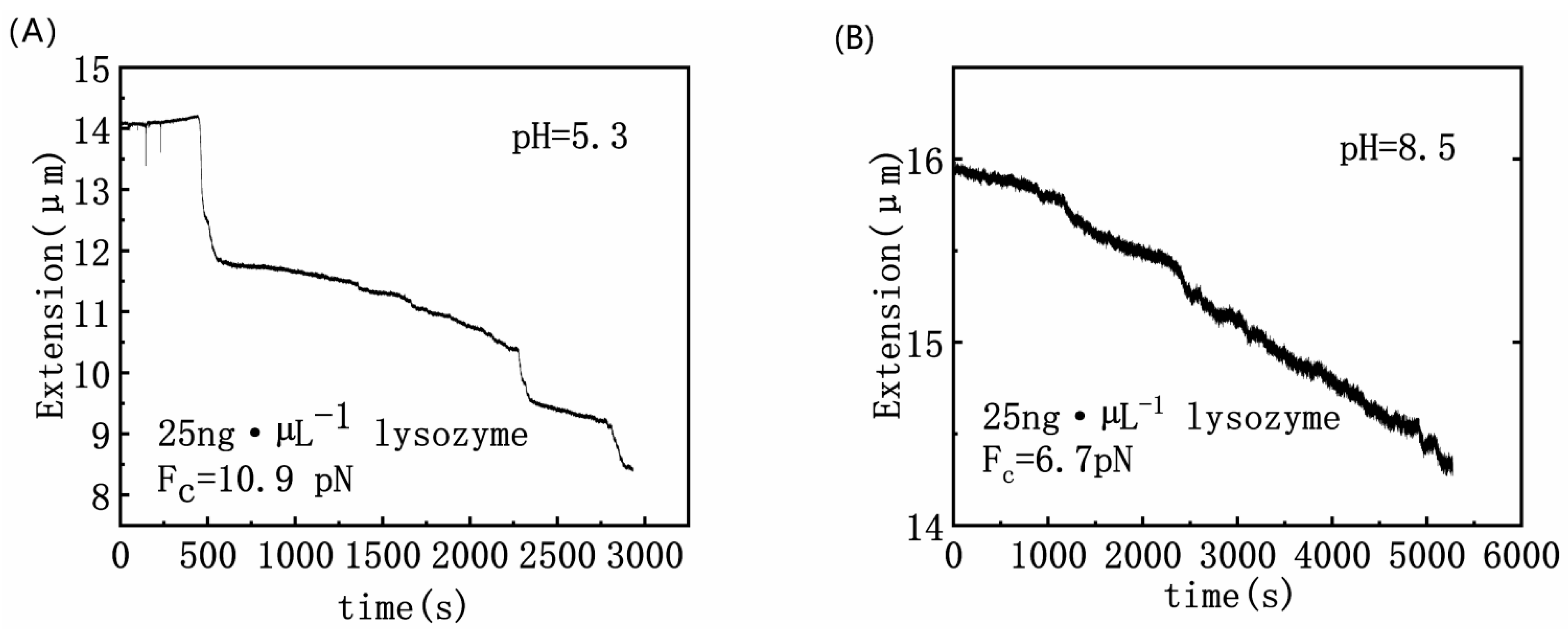
3.1. Condensing Force of DNA–Lysozyme Complex
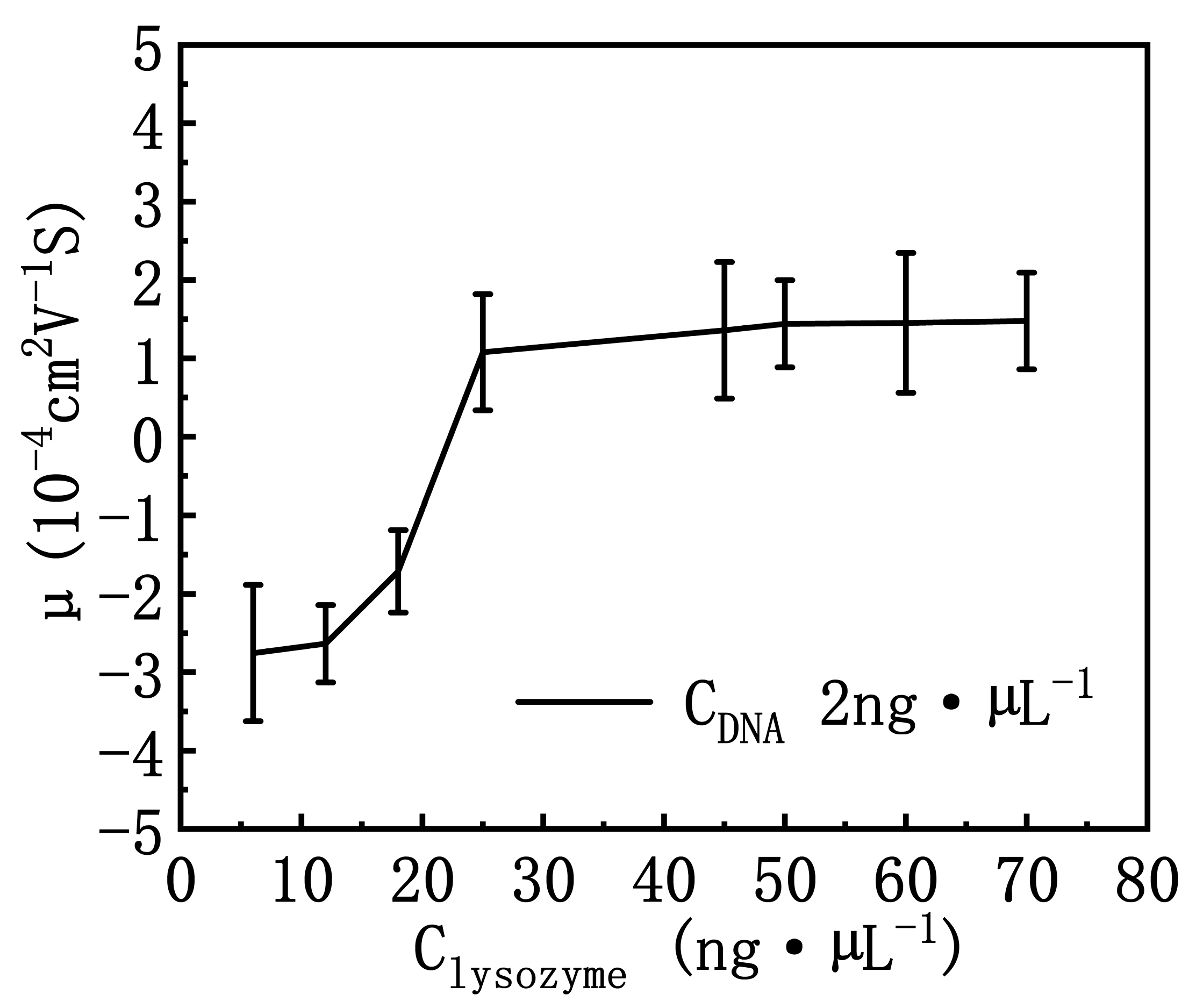
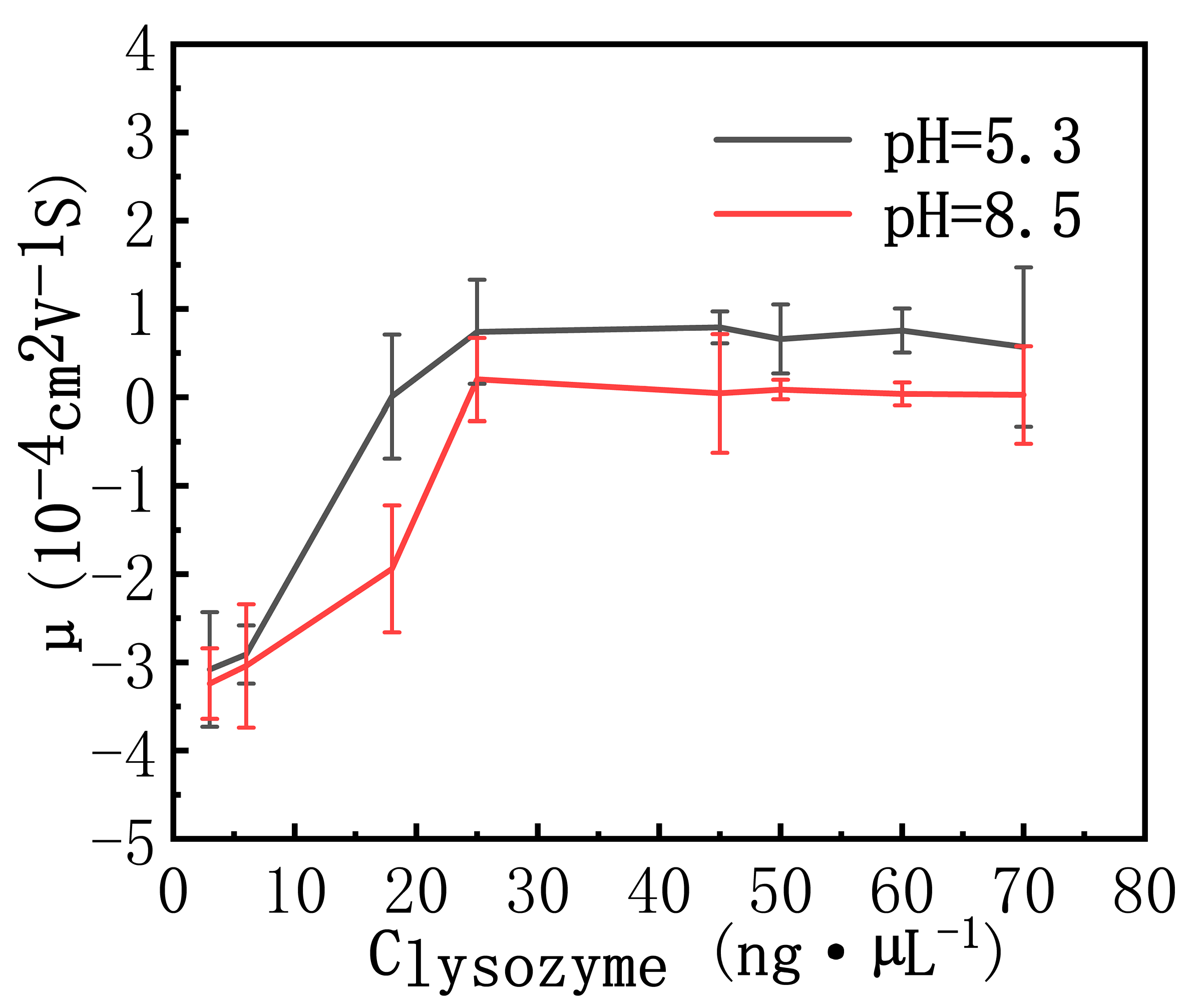
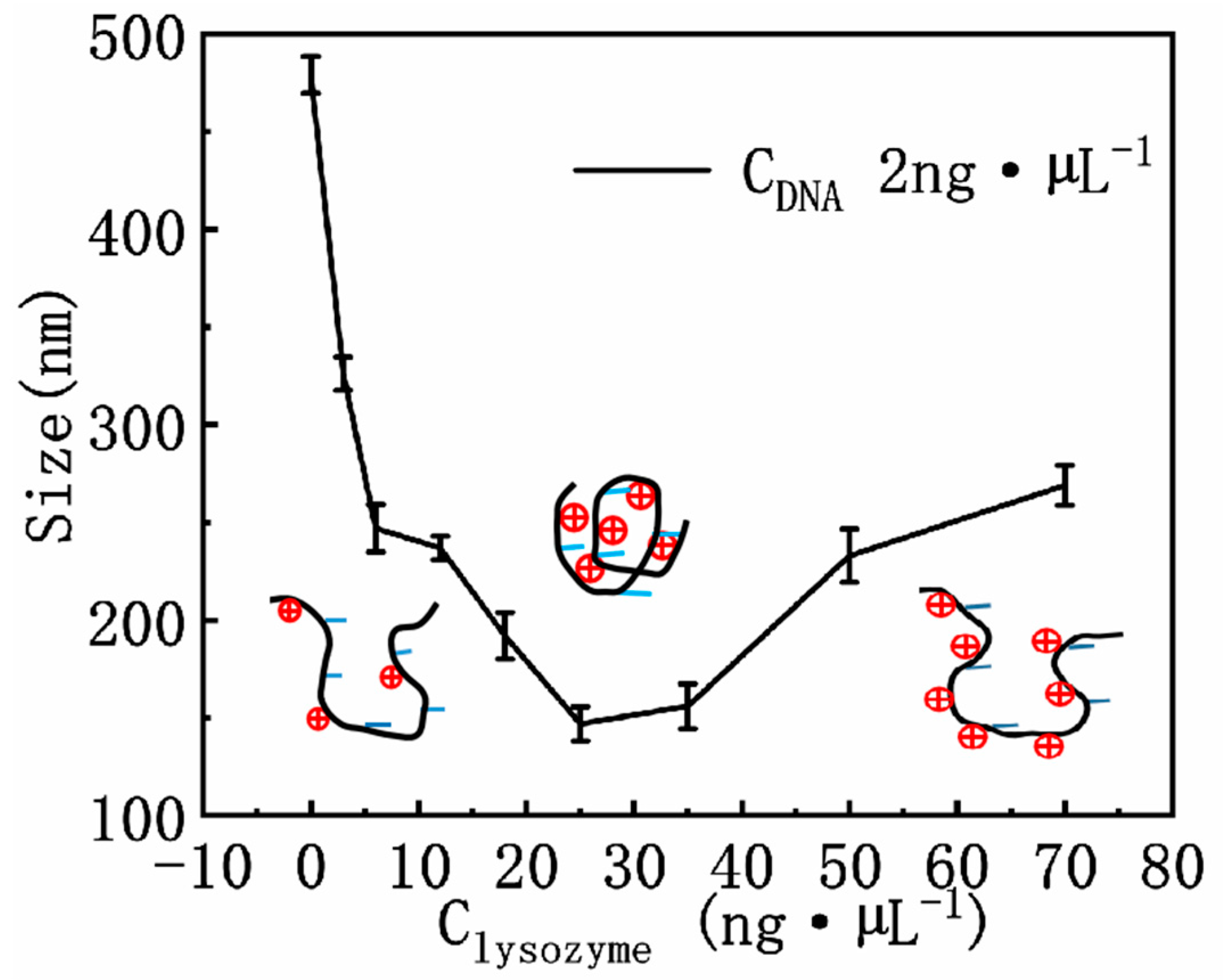
3.2. DNA Electrophoresis Mobility and Size in Lysozyme Solutions
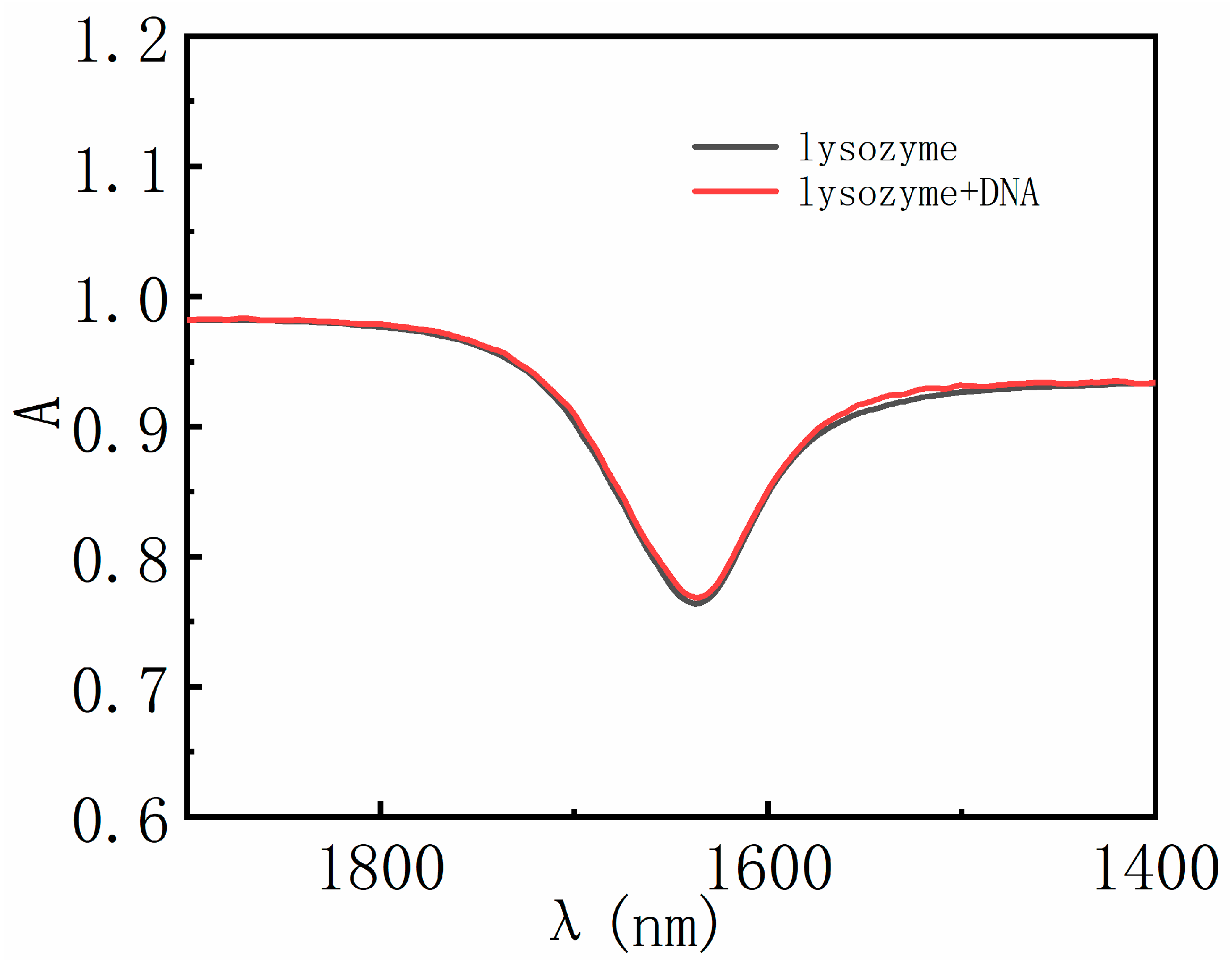
3.3. AFM Morphology of DNA
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alves, P.S.; Mesquita, O.N.; Rocha, M.S. Model for DNA interactions with proteins and other large ligands: Extracting physical chemistry from pure mechanical measurements. J. Phys. Chem. B 2020, 124, 1020–1024. [Google Scholar] [CrossRef]
- Wang, Y.; Ran, S.; Yang, G. Single molecular investigation of DNA looping and aggregation by restriction endonuclease BspMI. Sci. Rep. 2014, 4, 5897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, A.; Gope, A.; Kafle, R.; Iannacchione, G.S. Phase separation of a nematic liquid crystal in the self-assembly of lysozyme in a drying aqueous solution drop. MRS Commun. 2019, 9, 150–158. [Google Scholar] [CrossRef]
- Muschol, M.; Rosenberger, F. Liquid-liquid phase separation in supersaturated lysozyme solutions and associated precipitate formation/crystallization. J. Chem. Phys. 1997, 107, 1953–1962. [Google Scholar] [CrossRef]
- Taratuta, V.G.; Holschbach, A.; Thurston, G.M.; Blankschtein, D.; Benedek, G.B. Liquid-liquid phase separation of aqueous lysozyme solutions: Effects of pH and salt identity. J. Chem. Phys. 1990, 94, 2140–2144. [Google Scholar] [CrossRef]
- Grigsby, J.J.; Blanch, H.W.; Prausnitz, J.M. Cloud-point temperatures for lysozyme in electrolyte solutions: Effect of salt type, salt concentration and pH. Biophys. Chem. 2001, 91, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Blake, C.C.F.; Fenn, R.H.; North, A.C.T.; Phillips, D.C.; Poljak, R.J. Structure of lysozyme. a fourier map of the electron density at 6 angstrom resolution obtained by x-ray diffraction. Nature 1962, 196, 1173–1176. [Google Scholar] [CrossRef]
- Steinrauf, L.K.; Shiuan, D.; Yang, W.J.; Chiang, M.Y. Lysozyme association with nucleic acids. Biochem. Biophys. Res. Commun. 1999, 266, 366. [Google Scholar] [CrossRef]
- Lee-Huang, S.; Huang, P.L.; Sun, Y.; Huang, P.L.; Kung, H.F.; Blithe, D.L.; Chen, H.C. Lysozyme and RNases as anti-HIV components in β-core preparations of human chorionic gonadotropin. Proc. Natl. Acad. Sci. USA 1999, 96, 2678–2681. [Google Scholar] [CrossRef] [Green Version]
- Tao, Y.; Gang, W.; Zhuang, L. Electrostatic assembly of protein lysozyme on DNA visualized by atomic force microscopy. Appl. Surf. Sci. 2007, 253, 4311–4316. [Google Scholar]
- Lin, K.C.; Wey, M.T.; Kan, L.S.; Shiuan, D. Characterization of the Interactions of Lysozyme with DNA by Surface Plasmon Resonance and Circular Dichroism Spectroscopy. Appl. Biochem. Biotechnol. 2009, 158, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Dan, L.; Carnerup, A.M.; Janiak, J.; Schillén, K.; Miguel, M.D.G.; Lindman, B. Size and morphology of assemblies formed by DNA and lysozyme in dilute aqueous mixtures. Phys. Chem. Chem. Phys. 2011, 13, 3082–3091. [Google Scholar]
- Lundberg, D.; Carnerup, A.M.; Schillén, K.; Miguel, M.; Lindman, B. Phase behavior and coassembly of DNA and lysozyme in dilute aqueous mixtures: A model investigation of DNA-protein interactions. Langmuir 2010, 26, 2986–2988. [Google Scholar] [CrossRef]
- Ariga, K. Nanoarchitectonics: What’s Coming Next to Nanotechnology. Nanoscale Horiz. 2021, 6, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.; Jnsson, B. On the charge regulation of proteins. Biochemistry 2005, 44, 5722–5727. [Google Scholar] [CrossRef]
- Wang, Y.; Ran, S.; Man, B.; Yang, G. DNA condensations on mica surfaces induced collaboratively by alcohol and hexammine cobalt. Colloids Surf. B 2011, 83, 61–68. [Google Scholar] [CrossRef]
- Qiu, S.; Wang, Y.; Cao, B.; Guo, Z.; Chen, Y.; Yang, G. The suppression and promotion of DNA charge inversion by mixing counterions. Soft Matter 2015, 11, 4099–4105. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Cao, B.; Guo, Z.; Yang, G. Single molecular demonstration of modulating charge inversion of DNA. Sci. Rep. 2016, 6, 38628. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Tianyong, G.; Yanwei, W.; Guangcan, Y. Investigating the influence of magnesium Ions on p53–DNA binding using atomic force microscopy. Int. J. Mol. Sci. 2017, 18, 1585. [Google Scholar]
- Wang, Y.; Wang, R.; Gao, T.; Yang, G. The Mixing counterion effect on DNA compaction and charge neutralization at low ionic strength. Polymers 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Wang, Y.; Yang, G. The promotion and suppression of DNA charge neutralization by the cosolute ectoine. RSC Adv. 2019, 9, 41050–41057. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Zhang, W.; Wang, Y.; Yang, G. DNA compaction and charge neutralization regulated by divalent ions in very low pH solution. Polymers 2019, 11, 337. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, T.; Li, S.; Xia, W.; Zhang, W.; Yang, G. Direct demonstration of DNA compaction mediated by divalent counterions. J. Phys. Chem. B 2019, 123, 79–85. [Google Scholar] [CrossRef]
- Li, N.; Liao, Z.; He, S.; Chen, X.; Huang, S.; Wang, Y.; Yang, G. Demonstration of pH-controlled DNA–surfactant manipulation for biomolecules. RSC Adv. 2021, 11, 15099–15105. [Google Scholar] [CrossRef]
- Wang, Y.; Ran, S.; Man, B.; Yang, G. Ethanol induces condensation of single DNA molecules. Soft Matter 2011, 7, 4425. [Google Scholar] [CrossRef]
- Yoshizaki, I.; Nakamura, H.; Sato, T. Systematic analysis of the effect of supersaturation on protein crystal quality. J. Cryst. Growth 2002, 237, 295–299. [Google Scholar] [CrossRef]
- Dold, P.; Ono, E.; Tsukamoto, K.; Sazaki, G. Step velocity in tetragonal lysozyme growth as a function of impurity concentration and mass transport conditions. J. Cryst. Growth 2006, 293, 102–109. [Google Scholar] [CrossRef]
- Gliko, O.; Booth, N.A.; Rosenbach, E.; Vekilov, P.G. Phase-shifting interferometry for the study of the step dynamics during crystallization of proteins. Cryst. Growth Des. 2002, 2, 381–385. [Google Scholar] [CrossRef]
- Velev, O.D.; Kaler, E.W.; Lenhoff, A.M. Protein interactions in solution characterized by light and neutron scattering: Comparison of lysozyme and chymotrypsinogen. Biophys. J. 1998, 75, 2682–2697. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.B.; da Costa, A.R.; Garcia-Rojas, E.E. Heteroprotein complex coacervates of ovalbumin and lysozyme: Formation and thermodynamic characterization. Int. J. Biol. Macromol. 2018, 106, 1323–1329. [Google Scholar] [CrossRef]
- Xi, B.; Ran, S.Y. Formation of DNA pearl-necklace structures on mica surface governed by kinetics and thermodynamics. J. Polym. Sci. Pol. Phys. 2017, 55, 971–979. [Google Scholar] [CrossRef]
- Yan, J.; Skoko, D.; Marko, J.F. Near-field-magnetic-tweezer manipulation of single DNA molecules. Phys. Rev. E 2004, 70, 011905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Wang, Y.; Yang, A.; Yang, G. The effect of pH on charge inversion and condensation of DNA. Soft Matter 2016, 12, 6669–6674. [Google Scholar] [CrossRef]
- Smith, S.; Finzi, L.; Bustamante, C. Direct mechanical measurements of the elasticity of single DNA molecules by using magnetic beads. Science 1992, 258, 1122–1126. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Yang, G. Single molecular analysis of the interaction between DNA and chitosan. RSC Adv. 2015, 5, 29594–29600. [Google Scholar] [CrossRef]
- Bloomfield, V.A. DNA condensation by multivalent cations. Biopolymers 1997, 44, 269–282. [Google Scholar] [CrossRef]
- Tanaka, M.; Grosberg, A.Y. Electrophoresis of a charge-inverted macroion complex: Molecular-dynamics study. Eur. Phys. J. E 2002, 7, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.; Li, H.; Yang, W.; Luo, Y. Accurate Determination of Interfacial Protein Secondary Structure by Combining Interfacial-Sensitive Amide I and Amide III Spectral Signals. J. Am. Chem. Soc. 2014, 136, 1206–1209. [Google Scholar] [CrossRef]
- Besteman, K.; Van Eijk, K.; Lemay, S.G. Charge inversion accompanies DNA condensation by multivalent ions. Nat. Phys. 2007, 3, 641–644. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Wang, Y.; Yang, G. DNA–Lysozyme Nanoarchitectonics: Quantitative Investigation on Charge Inversion and Compaction. Polymers 2022, 14, 1377. https://doi.org/10.3390/polym14071377
Zhang R, Wang Y, Yang G. DNA–Lysozyme Nanoarchitectonics: Quantitative Investigation on Charge Inversion and Compaction. Polymers. 2022; 14(7):1377. https://doi.org/10.3390/polym14071377
Chicago/Turabian StyleZhang, Rongyan, Yanwei Wang, and Guangcan Yang. 2022. "DNA–Lysozyme Nanoarchitectonics: Quantitative Investigation on Charge Inversion and Compaction" Polymers 14, no. 7: 1377. https://doi.org/10.3390/polym14071377