

Structural Characterization and Glycosaminoglycan Impurities Analysis of Chondroitin Sulfate from Chinese Sturgeon
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Chinese Sturgeon Cartilage
2.3. Extraction of CS from Chinese Sturgeon Cartilage
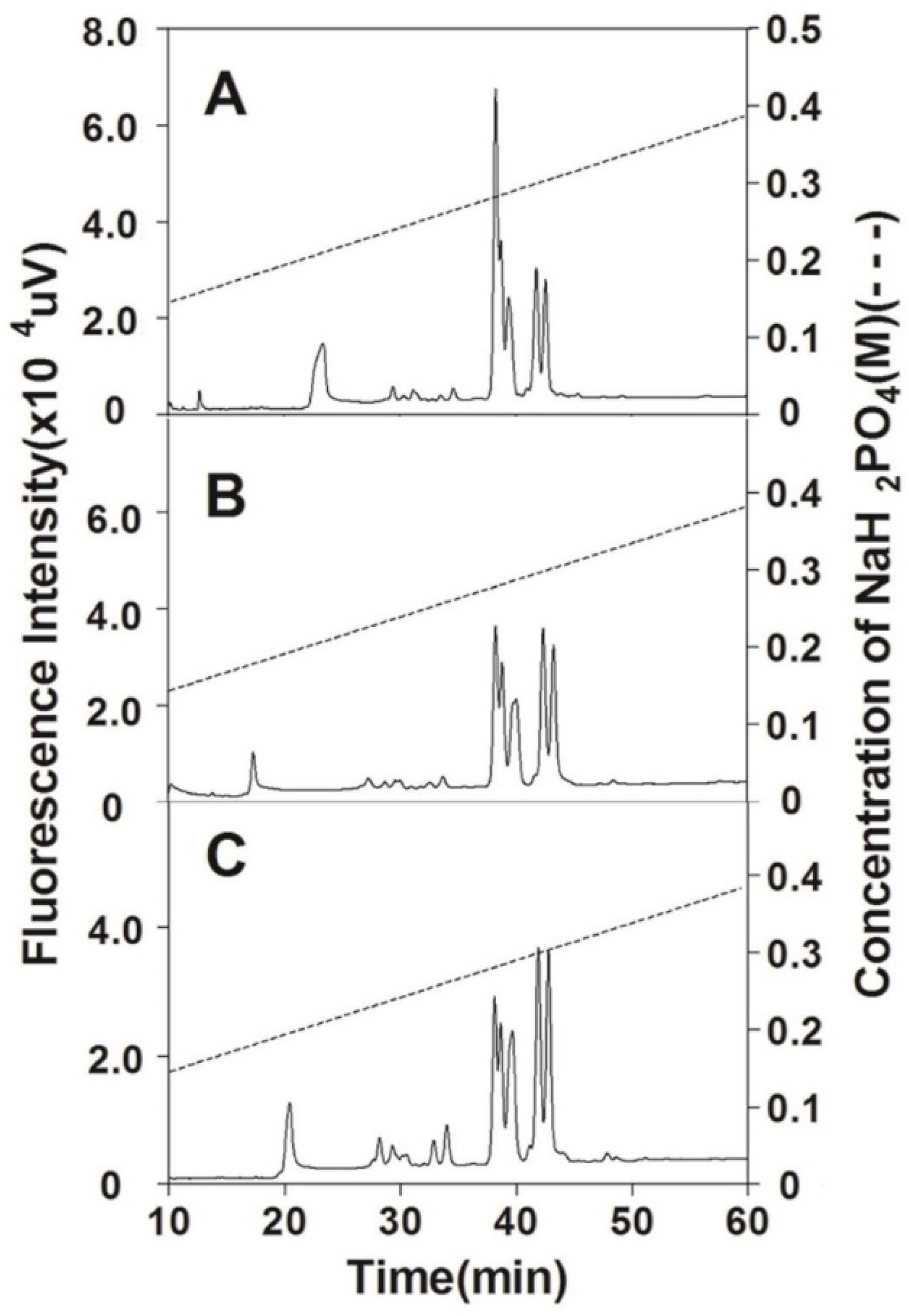
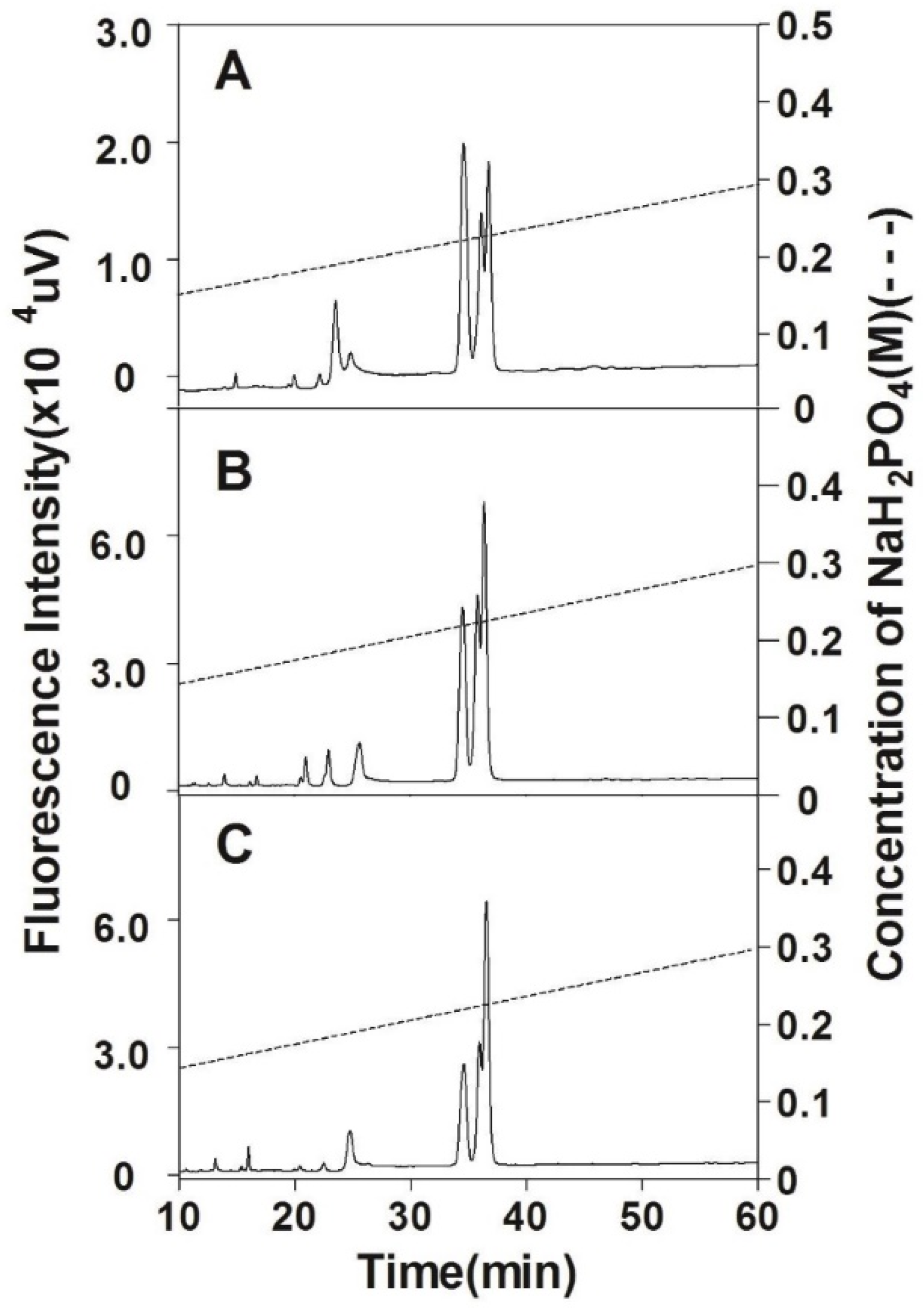
2.4. Purification of CS Preparations
2.5. Enzyme Analysis of CS Preparations
2.6. Analysis of Molecular Mass
2.7. Analysis of Disaccharide Composition
2.8. NMR Analysis of Purified CS Samples
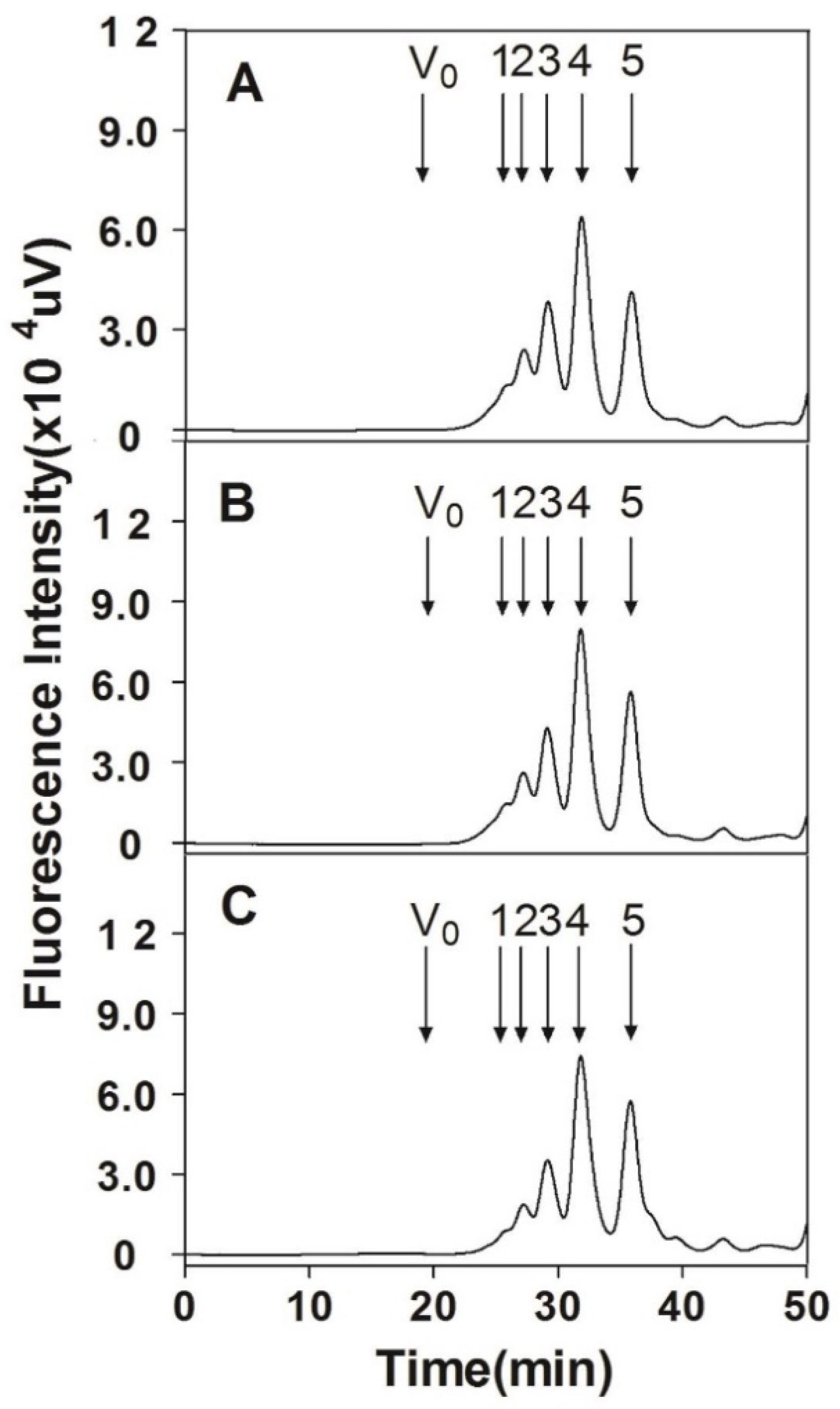
2.9. Analysis of Size-Defined Oligosaccharides
2.10. Analysis of Anticoagulant Activity
3. Results
3.1. Preparation of CS from Chinese Sturgeon Cartilage
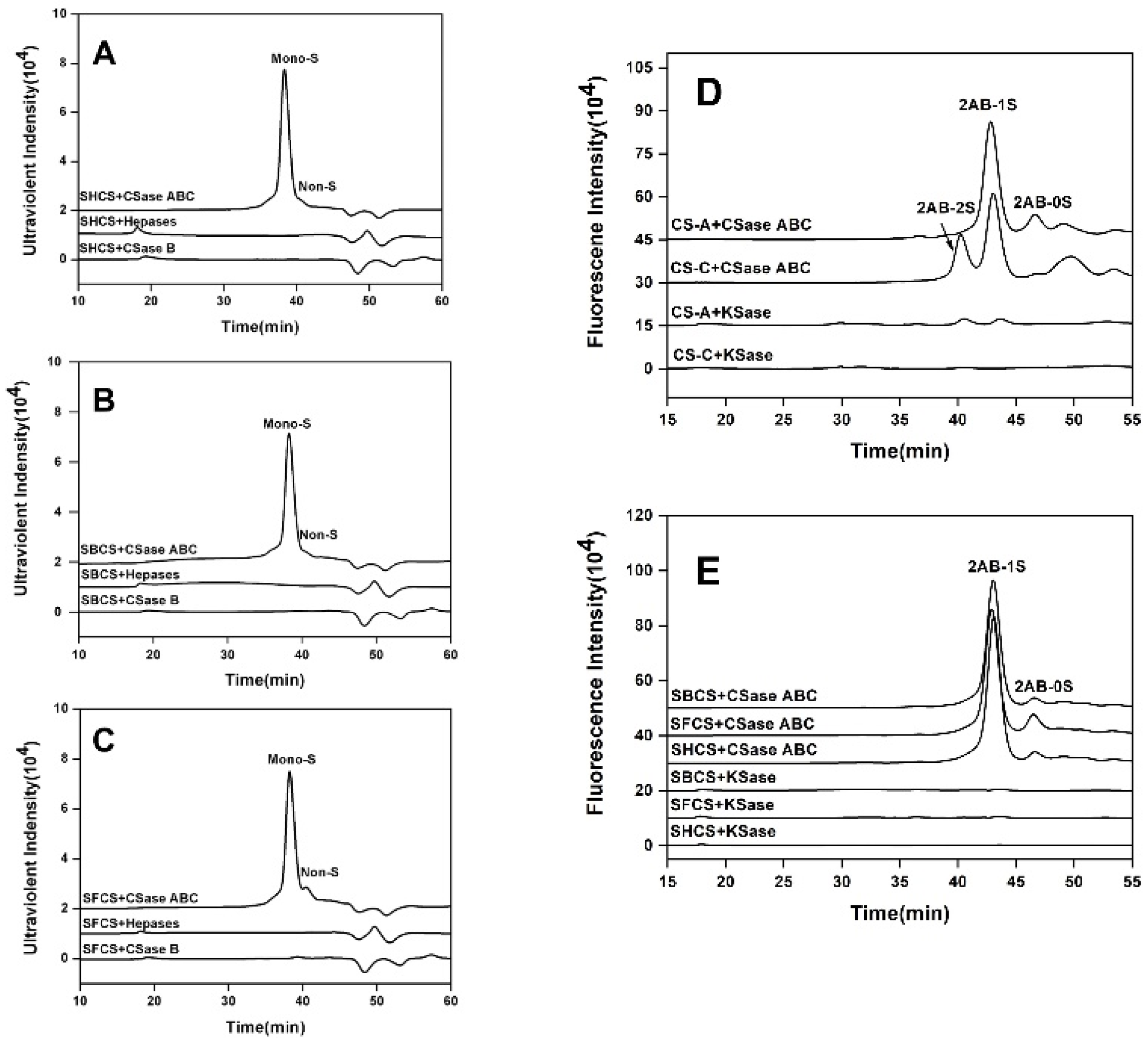
3.2. Enzyme Analysis of CS Preparations
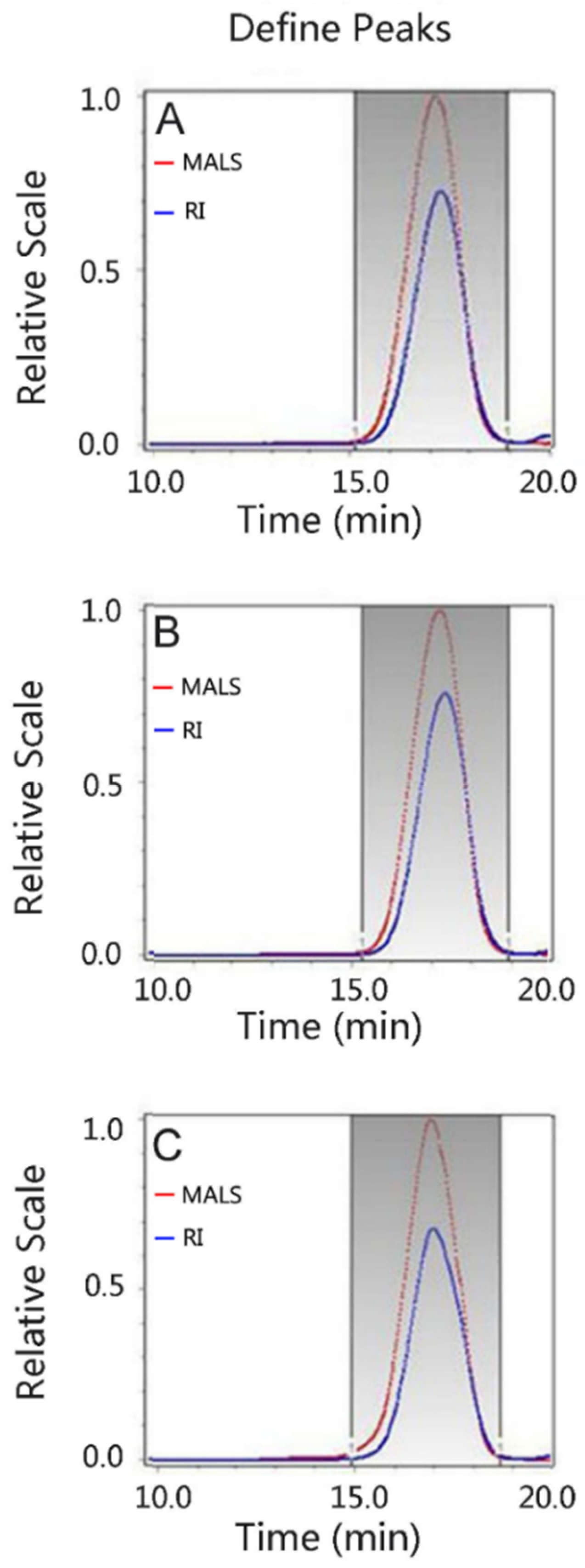
3.3. Analysis of the Molecular Mass
3.4. Analysis of the Disaccharide Composition
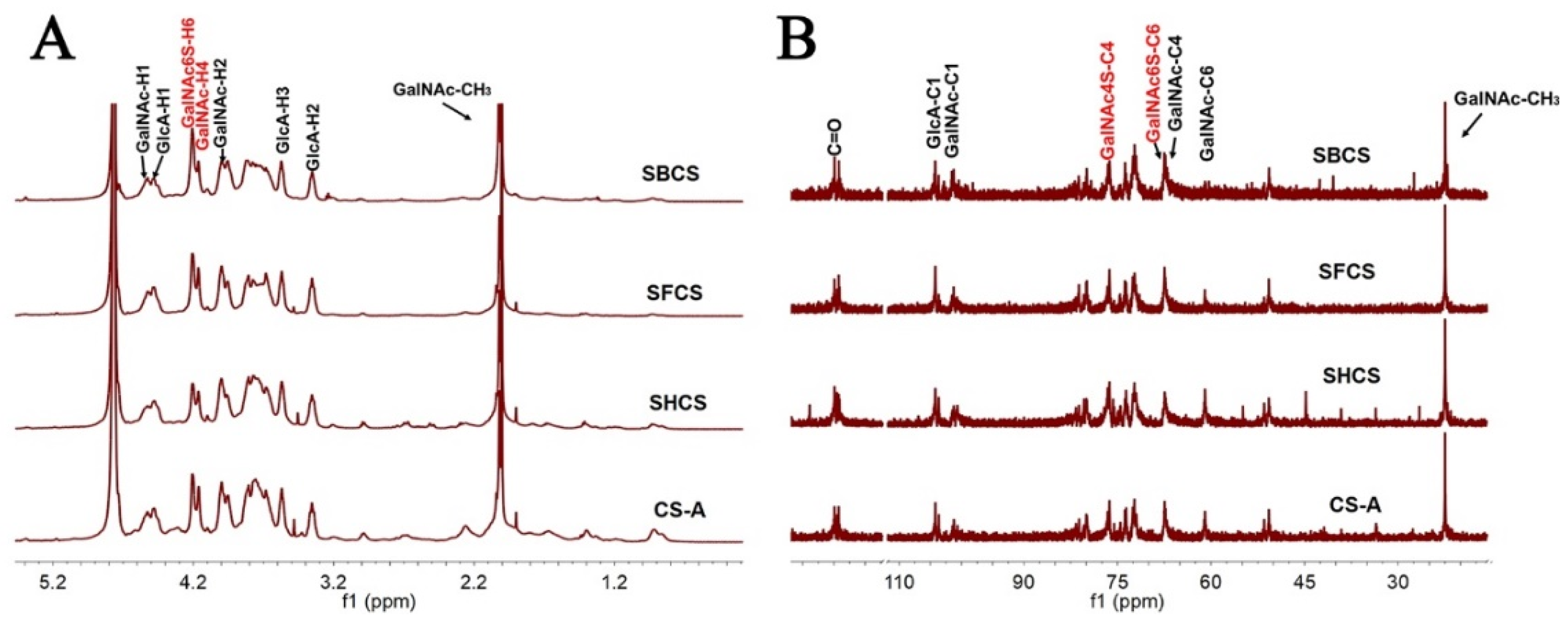
3.5. NMR Spectroscopy
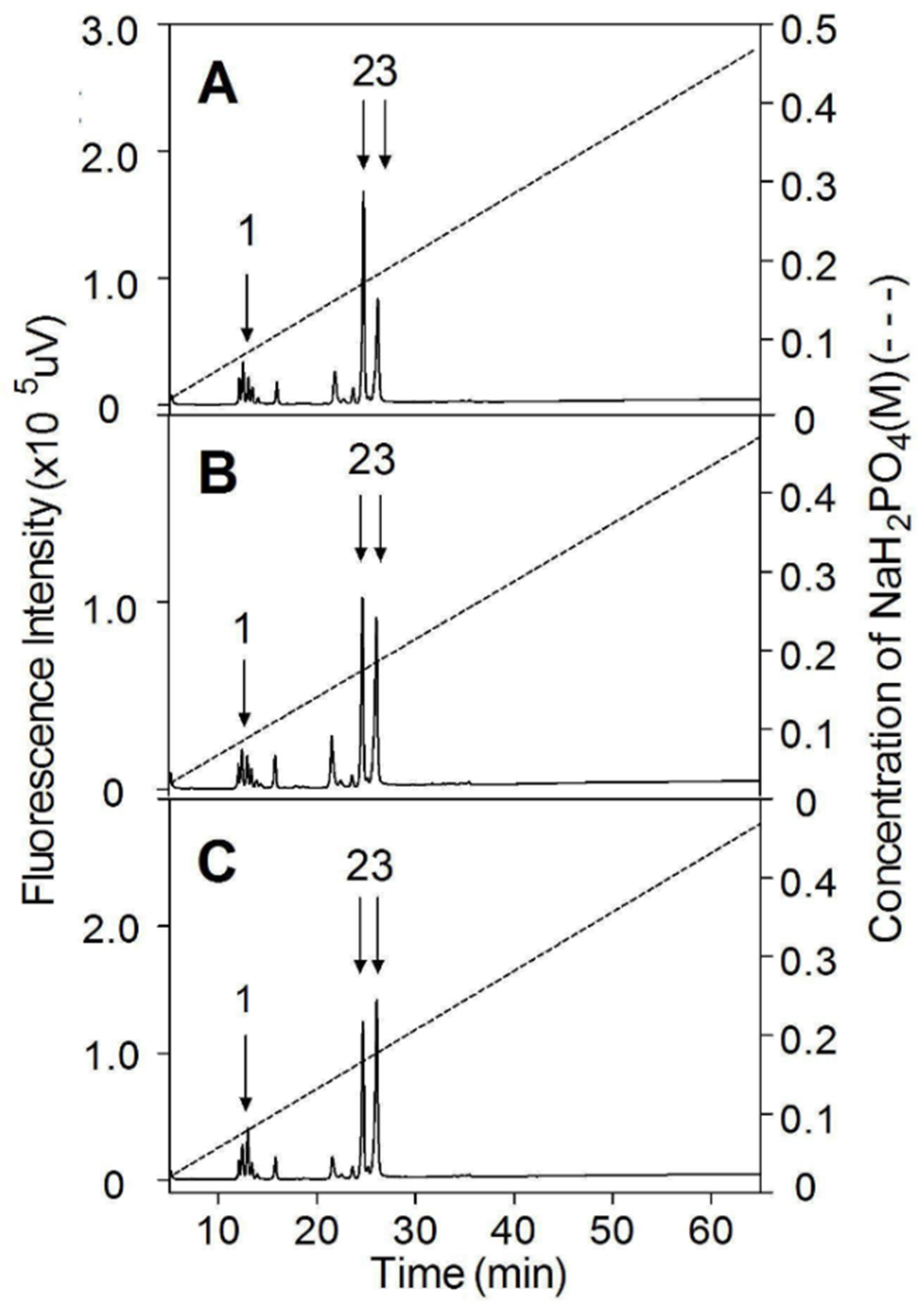
3.6. Analysis of Internal Structure
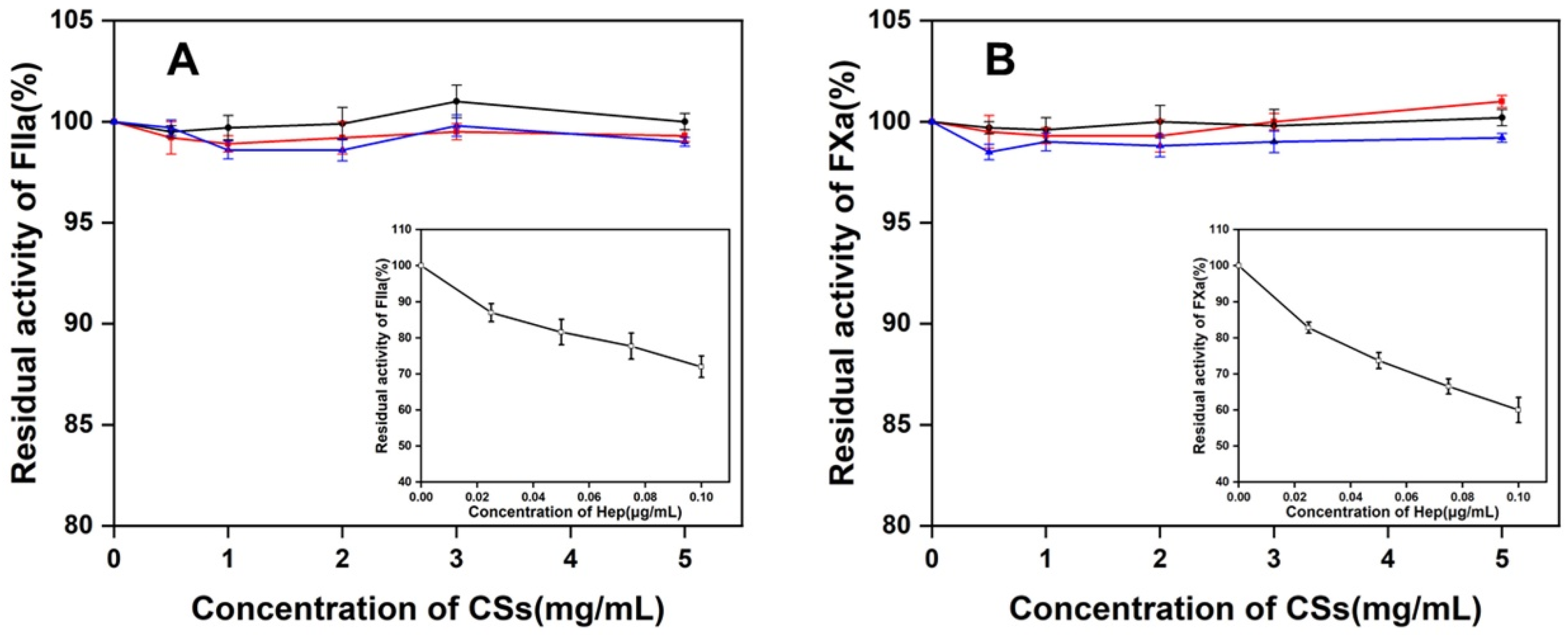
3.7. Effect of Three CS Preparations to the FXa or FIIa in the Presence of ATIII
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sugahara, K.; Mikami, T.; Uyama, T.; Mizuguchi, S.; Nomura, K.; Kitagawa, H. Recent Advances in the Structural Biology of Chondroitin Sulfate and Dermatan Sulfate. Curr. Opin. Struct. Biol. 2003, 13, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Silbert, J.E.; Sugumaran, G. Biosynthesis of Chondroitin/Dermatan Sulfate. IUBMB Life 2002, 54, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Nadanaka, S.; Clement, A.; Masayama, K.; Faissner, A.; Sugahara, K. Characteristic Hexasaccharide Sequences in Octasaccharides Derived from Shark Cartilage Chondroitin Sulfate D with a Neurite Outgrowth Promoting Activity. J. Biol. Chem. 1998, 273, 3296–3307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Masuko, S.; Solakyildirim, K.; Pu, D.; Linhardt, R.J.; Zhang, F. Glycosaminoglycans of the Porcine Central Nervous System. Biochemistry 2010, 49, 9839–9847. [Google Scholar] [CrossRef] [Green Version]
- Lafont, F.; Rouget, M.; Triller, A.; Prochiantz, A.; Rousselet, A. In Vitro Control of Neuronal Polarity by Glycosaminoglycans. Development 1992, 114, 17–29. [Google Scholar] [CrossRef]
- Faissner, A.; Clement, A.; Lochter, A.; Streit, A.; Mandl, C.; Schachner, M. Isolation of a Neural Chondroitin Sulfate Proteoglycan with Neurite Outgrowth Promoting Properties. J. Cell Biol. 1994, 126, 783–799. [Google Scholar] [CrossRef]
- Mencio, C.P.; Hussein, R.K.; Yu, P.; Geller, H.M. The Role of Chondroitin Sulfate Proteoglycans in Nervous System Development. J. Histochem. Cytochem. 2021, 69, 61–80. [Google Scholar] [CrossRef]
- Clement, A.M.; Nadanaka, S.; Masayama, K.; Mandl, C.; Sugahara, K.; Faissner, A. The DSD-1 Carbohydrate Epitope Depends on Sulfation, Correlates with Chondroitin Sulfate D Motifs, and Is Sufficient to Promote Neurite Outgrowth. J. Biol. Chem. 1998, 273, 28444–28453. [Google Scholar] [CrossRef] [Green Version]
- Trowbridge, J.M.; Rudisill, J.A.; Ron, D.; Gallo, R.L. Dermatan Sulfate Binds and Potentiates Activity of Keratinocyte Growth Factor (FGF-7). J. Biol. Chem. 2002, 277, 42815–42820. [Google Scholar] [CrossRef] [Green Version]
- Penc, S.F.; Pomahac, B.; Winkler, T.; Dorschner, R.A.; Eriksson, E.; Herndon, M.; Gallo, R.L. Dermatan Sulfate Released after Injury Is a Potent Promoter of Fibroblast Growth Factor-2 Function. J. Biol. Chem. 1998, 273, 28116–28121. [Google Scholar] [CrossRef]
- Williams, R.K.; Straus, S.E. Specificity and Affinity of Binding of Herpes Simplex Virus Type 2 Glycoprotein B to Glycosaminoglycans. J. Virol. 1997, 71, 1375–1380. [Google Scholar] [CrossRef] [Green Version]
- Bergefall, K.; Trybala, E.; Johansson, M.; Uyama, T.; Naito, S.; Yamada, S.; Kitagawa, H.; Sugahara, K.; Bergström, T. Chondroitin Sulfate Characterized by the E-Disaccharide Unit Is a Potent Inhibitor of Herpes Simplex Virus Infectivity and Provides the Virus Binding Sites on Gro2C Cells. J. Biol. Chem. 2005, 280, 32193–32199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantazaka, E.; Papadimitriou, E. Chondroitin Sulfate-Cell Membrane Effectors as Regulators of Growth Factor-Mediated Vascular and Cancer Cell Migration. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 2643–2650. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.R.; Gallo, R.L. Glycosaminoglycans and Their Proteoglycans: Host-Associated Molecular Patterns for Initiation and Modulation of Inflammation. FASEB J. 2006, 20, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.Y.; Olson, S.K.; Esko, J.D.; Horvitz, H.R. Caenorhabditis elegans Early Embryogenesis and Vulval Morphogenesis Require Chondroitin Biosynthesis. Nature 2003, 423, 439–443. [Google Scholar] [CrossRef]
- Mizuguchi, S.; Uyama, T.; Kitagawa, H.; Nomura, K.H.; Dejima, K.; Gengyo-Ando, K.; Mitani, S.; Sugahara, K.; Nomura, K. Chondroitin Proteoglycans Are Involved in Cell Division of Caenorhabditis elegans. Nature 2003, 423, 443–448. [Google Scholar] [CrossRef]
- White, J.; Bednarek, S. Cytokinesis: GAGs form the Walls that Separate Our Parts. Curr. Biol. 2003, 13, 717–718. [Google Scholar] [CrossRef] [Green Version]
- Olson, S.K.; Bishop, J.R.; Yates, J.R.; Oegema, K.; Esko, J.D. Identification of Novel Chondroitin Proteoglycans in Caenorhabditis elegans: Embryonic Cell Division Depends on CPG-1 and CPG-2. J. Cell Biol. 2006, 173, 985–994. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, T.; Zhuang, P.; Zhang, L.Z.; Liu, T. Annotation and Analysis of Expressed Sequence Tags (ESTs) from Chinese Sturgeon (Acipenser sinensis) Pituitary cDNA Library. Mar. Genom. 2011, 4, 173–179. [Google Scholar] [CrossRef]
- Zhuang, P.; Kynard, B.; Zhang, L.; Zhang, T.; Cao, W. Ontogenetic Behavior and Migration of Chinese Sturgeon, Acipenser sinensis. Environ. Biol. Fishes 2002, 65, 83–97. [Google Scholar] [CrossRef]
- Ludwig, A.; Belfiore, N.M.; Pitra, C.; Svirsky, V.; Jenneckens, I. Genome Duplication Events and Functional Reduction of Ploidy Levels in Sturgeon (Acipenser, Huso and Scaphirhynchus). Genetics 2001, 158, 1203–1215. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Gan, F.; Chen, X.H.; Liu, Z.G.; Li, L.X.; Wei, Q.W.; Tang, Y.K. Molecular and Expression Analysis of Apolipoprotein E Gene in the Chinese Sturgeon, Acipenser sinensis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 158, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Im, A.R.; Park, Y.; Kim, Y.S. Isolation and Characterization of Chondroitin Sulfates from Sturgeon (Acipenser sinensis) and Their Effects on Growth of Fibroblasts. Biol. Pharm. Bull. 2010, 33, 1268–1273. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Zhou, Y.; Mao, G.; Zou, Y.; Zhao, J.; Bai, S.; Yang, L.; Wu, X. Extraction, Purification and Characterisation of Chondroitin Sulfate in Chinese Sturgeon Cartilage. J. Sci. Food Agric. 2013, 93, 1633–1640. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, S.; Ren, S.; Zhang, X.; Yang, F.; Chen, Y.; Wang, B. Structural Characterization and Proliferation Activity of Chondroitin Sulfate from the Sturgeon, Acipenser schrenckii. Int. J. Biol. Macromol. 2020, 164, 3005–3011. [Google Scholar] [CrossRef]
- Wu, R.; Shang, N.; Gui, M.; Yin, J.; Li, P. Sturgeon (Acipenser)-Derived Chondroitin Sulfate Suppresses Human Colon Cancer HCT-116 both in vitro and in vivo by Inhibiting Proliferation and Inducing Apoptosis. Nutrients 2020, 12, 1130. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Liu, K.; Zha, F.; Wang, H.; Gao, R.; Wang, J.; Li, K.; Xu, X.; Zhao, Y. Preparation and Characterization of Chondroitin Sulfate from Large Hybrid Sturgeon Cartilage by Hot-Pressure and Its Effects on Acceleration of Wound Healing. Int. J. Biol. Macromol. 2022, 209, 1685–1694. [Google Scholar] [CrossRef]
- Wang, K.; Bai, F.; Zhou, X.; Wang, J.; Li, Y.; Xu, H.; Gao, R.; Wu, H.; Liu, K.; Zhao, Y. Characterization of Chondroitin Sulfates Isolated from Large Hybrid Sturgeon Cartilage and Their Gastroprotective Activity against Ethanol-Induced Gastric Ulcers. Food Chem. 2021, 363, 130436. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, K.; Suzuki, K.; Imai, K.; Mochizuki, H.; Morikawa, K.; Kyogashima, M.; Kimata, K.; Watanabe, H. Purification, Characterization, and Molecular Cloning of a Novel Keratan Sulfate Hydrolase, Endo-β-N-acetylglucosaminidase, from Bacillus circulans. J. Biol. Chem. 2003, 278, 25766–25772. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shetty, A.K.; Sugahara, K. Neuritogenic Activity of Chondroitin/Dermatan Sulfate Hybrid Chains of Embryonic Pig Brain and Their Mimicry from Shark Liver. J. Biol. Chem. 2007, 282, 2956–2966. [Google Scholar] [CrossRef]
- Bitter, T.; Muir, H.M. A Modified Uronic Acid Carbazole Reaction. Anal. Biochem. 1962, 4, 330–334. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, A.; Sugahara, K. Microanalysis of Glycosaminoglycan-Derived Oligosaccharides Labeled with a Fluorophore 2-Aminobenzamide by High-Performance Liquid Chromatography: Application to Disaccharide Composition Analysis and Exosequencing of Oligosaccharides. Anal. Biochem. 1999, 269, 367–378. [Google Scholar] [CrossRef]
- Garg, H.G.; Mrabat, H.; Yu, L.; Hales, C.A.; Li, B.; Moore, C.N.; Zhang, F.; Linhardt, R.J. Anti-Proliferative Effects of O-acyl-low-molecular-weight Heparin Derivatives on Bovine Pulmonary Artery Smooth Muscle Cells. Glycoconj. J. 2011, 28, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Li, Q.; Lv, Y.; Wang, Q.C.; Zhao, X. Preparation and Structural Characterization of Regioselective 4-O/6-O-Desulfated Chondroitin Sulfate. Carbohydr. Res. 2018, 460, 8–13. [Google Scholar] [CrossRef]
- Cai, C.; Solakyildirim, K.; Yang, B.; Beaudet, J.M.; Weyers, A.; Linhardt, R.J.; Zhang, F. Semi-Synthesis of Chondroitin Sulfate-E from Chondroitin Sulfate-A. Carbohydr. Polym. 2012, 87, 822–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucci, A.; Schenetti, L.; Volpi, N. 1H and 13C Nuclear Magnetic Resonance Identification and Characterization of Components of Chondroitin Sulfates of Various Origin. Carbohydr. Polym. 2000, 41, 37–45. [Google Scholar] [CrossRef]
- Kusche-Gullberg, M.; Kjellén, L. Sulfotransferases in Glycosaminoglycan Biosynthesis. Curr. Opin. Struct. Biol. 2003, 13, 605–611. [Google Scholar] [CrossRef]
- Izumikawa, T.; Kitagawa, H.; Mizuguchi, S.; Nomura, K.H.; Nomura, K.; Tamura, J.I.; Gengyo-Ando, K.; Mitani, S.; Sugahara, K. Nematode Chondroitin Polymerizing Factor Showing Cell-/Organ-Specific Expression Is Indispensable for Chondroitin Synthesis and Embryonic Cell Division. J. Biol. Chem. 2004, 279, 53755–53761. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Heinegård, D.; Malmström, A.; Schmidtchen, A.; Yoshida, K.; Fransson, L.A. Patterns of Uronosyl Epimerization and 4-/6-O-Sulphation in Chondroitin/Dermatan Sulphate from Decorin and Biglycan of Various Bovine Tissues. Glycobiology 1994, 4, 685–696. [Google Scholar] [CrossRef]
- Restaino, O.F.; Schiraldi, C. Chondroitin Sulfate: Are the Purity and the Structural Features Well Assessed? A Review on the Analytical Challenges. Carbohydr. Polym. 2022, 292, 119690. [Google Scholar] [CrossRef]
- Volpi, N. Quality of Different Chondroitin Sulfate Preparations in Relation to Their Therapeutic Activity. J. Pharm. Pharmacol. 2009, 61, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Volpi, N. Analytical Aspects of Pharmaceutical Grade Chondroitin Sulfates. J. Pharm. Sci. 2007, 96, 3168–3180. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Q.; Jiao, R.; Xu, Y.; Han, N.; Wang, W.; Zhu, C.; Li, F. A Novel Chondroitin Sulfate E from Dosidicus gigas Cartilage and Its Antitumor Metastatic Activity. Carbohydr. Polym. 2021, 262, 117971. [Google Scholar] [CrossRef] [PubMed]
- Scully, M.F.; Ellis, V.; Seno, N.; Kakkar, V.V. The Anticoagulant Properties of Mast Cell Product, Chondroitin Sulphate E. Biochem. Biophys. Res. Commun. 1986, 137, 15–22. [Google Scholar] [CrossRef]
- Bjornsson, T.D.; Nash, P.V.; Schaten, R. The Anticoagulant Effect of Chondroitin-4-Sulfate. Thromb. Res. 1982, 27, 15–21. [Google Scholar] [CrossRef]
- Ben Mansour, M.; Balti, R.; Ollivier, V.; Ben Jannet, H.; Chaubet, F.; Maaroufi, R.M. Characterization and Anticoagulant Activity of a Fucosylated Chondroitin Sulfate with Unusually Procoagulant Effect from Sea Cucumber. Carbohydr. Polym. 2017, 174, 760–771. [Google Scholar] [CrossRef]
- Fonseca, R.J.; Mourão, P.A. Fucosylated Chondroitin Sulfate as a New Oral Antithrombotic Agent. Thromb Haemost. 2006, 96, 822–829. [Google Scholar] [CrossRef]
- Krichen, F.; Bougatef, H.; Sayari, N.; Capitani, F.; Ben Amor, I.; Koubaa, I.; Maccari, F.; Mantovani, V.; Galeotti, F.; Volpi, N.; et al. Isolation, Purification and Structural Characterestics of Chondroitin Sulfate from Smooth Hound Cartilage: In Vitro Anticoagulant and Antiproliferative Properties. Carbohydr. Polym. 2018, 197, 451–459. [Google Scholar] [CrossRef]
- Dong, F.; Quan, X.; Wang, Q.; Liu, Z.; Cui, T.; Wang, W.; Tang, D.; Zhang, R.; Zhang, C.; Wang, H.; et al. Purification, Structural Characterization, and Anticoagulant Activity Evaluation of Chondroitin Sulfate from Codfish (Gadus macrocephalus) Bones. Int. J. Biol. Macromol. 2022, 210, 759–767. [Google Scholar] [CrossRef]
- Sie, P.; Ofosu, F.; Fernandez, F.; Buchanan, M.R.; Petitou, M.; Boneu, B. Respective Role of Antithrombin III and Heparin Cofactor II in the in vitro Anticoagulant Effect of Heparin and of Various Sulphated Polysaccharides. Br. J. Haematol. 1986, 64, 707–714. [Google Scholar] [CrossRef]
- Gui, M.; Song, J.; Zhang, L.; Wang, S.; Wu, R.; Ma, C.; Li, P. Chemical Characteristics and Antithrombotic Effect of Chondroitin Sulfates from Sturgeon Skull and Sturgeon Backbone. Carbohydr. Polym. 2015, 123, 454–460. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CS Samples | SHCS (%) | SBCS (%) | SFCS (%) | Total (%) |
|---|---|---|---|---|
| Dry weight of cartilage (g) | 4.87 (56.7) | 1.86 (21.6) | 1.87 (21.7) | 8.60 (100) |
| Purified CS (g) | 1.30 (53.3) | 0.58 (23.8) | 0.56 (22.9) | 2.44 (100) |
| Purity (%) | 96.0 | 98.59 | 96.63 | - |
| Yield (%) | 25.63 | 30.74 | 28.94 | - |
| CS Samples | Mn a | Mw b | Mz c | Mw/Mn d |
|---|---|---|---|---|
| kDa | ||||
| SHCS | 19.6 (0.4%) e | 21.0 (0.4%) | 22.7 (0.4%) | 1.07 |
| SBCS | 18.4 (0.4%) | 20.2 (0.4%) | 21.8 (0.4%) | 1.07 |
| SFCS | 25.2 (0.4%) | 26.5 (0.4%) | 28.4 (0.4%) | 1.05 |
| CS Samples | SHCS | SBCS | SFCS |
|---|---|---|---|
| mol % | |||
| ΔO unit | 7.20 | 5.60 | 5.37 |
| ΔC unit | 56.56 | 44.08 | 39.70 |
| ΔA unit | 36.24 | 50.32 | 54.93 |
| Total | 100 | 100 | 100 |
| S/unit | 0.93 | 0.94 | 0.95 |
| Oligosaccharides | SHCS | SBCS | SFCS |
|---|---|---|---|
| mol % | |||
| Tetrasccharide | 26.55 | 28.41 | 32.77 |
| Hexasaccharider | 37.88 | 38.59 | 39.16 |
| Octasaccharide | 22.48 | 20.70 | 17.71 |
| Decasaccharide | 13.07 | 12.30 | 9.85 |
| Total | 100 | 100 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, M.; Qin, Y.; Fan, Y.; Wang, X.; Yi, H.; Cui, X.; Li, F.; Wang, W. Structural Characterization and Glycosaminoglycan Impurities Analysis of Chondroitin Sulfate from Chinese Sturgeon. Polymers 2022, 14, 5311. https://doi.org/10.3390/polym14235311
Zhao M, Qin Y, Fan Y, Wang X, Yi H, Cui X, Li F, Wang W. Structural Characterization and Glycosaminoglycan Impurities Analysis of Chondroitin Sulfate from Chinese Sturgeon. Polymers. 2022; 14(23):5311. https://doi.org/10.3390/polym14235311
Chicago/Turabian StyleZhao, Mei, Yong Qin, Ying Fan, Xu Wang, Haixin Yi, Xiaoyu Cui, Fuchuan Li, and Wenshuang Wang. 2022. "Structural Characterization and Glycosaminoglycan Impurities Analysis of Chondroitin Sulfate from Chinese Sturgeon" Polymers 14, no. 23: 5311. https://doi.org/10.3390/polym14235311