
Controlled and Synchronised Vascular Regeneration upon the Implantation of Iloprost- and Cationic Amphiphilic Drugs-Conjugated Tissue-Engineered Vascular Grafts into the Ovine Carotid Artery: A Proteomics-Empowered Study

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fabrication of TEVGs
2.2. Anti-Thrombotic and Antimicrobial Modification and Sterilisation of TEVGs
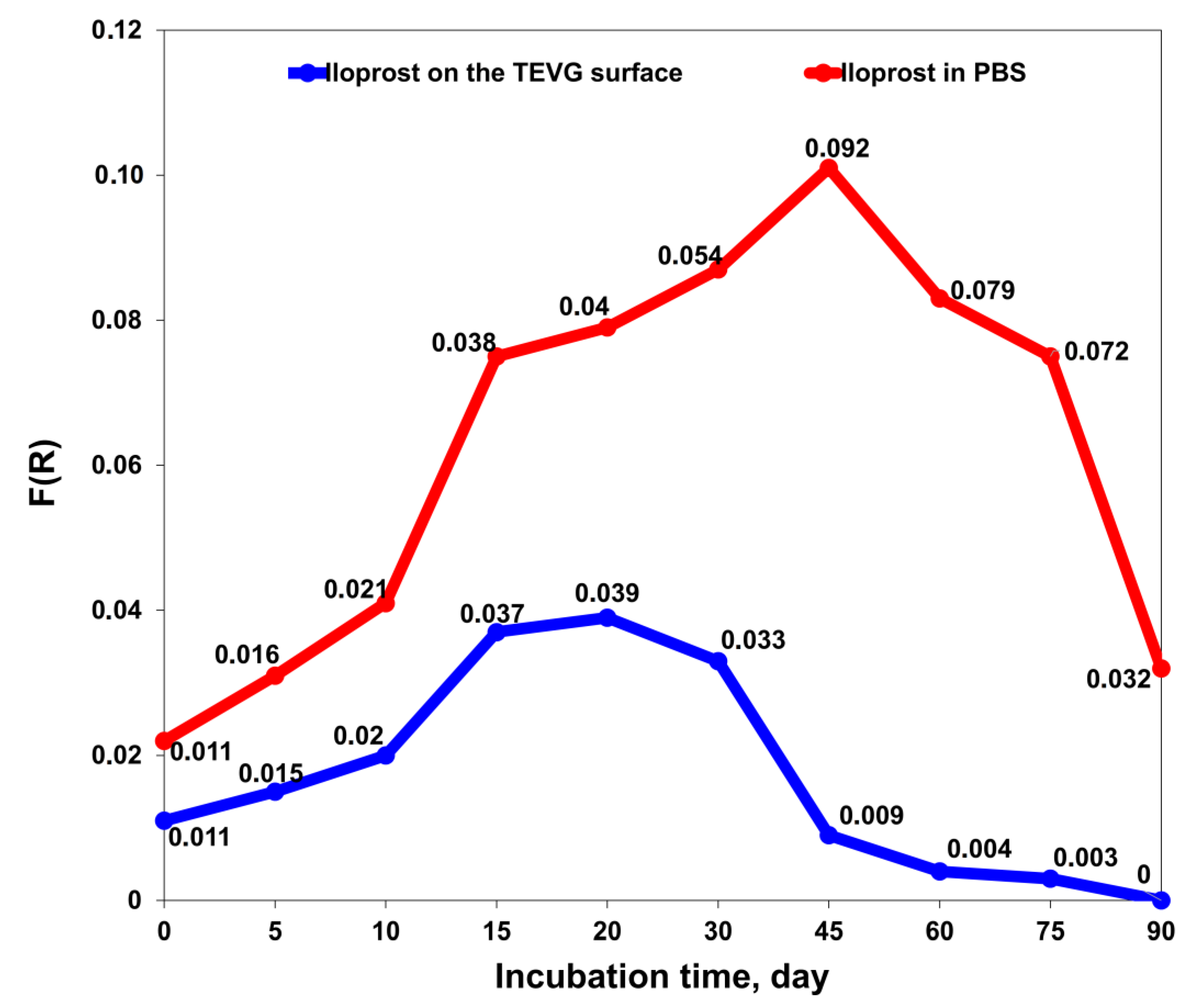
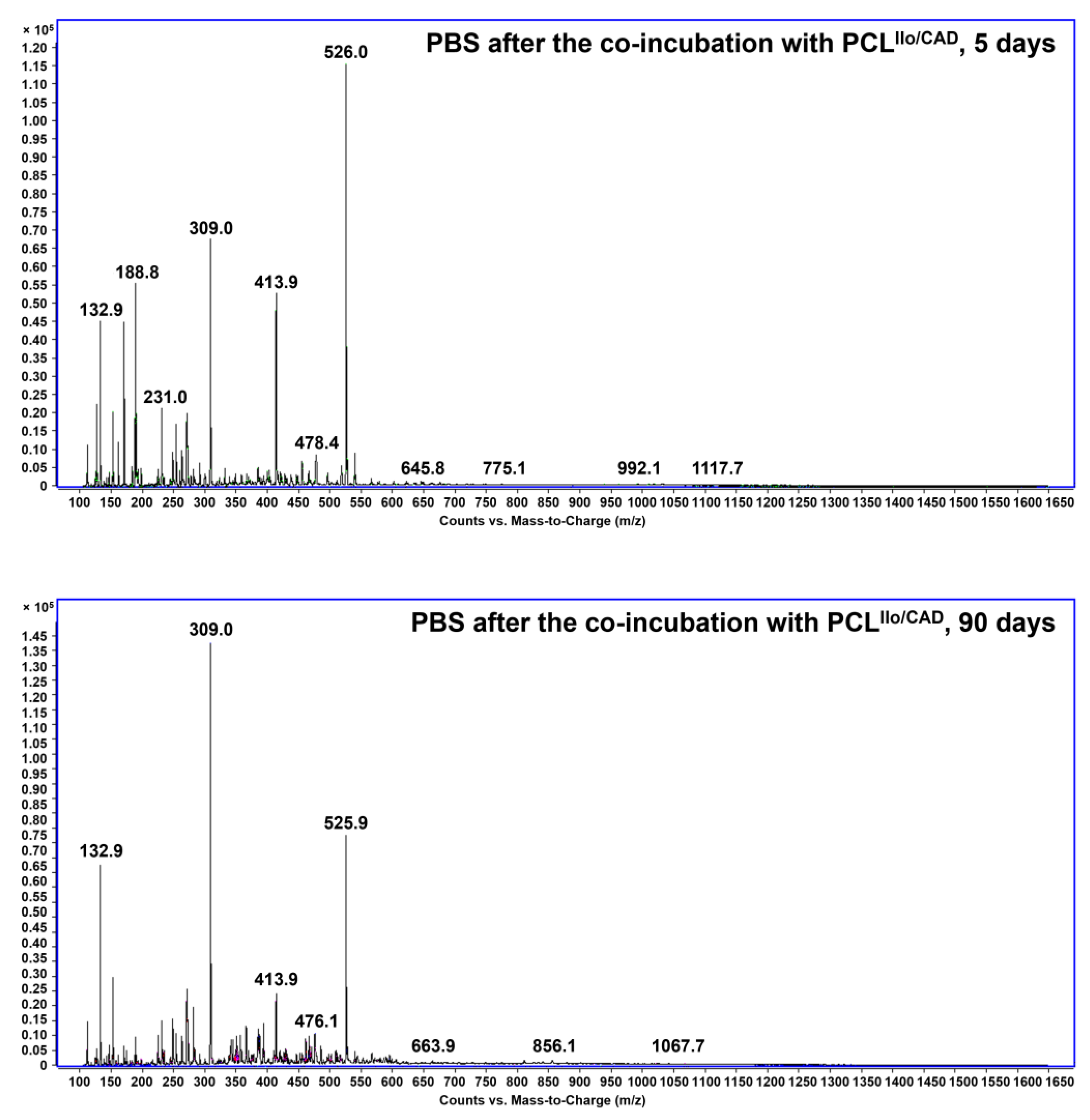
2.3. Conjugation Verification and Controlled Release Evaluation
2.4. Implantation of TEVGs into the Ovine Carotid Artery
2.5. Histological Examination
2.6. Immunostaining
2.7. Ultrastructural Analysis
2.8. Proteomic Profiling
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
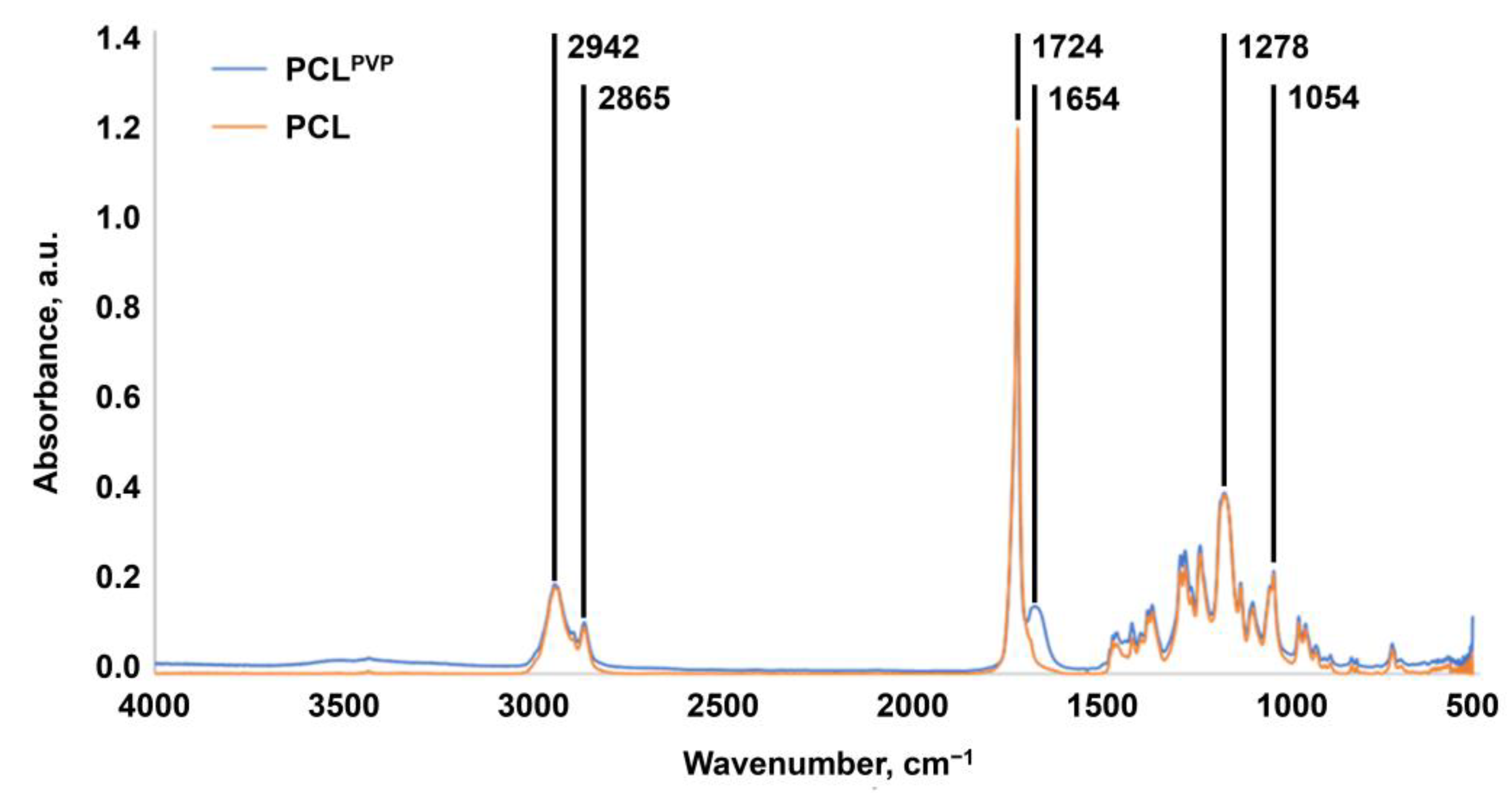
3.1. Biodegradable Electrospun TEVGs Can Be Reinforced with Anti-Aneurysmatic PCL Sheath and Modified with Iloprost and 1,5-bis-(4-tetradecyl-1,4-diazoniabicyclo [2.2.2] octan-1-yl) Pentane Tetrabromide
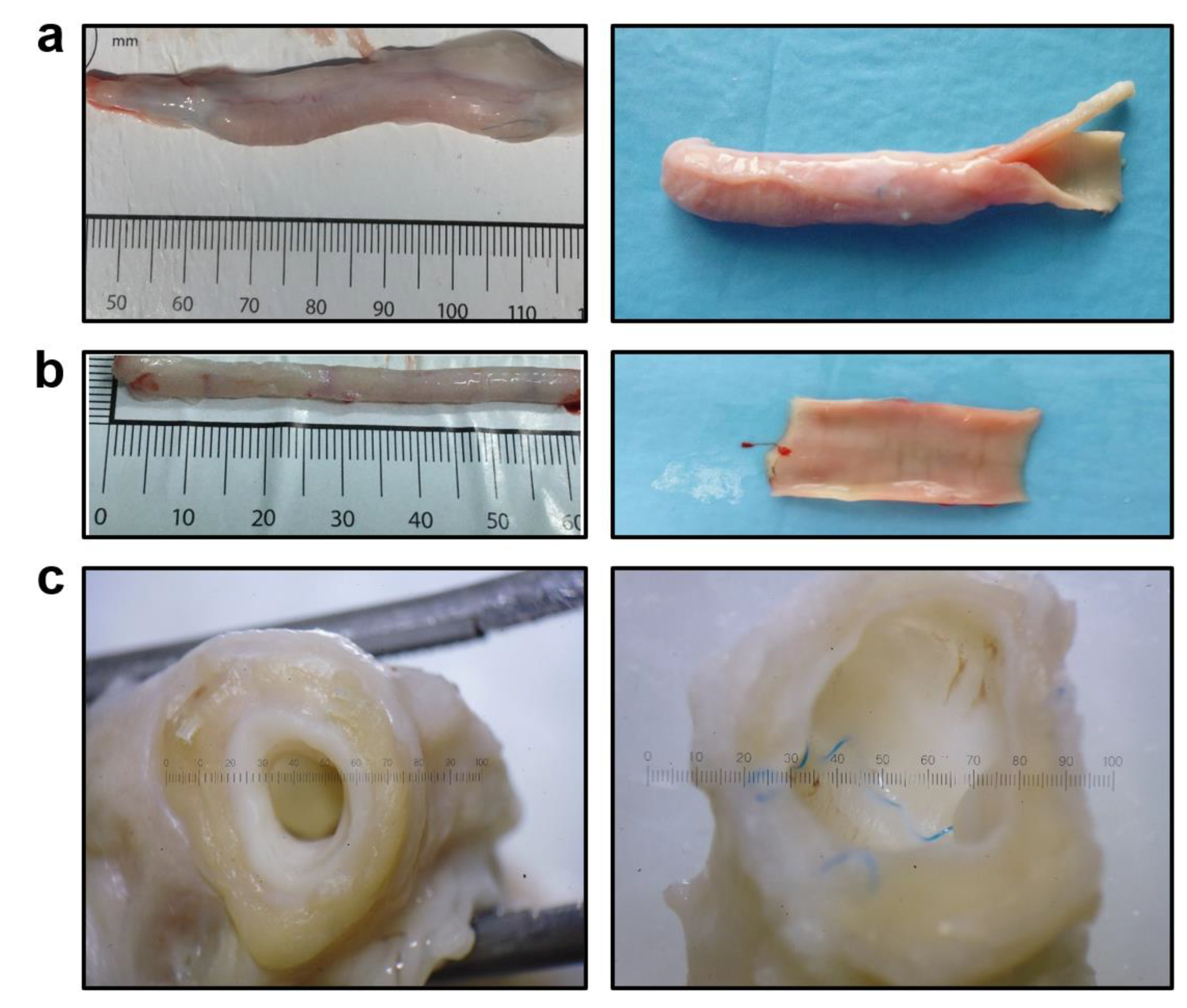
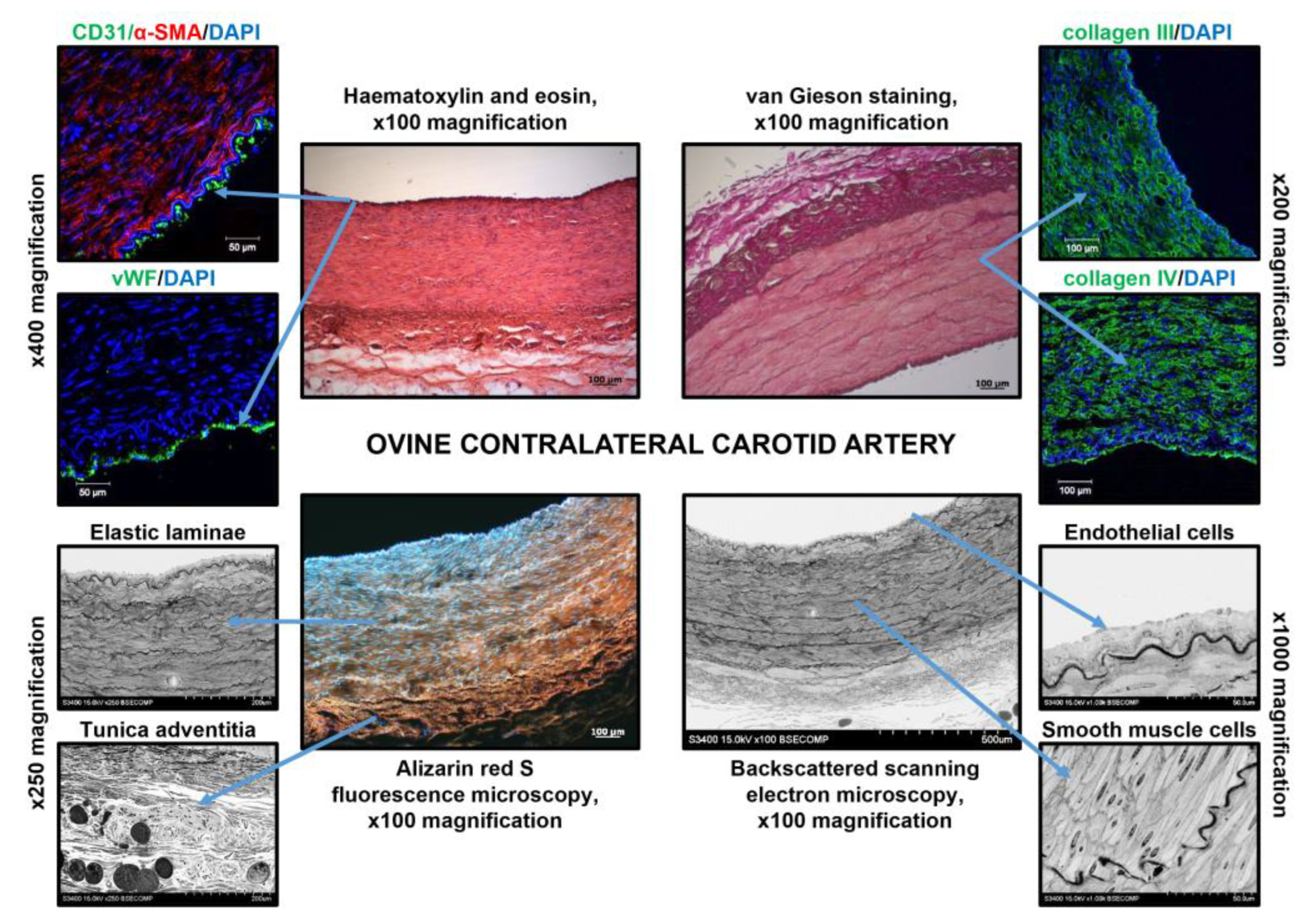
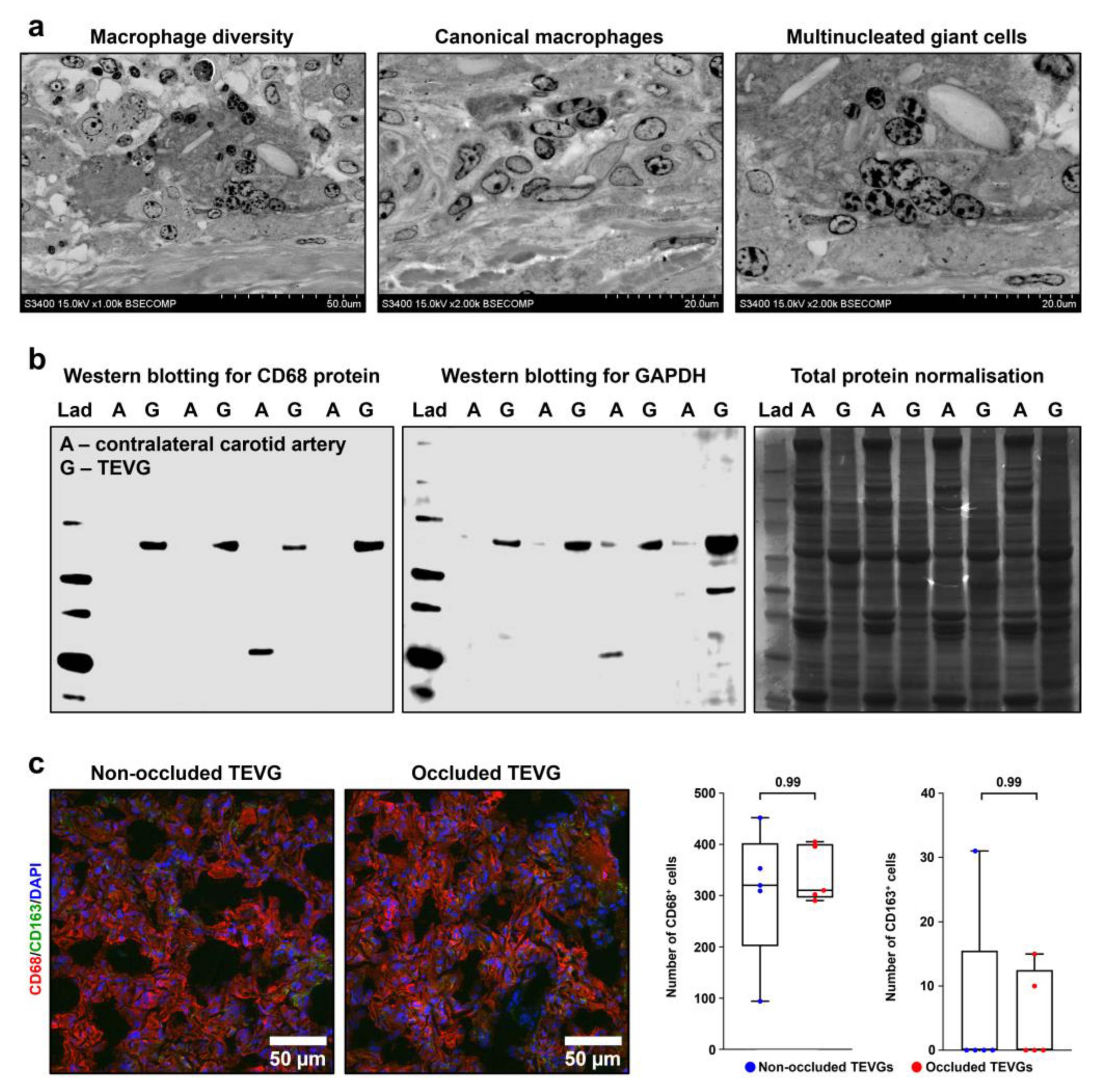
3.2. PCLIlo/CAD TEVGs Are Endothelialised and Show Hierarchical Structure 6 Months upon the Implantation into the Ovine Carotid Artery
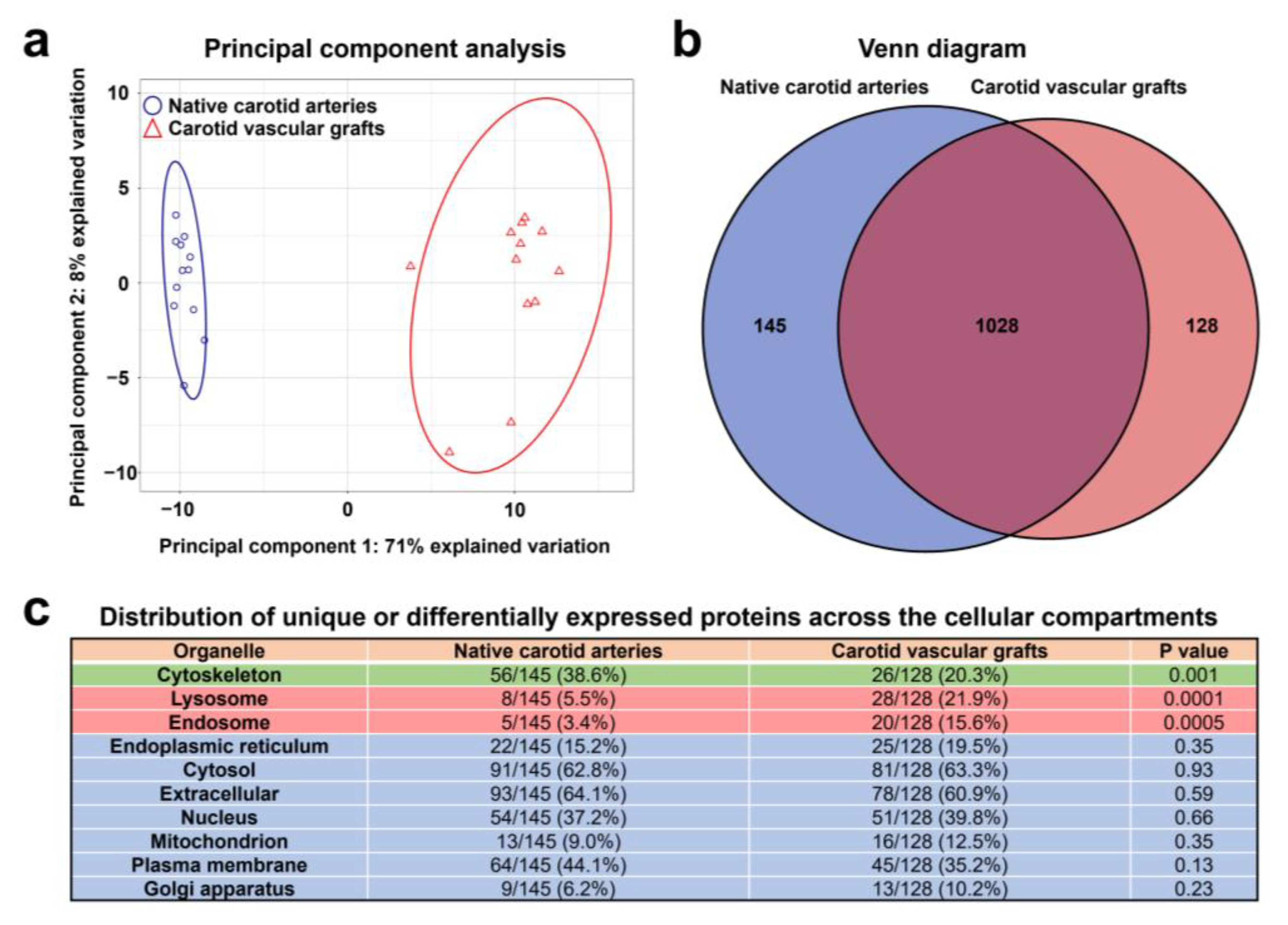
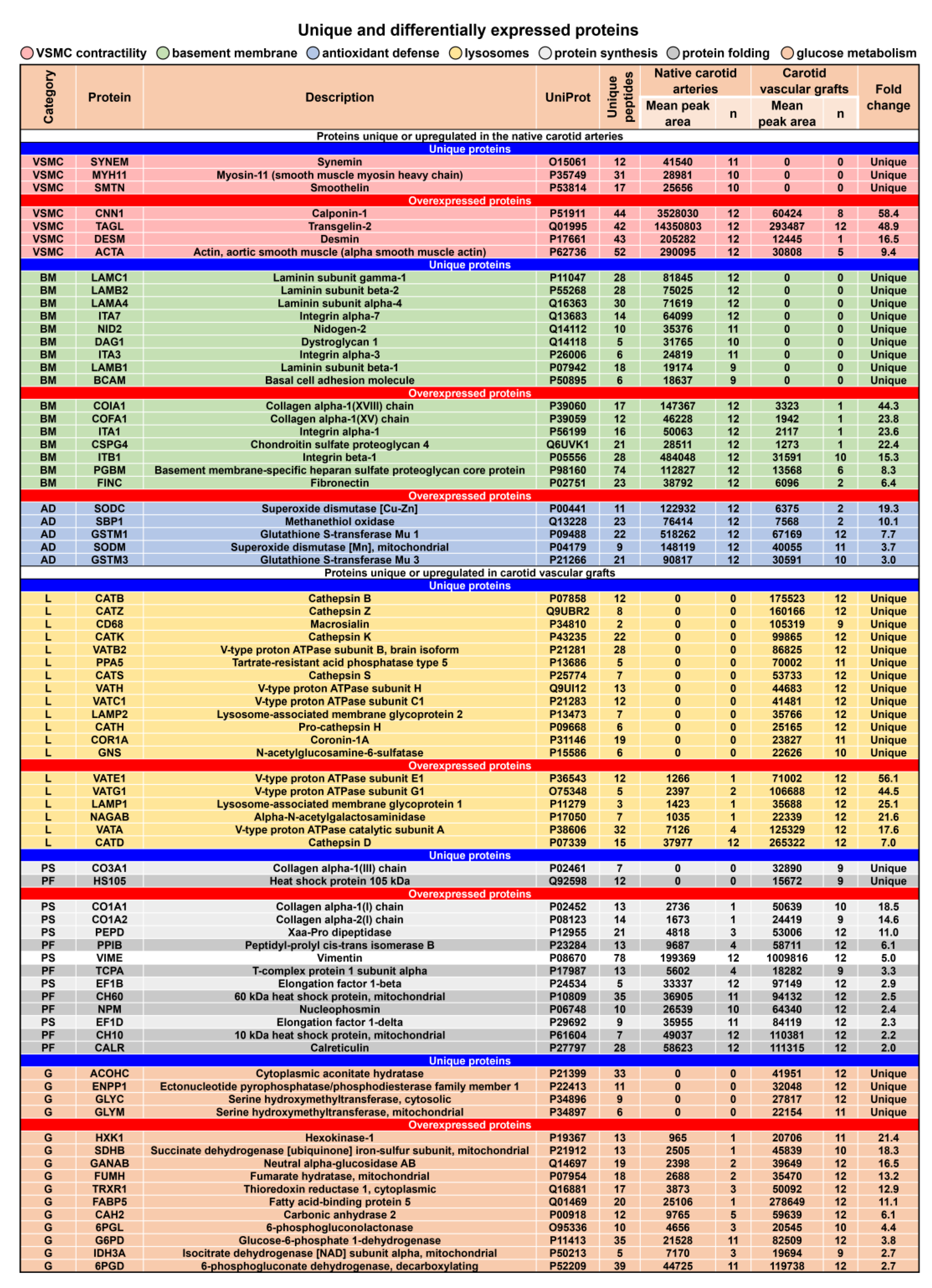
3.3. Proteomic Profiling of PCLIlo/CAD TEVGs and Contralateral Carotid Arteries Reveal Molecular Signatures of TEVG Remodeling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naegeli, K.M.; Kural, M.H.; Li, Y.; Wang, J.; Hugentobler, E.A.; Niklason, L.E. Bioengineering Human Tissues and the Future of Vascular Replacement. Circ. Res. 2022, 131, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, R.; Shoji, T.; Matsuzaki, Y.; Ulziibayar, A.; Shinoka, T. Current status of developing tissue engineering vascular technologies. Expert Opin. Biol. Ther. 2021, 22, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Durán-Rey, D.; Crisóstomo, V.; Sánchez-Margallo, J.A.; Sánchez-Margallo, F.M. Systematic Review of Tissue-Engineered Vascular Grafts. Front. Bioeng. Biotechnol. 2021, 9, 771400. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Ellman, D.; Andersen, D. Review: Tissue Engineering of Small-Diameter Vascular Grafts and Their In Vivo Evaluation in Large Animals and Humans. Cells 2021, 10, 713. [Google Scholar] [CrossRef]
- Kitsuka, T.; Hama, R.; Ulziibayar, A.; Matsuzaki, Y.; Kelly, J.; Shinoka, T. Clinical Application for Tissue Engineering Focused on Materials. Biomedicines 2022, 10, 1439. [Google Scholar] [CrossRef]
- Moore, M.J.; Tan, R.P.; Yang, N.; Rnjak-Kovacina, J.; Wise, S.G. Bioengineering artificial blood vessels from natural materials. Trends Biotechnol. 2022, 40, 693–707. [Google Scholar] [CrossRef]
- Weekes, A.; Bartnikowski, N.; Pinto, N.; Jenkins, J.; Meinert, C.; Klein, T.J. Biofabrication of small diameter tissue-engineered vascular grafts. Acta Biomater. 2022, 138, 92–111. [Google Scholar] [CrossRef]
- Rickel, A.P.; Deng, X.; Engebretson, D.; Hong, Z. Electrospun nanofiber scaffold for vascular tissue engineering. Mater. Sci. Eng. C 2021, 129, 112373. [Google Scholar] [CrossRef]
- Blum, K.M.; Mirhaidari, G.J.; Breuer, C.K. Tissue engineering: Relevance to neonatal congenital heart disease. Semin. Fetal Neonatal Med. 2022, 27, 101225. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Wiet, M.; Boe, B.; Shinoka, T. The Real Need for Regenerative Medicine in the Future of Congenital Heart Disease Treatment. Biomedicines 2021, 9, 478. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, F.; Guo, Z.; Zhao, Q. Tissue-engineered vascular grafts and regeneration mechanisms. J. Mol. Cell. Cardiol. 2021, 165, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Chen, C.; Liu, Y.; Lu, T.; Wu, Z. Strategies in cell-free tissue-engineered vascular grafts. J. Biomed. Mater. Res. Part A 2020, 108, 426–445. [Google Scholar] [CrossRef]
- Niklason, L.E.; Lawson, J.H. Bioengineered human blood vessels. Science 2020, 370, eaaw8682. [Google Scholar] [CrossRef] [PubMed]
- Toong, D.W.Y.; Toh, H.W.; Ng, J.C.K.; Wong, P.E.H.; Leo, H.L.; Venkatraman, S.; Tan, L.P.; Ang, H.Y.; Huang, Y. Bioresorbable Polymeric Scaffold in Cardiovascular Applications. Int. J. Mol. Sci. 2020, 21, 3444. [Google Scholar] [CrossRef]
- Stowell, C.E.; Wang, Y. Quickening: Translational design of resorbable synthetic vascular grafts. Biomaterials 2018, 173, 71–86. [Google Scholar] [CrossRef]
- Schmitt, P.R.; Dwyer, K.D.; Coulombe, K.L.K. Current Applications of Polycaprolactone as a Scaffold Material for Heart Regeneration. ACS Appl. Bio. Mater. 2022, 5, 2461–2480. [Google Scholar] [CrossRef] [PubMed]
- Backes, E.H.; Harb, S.V.; Beatrice, C.A.G.; Shimomura, K.M.B.; Passador, F.R.; Costa, L.C.; Pessan, L.A. Polycaprolactone usage in additive manufacturing strategies for tissue engineering applications: A review. J. Biomed. Mater. Res. Part B Appl. Biomater. 2022, 110, 1479–1503. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, X.; Yang, L.; Sun, L.; Mu, S.; Zong, H.; Li, Q.; Wang, F.; Song, S.; Yang, C.; et al. Evaluation of remodeling and regeneration of electrospun PCL/fibrin vascular grafts in vivo. Mater. Sci. Eng. C 2021, 118, 111441. [Google Scholar] [CrossRef] [PubMed]
- Fukunishi, T.; Best, C.A.; Sugiura, T.; Shoji, T.; Yi, T.; Udelsman, B.; Ohst, D.; Ong, C.S.; Zhang, H.; Shinoka, T.; et al. Tissue-Engineered Small Diameter Arterial Vascular Grafts from Cell-Free Nanofiber PCL/Chitosan Scaffolds in a Sheep Model. PLoS ONE 2016, 11, e0158555. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Wang, L.; Zhang, Q.; Dong, X.; Zhu, T.; Lu, S. Electrospun PCL/collagen hybrid nanofibrous tubular graft based on post-network bond processing for vascular substitute. Biomater. Adv. 2022, 139, 213031. [Google Scholar] [CrossRef]
- Swartz, D.D.; Andreadis, S.T. Animal models for vascular tissue-engineering. Curr. Opin. Biotechnol. 2013, 24, 916–925. [Google Scholar] [CrossRef] [Green Version]
- Thomas, L.V.; Lekshmi, V.; Nair, P.D. Tissue engineered vascular grafts—preclinical aspects. Int. J. Cardiol. 2013, 167, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Liesenborghs, L.; Meyers, S.; Vanassche, T.; Verhamme, P. Coagulation: At the heart of infective endocarditis. J. Thromb. Haemost. 2020, 18, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Polzin, A.; Dannenberg, L.; M’Pembele, R.; Mourikis, P.; Naguib, D.; Zako, S.; Helten, C.; Petzold, T.; Levkau, B.; Hohlfeld, T.; et al. Staphylococcus aureus increases platelet reactivity in patients with infective endocarditis. Sci. Rep. 2022, 12, 12933. [Google Scholar] [CrossRef] [PubMed]
- Antonova, L.; Krivkina, E.; Sevostianova, V.; Mironov, A.; Rezvova, M.; Shabaev, A.; Tkachenko, V.; Krutitskiy, S.; Khanova, M.; Sergeeva, T.; et al. Tissue-Engineered Carotid Artery Interposition Grafts Demonstrate High Primary Patency and Promote Vascular Tissue Regeneration in the Ovine Model. Polymers 2021, 13, 2637. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Miyamoto, S.; Miyachi, H.; Iwaki, R.; Shoji, T.; Blum, K.; Chang, Y.-C.; Kelly, J.; Reinhardt, J.W.; Nakayama, H.; et al. Improvement of a Novel Small-diameter Tissue-engineered Arterial Graft with Heparin Conjugation. Ann. Thorac. Surg. 2021, 111, 1234–1241. [Google Scholar] [CrossRef]
- Matsuzaki, Y.; Iwaki, R.; Reinhardt, J.W.; Chang, Y.-C.; Miyamoto, S.; Kelly, J.; Zbinden, J.; Blum, K.; Mirhaidari, G.; Ulziibayar, A.; et al. The effect of pore diameter on neo-tissue formation in electrospun biodegradable tissue-engineered arterial grafts in a large animal model. Acta Biomater. 2020, 115, 176–184. [Google Scholar] [CrossRef]
- Elliott, M.B.; Matsushita, H.; Shen, J.; Yi, J.; Inoue, T.; Brady, T.; Santhanam, L.; Mao, H.Q.; Hibino, N.; Gerecht, S. Off-the-shelf, heparinized small diameter vascular graft limits acute thrombogenicity in a porcine model. Acta Biomater. 2022, 151, 134–147. [Google Scholar] [CrossRef]
- Blum, K.M.; Zbinden, J.C.; Ramachandra, A.B.; Lindsey, S.E.; Szafron, J.M.; Reinhardt, J.W.; Heitkemper, M.; Best, C.A.; Mirhaidari, G.J.M.; Chang, Y.-C.; et al. Tissue engineered vascular grafts transform into autologous neovessels capable of native function and growth. Commun. Med. 2022, 2, 3. [Google Scholar] [CrossRef]
- Fang, S.; Riber, S.S.; Hussein, K.; Ahlmann, A.H.; Harvald, E.B.; Khan, F.; Beck, H.C.; Weile, L.K.K.; Sørensen, J.A.; Sheikh, S.P.; et al. Decellularized human umbilical artery: Biocompatibility and in vivo functionality in sheep carotid bypass model. Mater. Sci. Eng. C 2020, 112, 110955. [Google Scholar] [CrossRef]
- Mallis, P.; Sokolis, D.P.; Makridakis, M.; Zoidakis, J.; Velentzas, A.D.; Katsimpoulas, M.; Vlahou, A.; Kostakis, A.; Stavropoulos-Giokas, C.; Michalopoulos, E. Insights into Biomechanical and Proteomic Characteristics of Small Diameter Vascular Grafts Utilizing the Human Umbilical Artery. Biomedicines 2020, 8, 280. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.H.; Enayati, M.; Grasl, C.; Walter, I.; Budinsky, L.; Zebic, G.; Kaun, C.; Wagner, A.; Kratochwill, K.; Redl, H.; et al. Acellular vascular matrix grafts from human placenta chorion: Impact of ECM preservation on graft characteristics, protein composition and in vivo performance. Biomaterials 2018, 177, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Yarinich, L.A.; Burakova, E.A.; Zakharov, B.A.; Boldyreva, E.V.; Babkina, I.N.; Tikunova, N.V.; Silnikov, V.N. Synthesis and structure–activity relationship of novel 1,4-diazabicyclo[2.2.2]octane derivatives as potent antimicrobial agents. Eur. J. Med. Chem. 2015, 95, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Antonova, L.V.; Silnikov, V.N.; Sevostyanova, V.V.; Yuzhalin, A.E.; Koroleva, L.S.; Velikanova, E.A.; Mironov, A.V.; Godovikova, T.S.; Kutikhin, A.G.; Glushkova, T.V.; et al. Biocompatibility of Small-Diameter Vascular Grafts in Different Modes of RGD Modification. Polymers 2019, 11, 174. [Google Scholar] [CrossRef] [Green Version]
- Mukhamadiyarov, R.A.; Bogdanov, L.A.; Glushkova, T.V.; Shishkova, D.K.; Kostyunin, A.E.; Koshelev, V.A.; Shabaev, A.R.; Frolov, A.V.; Stasev, A.N.; Lyapin, A.A.; et al. EMbedding and Backscattered Scanning Electron Microscopy: A Detailed Protocol for the Whole-Specimen, High-Resolution Analysis of Cardiovascular Tissues. Front. Cardiovasc. Med. 2021, 8, 739549. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.; Tibshirani, R.; Narasimhan, B.; Chu, G. Impute: Impute: Imputation for Microarray Data, R package version 1.70.0; 2022. Available online: https://bioconductor.org/packages/release/bioc/html/impute.html (accessed on 17 November 2022).
- Ritchie, M.E.; Belinda, P.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling, R package version 1.14.0; 2022. Available online: https://bioconductor.org/packages/devel/bioc/vignettes/EnhancedVolcano/inst/doc/EnhancedVolcano.html (accessed on 17 November 2022).
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef] [PubMed]
- Binder, J.X.; Pletscher-Frankild, S.; Tsafou, K.; Stolte, C.; O’Donoghue, S.; Schneider, R.; Jensen, L.J. Compartments: Unification and visualization of protein subcellular localization evidence. Database 2014, 2014, bau012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Fabregat, A.; Jupe, S.; Matthews, L.; Sidiropoulos, K.; Gillespie, M.; Garapati, P.; Haw, R.; Jassal, B.; Korninger, F.; May, B.; et al. The Reactome Pathway Knowledgebase. Nucleic Acids Res. 2018, 46, D649–D655. [Google Scholar] [CrossRef] [PubMed]
- Griss, J.; Viteri, G.; Sidiropoulos, K.; Nguyen, V.; Fabregat, A.; Hermjakob, H. ReactomeGSA—Efficient Multi-Omics Comparative Pathway Analysis. Mol. Cell. Proteom. 2020, 19, 2115–2125. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Moezinia, C.J.; Ji-Xu, A.; Azari, A.; Horlick, S.; Denton, C.; Stratton, R. Iloprost for COVID-19-related vasculopathy. Lancet Rheumatol. 2020, 2, e582–e583. [Google Scholar] [CrossRef]
- Faggioli, P.M.; Mumoli, N.; Mazzone, A. Iloprost in COVID-19: The Rationale of Therapeutic Benefit. Front. Cardiovasc. Med. 2021, 8, 649499. [Google Scholar] [CrossRef]
- Musiał, J.; Wilczyńska, M.; Sładek, K.; Cierniewski, C.; Nizankowski, R.; Szczeklik, A. Fibrinolytic activity of prostacyclin and iloprost in patients with peripheral arterial disease. Prostaglandins 1986, 31, 61–70. [Google Scholar] [CrossRef]
- Bertelé, V.; Mussoni, L.; del Rosso, G.; Pintucci, G.; Carriero, M.R.; Merati, M.G.; Libretti, A.; de Gaetano, G. Defective fibrinolytic response in atherosclerotic patients—effect of iloprost and its possible mechanism of action. Thromb. Haemost. 1988, 60, 141–144. [Google Scholar] [PubMed]
- Bertelé, V.; Mussoni, L.; Pintucci, G.; del Rosso, G.; Romano, G.; de Gaetano, G.; Libretti, A. The Inhibitory Effect of Aspirin on Fibrinolysis Is Reversed by Iloprost, a Prostacyclin Analogue. Thromb. Haemost. 1989, 61, 286–288. [Google Scholar] [CrossRef]
- Iacoviello, L.; De Curtis, A.; D’Adamo, M.C.; Amore, C.; Buczko, W.; De Gaetano, G.; Donati, M.B. Prostacyclin is required for t-PA release after venous occlusion. Am. J. Physiol. Circ. Physiol. 1994, 266, H429–H434. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, A.A.; Azzami, K.; Ryabchikova, E.I.; Spitsyna, Y.E.; Silnikov, V.N.; Ritter, W.; Gross, H.J.; Tautz, J.; Vlassov, V.; Beier, H.; et al. Inactivation of a non-enveloped RNA virus by artificial ribonucleases: Honey bees and Acute bee paralysis virus as a new experimental model for in vivo antiviral activity assessment. Antivir. Res. 2011, 91, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, A.A.; Goncharova, E.P.; Koroleva, L.S.; Burakova, E.A.; Ryabchikova, E.I.; Bichenkova, E.V.; Silnikov, V.N.; Vlassov, V.V.; Zenkova, M.A. Artificial ribonucleases inactivate a wide range of viruses using their ribonuclease, membranolytic, and chaotropic-like activities. Antivir. Res. 2016, 133, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Burakova, E.A.; Saranina, I.V.; Tikunova, N.V.; Nazarkina, Z.K.; Laktionov, P.P.; Karpinskaya, L.A.; Anikin, V.B.; Zarubaev, V.V.; Silnikov, V.N. Biological evaluation of tetracationic compounds based on two 1,4-diazabicyclo[2.2.2]octane moieties connected by different linkers. Bioorg. Med. Chem. 2016, 24, 6012–6020. [Google Scholar] [CrossRef]
- Antonova, L.V.; Krivkina, E.O.; Rezvova, M.A.; Sevost’yanova, V.V.; Mironov, A.V.; Glushkova, T.V.; Klyshnikov, K.Y.; Ovcharenko, E.A.; Kudryavceva, Y.A.; Barbarash, L.S. Biodegradable vascular graft reinforced with a biodegradable sheath. Compl, Iss, Cardiovasc. Dis. 2019, 8, 87–97. [Google Scholar] [CrossRef]
- Koch, S.E.; de Kort, B.J.; Holshuijsen, N.; Brouwer, H.F.M.; van der Valk, D.C.; Dankers, P.Y.W.; van Luijk, J.A.K.R.; Hooijmans, C.R.; de Vries, R.B.M.; Bouten, C.V.C.; et al. Animal studies for the evaluation of in situ tissue-engineered vascular grafts — a systematic review, evidence map, and meta-analysis. NPJ Regen. Med. 2022, 7, 17. [Google Scholar] [CrossRef]
- Anes, E.; Pires, D.; Mandal, M.; Azevedo-Pereira, J.M. Spatial localization of cathepsins: Implications in immune activation and resolution during infections. Front. Immunol. 2022, 13, 955407. [Google Scholar] [CrossRef]
- Trivedi, P.C.; Bartlett, J.J.; Pulinilkunnil, T. Lysosomal Biology and Function: Modern View of Cellular Debris Bin. Cells 2020, 9, 1131. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, S.; Wang, M.; Shi, G.P. Cysteinyl cathepsins in cardiovascular diseases. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140360. [Google Scholar] [CrossRef] [PubMed]
- Bonito, V.; Koch, S.E.; Krebber, M.M.; Carvajal-Berrio, D.A.; Marzi, J.; Duijvelshoff, R.; Lurier, E.B.; Buscone, S.; Dekker, S.; Jong, S.M.J.; et al. Distinct Effects of Heparin and Interleukin-4 Functionalization on Macrophage Polarization and In Situ Arterial Tissue Regeneration Using Resorbable Supramolecular Vascular Grafts in Rats. Adv. Healthc. Mater. 2021, 10, e2101103. [Google Scholar] [CrossRef] [PubMed]
- Best, C.A.; Szafron, J.M.; Rocco, K.A.; Zbinden, J.; Dean, E.W.; Maxfield, M.W.; Kurobe, H.; Tara, S.; Bagi, P.S.; Udelsman, B.V.; et al. Differential outcomes of venous and arterial tissue engineered vascular grafts highlight the importance of coupling long-term implantation studies with computational modeling. Acta Biomater. 2019, 94, 183–194. [Google Scholar] [CrossRef]
- Fukunishi, T.; Best, C.A.; Sugiura, T.; Opfermann, J.; Ong, C.S.; Shinoka, T.; Breuer, C.K.; Krieger, A.; Johnson, J.; Hibino, N. Preclinical study of patient-specific cell-free nanofiber tissue-engineered vascular grafts using 3-dimensional printing in a sheep model. J. Thorac Cardiovasc. Surg. 2017, 153, 924–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurobe, H.; Maxfield, M.W.; Tara, S.; Rocco, K.A.; Bagi, P.S.; Yi, T.; Udelsman, B.; Zhuang, Z.W.; Cleary, M.; Iwakiri, Y.; et al. Development of Small Diameter Nanofiber Tissue Engineered Arterial Grafts. PLoS ONE 2015, 10, e0120328. [Google Scholar] [CrossRef] [Green Version]
- Fukunishi, T.; Ong, C.S.; Yesantharao, P.; Best, C.A.; Yi, T.; Zhang, H.; Mattson, G.; Boktor, J.; Nelson, K.; Shinoka, T.; et al. Different degradation rates of nanofiber vascular grafts in small and large animal models. J. Tissue Eng. Regen. Med. 2020, 14, 203–214. [Google Scholar] [CrossRef]
- Antonova, L.; Kutikhin, A.; Sevostianova, V.; Velikanova, E.; Matveeva, V.; Glushkova, T.; Mironov, A.; Krivkina, E.; Shabaev, A.; Senokosova, E.; et al. bFGF and SDF-1α Improve In Vivo Performance of VEGF-Incorporating Small-Diameter Vascular Grafts. Pharmaceuticals 2021, 14, 302. [Google Scholar] [CrossRef]
- Antonova, L.V.; Seifalian, A.M.; Kutikhin, A.G.; Sevostyanova, V.V.; Matveeva, V.G.; Velikanova, E.A.; Mironov, A.V.; Shabaev, A.R.; Glushkova, T.V.; Senokosova, E.A.; et al. Conjugation with RGD Peptides and Incorporation of Vascular Endothelial Growth Factor Are Equally Efficient for Biofunctionalization of Tissue-Engineered Vascular Grafts. Int. J. Mol. Sci. 2016, 17, 1920. [Google Scholar] [CrossRef] [Green Version]
- Antonova, L.V.; Sevostyanova, V.V.; Kutikhin, A.G.; Mironov, A.V.; Krivkina, E.O.; Shabaev, A.R.; Matveeva, V.G.; Velikanova, E.A.; Sergeeva, E.A.; Burago, A.Y.; et al. Vascular Endothelial Growth Factor Improves Physico-Mechanical Properties and Enhances Endothelialization of Poly(3-hydroxybutyrate-co-3-hydroxyvalerate)/Poly(ε-caprolactone) Small-Diameter Vascular Grafts In vivo. Front. Pharmacol. 2016, 7, 230. [Google Scholar] [CrossRef] [Green Version]
- Antonova, L.V.; Seifalian, A.M.; Kutikhin, A.G.; Sevostyanova, V.V.; Krivkina, E.O.; Mironov, A.V.; Burago, A.Y.; Velikanova, E.A.; Matveeva, V.G.; Glushkova, T.V.; et al. Bioabsorbable Bypass Grafts Biofunctionalised with RGD Have Enhanced Biophysical Properties and Endothelialisation Tested In vivo. Front. Pharmacol. 2016, 7, 136. [Google Scholar] [CrossRef]
- Wang, Z.; Mithieux, S.M.; Vindin, H.; Wang, Y.; Zhang, M.; Liu, L.; Zbinden, J.; Blum, K.M.; Yi, T.; Matsuzaki, Y.; et al. Rapid Regeneration of a Neoartery with Elastic Lamellae. Adv. Mater. 2022, 34, e2205614. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-L.; Szafron, J.M.; Blum, K.M.; Zbinden, J.C.; Khosravi, M.R.; Best, C.A.; Reinhardt, J.W.; Zheng, Q.; Yi, T.; Shinoka, T.; et al. Electrospun Tissue-Engineered Arterial Graft Thickness Affects Long-Term Composition and Mechanics. Tissue Eng. Part A 2021, 27, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Elliott, M.B.; Ginn, B.; Fukunishi, T.; Bedja, D.; Suresh, A.; Chen, T.; Inoue, T.; Dietz, H.C.; Santhanam, L.; Mao, H.-Q.; et al. Regenerative and durable small-diameter graft as an arterial conduit. Proc. Natl. Acad. Sci USA 2019, 116, 12710–12719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukunishi, T.; Lui, C.; Ong, C.S.; Dunn, T.; Xu, S.; Smoot, C.; Smalley, R.; Harris, J.; Gabriele, P.; Santhanam, L.; et al. Extruded poly (glycerol sebacate) and polyglycolic acid vascular graft forms a neoartery. J. Tissue Eng. Regen. Med. 2022, 16, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Bono, N.; Pezzoli, D.; Levesque, L.; Loy, C.; Candiani, G.; Fiore, G.; Mantovani, D. Unraveling the role of mechanical stimulation on smooth muscle cells: A comparative study between 2D and 3D models. Biotechnol. Bioeng. 2016, 113, 2254–2263. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.-J.; Liu, B.; Wang, H.-Q.; Yan, Z.-Q.; Shen, B.-R.; Jiang, Z.-L. Frequency-Dependent Phenotype Modulation of Vascular Smooth Muscle Cells under Cyclic Mechanical Strain. J. Vasc. Res. 2007, 44, 345–353. [Google Scholar] [CrossRef]
- Reusch, P.; Wagdy, H.; Reusch, R.; Wilson, E.; Ives, H.E. Mechanical Strain Increases Smooth Muscle and Decreases Nonmuscle Myosin Expression in Rat Vascular Smooth Muscle Cells. Circ. Res. 1996, 79, 1046–1053. [Google Scholar] [CrossRef]
- Fontana, L. Interventions to promote cardiometabolic health and slow cardiovascular ageing. Nat. Rev. Cardiol. 2018, 15, 566–577. [Google Scholar] [CrossRef]
- Fiuza-Luces, C.; Santos-Lozano, A.; Joyner, M.; Carrera-Bastos, P.; Picazo, O.; Zugaza, J.L.; Izquierdo, M.; Ruilope, L.M.; Lucia, A. Exercise benefits in cardiovascular disease: Beyond attenuation of traditional risk factors. Nat. Rev. Cardiol. 2018, 15, 731–743. [Google Scholar] [CrossRef]
- Marques, F.Z.; Mackay, C.R.; Kaye, D.M. Beyond gut feelings: How the gut microbiota regulates blood pressure. Nat. Rev. Cardiol. 2018, 15, 20–32. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Li, D.Y.; Hazen, S.L. Dietary metabolism, the gut microbiome, and heart failure. Nat. Rev. Cardiol. 2019, 16, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Chakaroun, R.M.; Olsson, L.M.; Bäckhed, F. The potential of tailoring the gut microbiome to prevent and treat cardiometabolic disease. Nat. Rev. Cardiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Nguyen, L.H.; Li, Y.; Yan, Y.; Ma, W.; Rinott, E.; Ivey, K.L.; Shai, I.; Willett, W.C.; Hu, F.B.; et al. The gut microbiome modulates the protective association between a Mediterranean diet and cardiometabolic disease risk. Nat. Med. 2021, 27, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, Z.; Kazemi, A.; Bartolomaeus, T.U.; Martami, F.; Noormohammadi, M.; Salari, A.; Löber, U.; Balou, H.A.; Forslund, S.K.; Mahdavi-Roshan, M. The effect of probiotic and synbiotic supplementation on lipid parameters among patients with cardiometabolic risk factors: A systematic review and meta-analysis of clinical trials. Cardiovasc. Res. 2022, 1–52. [Google Scholar] [CrossRef]
- Innocente, F.; Mandracchia, D.; Pektok, E.; Nottelet, B.; Tille, J.-C.; de Valence, S.; Faggian, G.; Mazzucco, A.; Kalangos, A.; Gurny, R.; et al. Paclitaxel-eluting biodegradable synthetic vascular prostheses: A step towards reduction of neointima formation? Circulation 2009, 120, S37–S45. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.S.; Buchen, J.T.; Pasquina, P.F.; Niklason, L.E.; Alvarez, L.M.; Jariwala, S.H. Critical Considerations for Regeneration of Vascularized Composite Tissues. Tissue Eng. Part B Rev. 2021, 27, 366–381. [Google Scholar] [CrossRef]
- Kutikhin, A.G.; Tupikin, A.E.; Matveeva, V.G.; Shishkova, D.K.; Antonova, L.V.; Kabilov, M.R.; Velikanova, E.A. Human Peripheral Blood-Derived Endothelial Colony-Forming Cells Are Highly Similar to Mature Vascular Endothelial Cells yet Demonstrate a Transitional Transcriptomic Signature. Cells 2020, 9, 876. [Google Scholar] [CrossRef] [Green Version]
- Kutikhin, A.G.; Shishkova, D.K.; Velikanova, E.A.; Sinitsky, M.Y.; Sinitskaya, A.V.; Markova, V.E. Endothelial Dysfunction in the Context of Blood-Brain Barrier Modeling. J. Evol. Biochem. Physiol. 2022, 58, 781–806. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
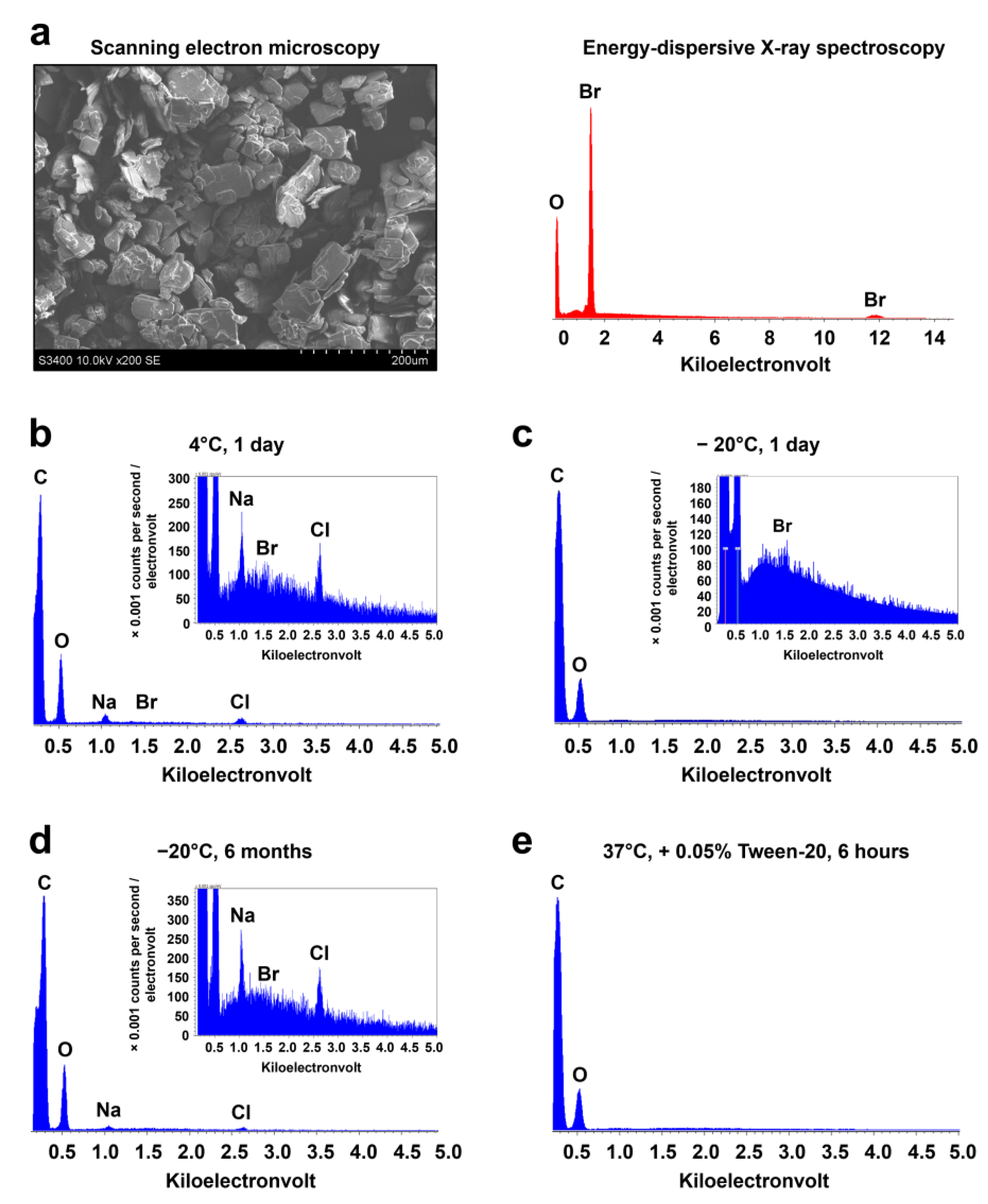
| Incubation time | T, °C | Conditions | Br (wt.%) | O (wt.%) | Na (wt.%) | Cl (wt.%) |
|---|---|---|---|---|---|---|
| 24 h | 5 | Without PBS | 2.84 | 97.16 | 0.0 | 0.0 |
| 24 h | −20 | Without PBS | 2.75 | 97.25 | 0.0 | 0.0 |
| 6 months | −20 | Without PBS | 1.39 | 93.62 | 3.82 | 1.17 |
| 24 h | 37 | PBS + 0.05% Tween-20 | 0.0 | 100.0 | 0.0 | 0.0 |
| Gene Ontology Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| Stress fiber | 72 | 17 | 33.8 | 4.11 × 10−18 |
| Contractile fiber | 244 | 31 | 18.2 | 2.36 × 10−26 |
| Basement membrane | 101 | 11 | 15.6 | 2.14 × 10−8 |
| Focal adhesion | 423 | 41 | 13.9 | 2.65 × 10−31 |
| Actin cytoskeleton | 510 | 37 | 10.4 | 4.78 × 10−24 |
| Collagen-containing extracellular matrix | 434 | 24 | 7.9 | 8.82 × 10−13 |
| Extracellular matrix | 575 | 24 | 6.0 | 2.50 × 10−10 |
| Extracellular vesicle | 2123 | 82 | 5.5 | 1.96 × 10−39 |
| Cytoskeleton | 2401 | 57 | 3.4 | 1.40 × 10−15 |
| Extracellular region | 4396 | 92 | 3.0 | 1.95 × 10−25 |
| Carotid vascular grafts | ||||
| Smooth endoplasmic reticulum | 32 | 3 | 15.1 | 2.59 × 10−2 |
| Endoplasmic reticulum-Golgi intermediate compartment | 135 | 6 | 7.1 | 6.11 × 10−3 |
| Collagen-containing extracellular matrix | 434 | 15 | 5.6 | 5.95 × 10−6 |
| Lysosome | 749 | 28 | 6.0 | 4.49 × 10−12 |
| Coated vesicle | 310 | 11 | 5.7 | 1.88 × 10−4 |
| Extracellular vesicle | 2123 | 70 | 5.3 | 5.60 × 10−32 |
| Extracellular matrix | 575 | 15 | 4.2 | 1.53 × 10−4 |
| Endosome | 1036 | 22 | 3.4 | 2.35 × 10−5 |
| Extracellular region | 4396 | 79 | 2.9 | 3.00 × 10−20 |
| Endoplasmic reticulum | 2045 | 29 | 2.3 | 6.96 × 10−4 |
| UniProtKB Keywords Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| Basement membrane | 7 | 4.6 | 22.0 | 3.2 × 10−6 |
| Extracellular matrix | 14 | 9.3 | 6.9 | 6.7 × 10−7 |
| Cytoskeleton | 41 | 27.2 | 4.2 | 4.0 × 10−14 |
| Cell junction | 24 | 15.9 | 3.7 | 6.1 × 10−7 |
| Cytoplasm | 88 | 58.3 | 2.2 | 4.1 × 10−16 |
| Carotid vascular grafts | ||||
| Lysosome | 16 | 12.5 | 6.3 | 8.8 × 10−7 |
| Cytoplasmic vesicle | 15 | 11.7 | 3.2 | 2.2 × 10−3 |
| Cell junction | 14 | 10.9 | 2.4 | 3.4 × 10−2 |
| Cytoskeleton | 20 | 15.6 | 2.2 | 9.6 × 10−3 |
| Cytoplasm | 66 | 51.6 | 1.8 | 8.8 × 10−7 |
| Gene Ontology Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| Laminin binding | 28 | 5 | 25.5 | 8.45 × 10−4 |
| Integrin binding | 159 | 15 | 13.5 | 7.24 × 10−10 |
| Actin binding | 451 | 35 | 11.1 | 4.67 × 10−22 |
| Collagen binding | 70 | 5 | 10.2 | 2.14 × 10−2 |
| Cytoskeletal protein binding | 1006 | 44 | 6.2 | 1.78 × 10−19 |
| Carotid vascular grafts | ||||
| Proton-transporting ATPase activity, rotational mechanism | 24 | 6 | 40.2 | 1.85 × 10−5 |
| Collagen binding | 70 | 6 | 13.8 | 1.57 × 10−3 |
| Protein folding chaperone | 68 | 5 | 11.8 | 1.31 × 10−2 |
| Unfolded protein binding | 124 | 7 | 9.1 | 3.30 × 10−3 |
| Chaperone binding | 107 | 6 | 9.0 | 1.21 × 10−2 |
| UniProtKB Keywords Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| Muscle protein | 13 | 8.6 | 24.2 | 2.8 × 10−12 |
| Integrin | 5 | 3.3 | 12.9 | 6.2 × 10−3 |
| Actin-binding | 24 | 15.9 | 9.9 | 3.7 × 10−15 |
| Oxidoreductase | 15 | 9.9 | 2.9 | 6.2 × 10−3 |
| Carotid vascular grafts | ||||
| Actin capping | 4 | 3.1 | 26.4 | 7.0 × 10−3 |
| Chaperone | 10 | 7.8 | 5.6 | 2.1 × 10−3 |
| Actin-binding | 11 | 8.6 | 4.8 | 2.1 × 10−3 |
| Gene Ontology Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| Actin filament depolymerization | 12 | 4 | 47.7 | 7.46 × 10−4 |
| Basement membrane organization | 30 | 5 | 23.8 | 8.01 × 10−4 |
| Stress fiber assembly | 18 | 3 | 23.8 | 3.23 × 10−2 |
| Myofibril assembly | 62 | 10 | 23.1 | 5.04 × 10−8 |
| Focal adhesion assembly | 28 | 4 | 20.4 | 7.90 × 10−3 |
| Extracellular matrix assembly | 35 | 4 | 16.3 | 1.54 × 10−2 |
| Endothelial cell development | 51 | 5 | 14.0 | 5.61 × 10−3 |
| Integrin-mediated signaling pathway | 101 | 9 | 12.7 | 2.35 × 10−5 |
| Muscle cell development | 160 | 14 | 12.5 | 1.79 × 10−8 |
| Cellular detoxification | 108 | 8 | 10.6 | 3.52 × 10−4 |
| Muscle contraction | 230 | 15 | 9.3 | 1.15 × 10−7 |
| Actin cytoskeleton organization | 549 | 30 | 7.8 | 2.53 × 10−14 |
| Cell-cell adhesion | 543 | 20 | 5.3 | 1.14 × 10−6 |
| Extracellular matrix organization | 280 | 10 | 5.1 | 4.99 × 10−3 |
| Cytoskeleton organization | 1240 | 39 | 4.5 | 5.38 × 10−12 |
| Carotid vascular grafts | ||||
| NADPH regeneration | 16 | 4 | 40.2 | 2.33 × 10−3 |
| Pentose metabolic process | 12 | 3 | 40.2 | 1.67 × 10−2 |
| Glucose 6-phosphate metabolic process | 23 | 5 | 35.0 | 4.58 × 10−4 |
| Tricarboxylic acid cycle | 31 | 4 | 20.8 | 1.26 × 10−2 |
| Collagen metabolic process | 61 | 7 | 18.5 | 2.94 × 10−4 |
| NADP metabolic process | 37 | 4 | 17.4 | 1.89 × 10−2 |
| Collagen catabolic process | 41 | 4 | 15.7 | 2.32 × 10−2 |
| Intracellular pH reduction | 50 | 4 | 12.9 | 3.88 × 10−2 |
| Chaperone-mediated protein folding | 71 | 5 | 11.3 | 1.79 × 10−2 |
| Response to unfolded protein | 123 | 6 | 7.8 | 2.16 × 10−2 |
| Protein folding | 221 | 9 | 6.5 | 4.17 × 10−3 |
| Generation of precursor metabolites and energy | 406 | 12 | 4.7 | 3.74 × 10−3 |
| Carbohydrate metabolic process | 475 | 11 | 3.7 | 2.76 × 10−2 |
| Carbohydrate derivative metabolic process | 1005 | 22 | 3.5 | 3.50 × 10−4 |
| Regulation of proteolysis | 743 | 14 | 3.0 | 3.04 × 10−2 |
| Reactome Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| Fibronectin matrix formation | 6 | 3 | 71.5 | 1.80 × 10−3 |
| Smooth muscle contraction | 39 | 16 | 58.7 | 7.34 × 10−19 |
| Cell–extracellular matrix interactions | 18 | 7 | 55.6 | 1.04 × 10−7 |
| Laminin interactions | 30 | 11 | 52.4 | 2.70 × 10−12 |
| Protein repair | 6 | 2 | 47.7 | 4.07 × 10−2 |
| Dermatan sulfate biosynthesis | 11 | 3 | 39.0 | 5.86 × 10−3 |
| Chondroitin sulfate biosynthesis | 19 | 3 | 22.6 | 1.79 × 10−2 |
| Non-integrin membrane-ECM interactions | 59 | 9 | 21.8 | 2.82 × 10−7 |
| ECM proteoglycans | 76 | 11 | 20.7 | 1.10 × 10−8 |
| Integrin signaling | 26 | 3 | 16.5 | 3.49 × 10−2 |
| Syndecan interactions | 27 | 3 | 15.9 | 3.76 × 10−2 |
| Elastic fibre formation | 45 | 5 | 15.9 | 1.72 × 10−3 |
| Molecules associated with elastic fibres | 38 | 4 | 15.0 | 9.95 × 10−3 |
| Integrin cell surface interactions | 84 | 8 | 13.6 | 3.95 × 10−5 |
| Cell junction organization | 91 | 8 | 12.6 | 5.87 × 10−5 |
| Carotid vascular grafts | ||||
| Pentose phosphate pathway | 15 | 4 | 42.9 | 6.58 × 10−4 |
| Insulin receptor recycling | 26 | 6 | 37.1 | 1.63 × 10−5 |
| ROS and RNS production in phagocytes | 36 | 6 | 26.8 | 4.90 × 10−5 |
| Citric acid cycle (TCA cycle) | 22 | 3 | 21.9 | 2.49 × 10−2 |
| Assembly of collagen fibrils and other multimeric structures | 60 | 6 | 16.1 | 5.17 × 10−4 |
| Collagen degradation | 64 | 6 | 15.1 | 6.50 × 10−4 |
| Collagen formation | 89 | 8 | 14.5 | 3.87 × 10−5 |
| Collagen biosynthesis and modifying enzymes | 67 | 5 | 12.0 | 6.95 × 10−3 |
| Trans-Golgi network vesicle budding | 72 | 5 | 11.2 | 8.98 × 10−3 |
| Degradation of the extracellular matrix | 140 | 8 | 9.2 | 5.71 × 10−4 |
| Eukaryotic translation elongation | 94 | 5 | 8.6 | 2.03 × 10−2 |
| Endoplasmic reticulum to Golgi anterograde transport | 155 | 6 | 6.2 | 2.54 × 10−2 |
| Asparagine N-linked glycosylation | 305 | 10 | 5.3 | 2.56 × 10−3 |
| Transport to the Golgi and subsequent modification | 186 | 6 | 5.2 | 4.94 × 10−2 |
| Membrane trafficking | 626 | 12 | 3.1 | 2.95 × 10−2 |
| Kyoto Encyclopedia of Genes and Genomes Term | Total Proteins | Differentially Expressed Proteins | Fold Enrichment | FDR-Corrected p Value |
|---|---|---|---|---|
| Native carotid arteries | ||||
| ECM–receptor interaction | 12 | 7.9 | 13.4 | 5.8 × 10−8 |
| Focal adhesion | 24 | 15.9 | 11.7 | 1.5 × 10−16 |
| Vascular smooth muscle contraction | 10 | 6.6 | 7.3 | 1.3 × 10−4 |
| Regulation of actin cytoskeleton | 16 | 10.6 | 7.2 | 9.6 × 10−8 |
| Carotid vascular grafts | ||||
| Lysosome | 14 | 10.9 | 9.3 | 4.3 × 10−7 |
| Carbon metabolism | 11 | 8.6 | 8.4 | 3.8 × 10−5 |
| Phagosome | 13 | 10.2 | 7.5 | 1.1 × 10−5 |
| Biosynthesis of amino acids | 6 | 4.7 | 7.0 | 3.0 × 10−2 |
| Metabolic pathways | 38 | 29.7 | 2.2 | 7.2 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonova, L.; Kutikhin, A.; Sevostianova, V.; Lobov, A.; Repkin, E.; Krivkina, E.; Velikanova, E.; Mironov, A.; Mukhamadiyarov, R.; Senokosova, E.; et al. Controlled and Synchronised Vascular Regeneration upon the Implantation of Iloprost- and Cationic Amphiphilic Drugs-Conjugated Tissue-Engineered Vascular Grafts into the Ovine Carotid Artery: A Proteomics-Empowered Study. Polymers 2022, 14, 5149. https://doi.org/10.3390/polym14235149
Antonova L, Kutikhin A, Sevostianova V, Lobov A, Repkin E, Krivkina E, Velikanova E, Mironov A, Mukhamadiyarov R, Senokosova E, et al. Controlled and Synchronised Vascular Regeneration upon the Implantation of Iloprost- and Cationic Amphiphilic Drugs-Conjugated Tissue-Engineered Vascular Grafts into the Ovine Carotid Artery: A Proteomics-Empowered Study. Polymers. 2022; 14(23):5149. https://doi.org/10.3390/polym14235149
Chicago/Turabian StyleAntonova, Larisa, Anton Kutikhin, Viktoriia Sevostianova, Arseniy Lobov, Egor Repkin, Evgenia Krivkina, Elena Velikanova, Andrey Mironov, Rinat Mukhamadiyarov, Evgenia Senokosova, and et al. 2022. "Controlled and Synchronised Vascular Regeneration upon the Implantation of Iloprost- and Cationic Amphiphilic Drugs-Conjugated Tissue-Engineered Vascular Grafts into the Ovine Carotid Artery: A Proteomics-Empowered Study" Polymers 14, no. 23: 5149. https://doi.org/10.3390/polym14235149