Xylans of Red and Green Algae: What Is Known about Their Structures and How They Are Synthesised?



Abstract
:1. Introduction
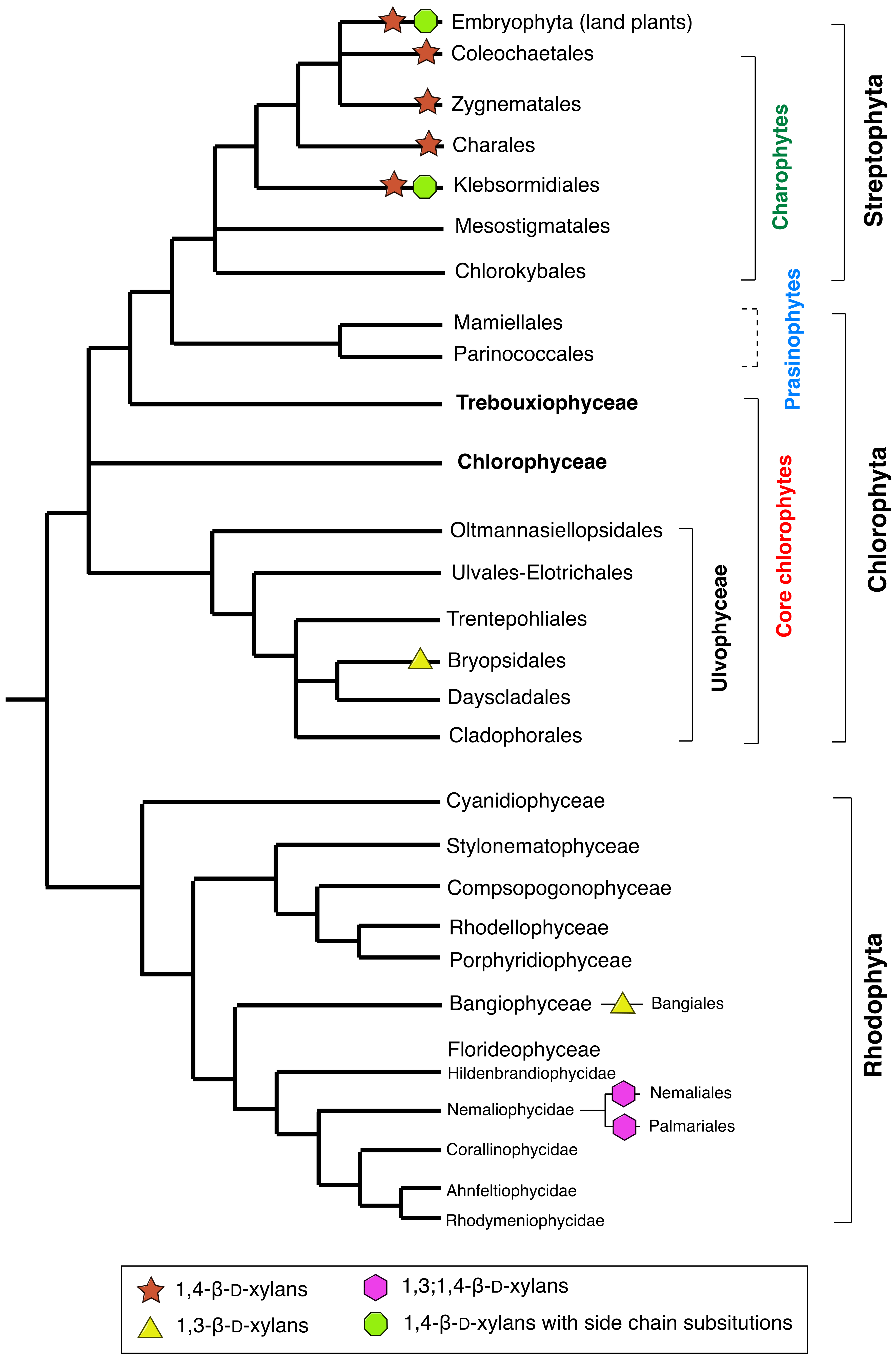
2. Xylans in Green and Red Algae
2.1. Xylans in Charophyte Green Algae
2.2. Xylans in Chlorophyte Green Algae
2.3. Xylans in Red Algae (Rhodophyta)
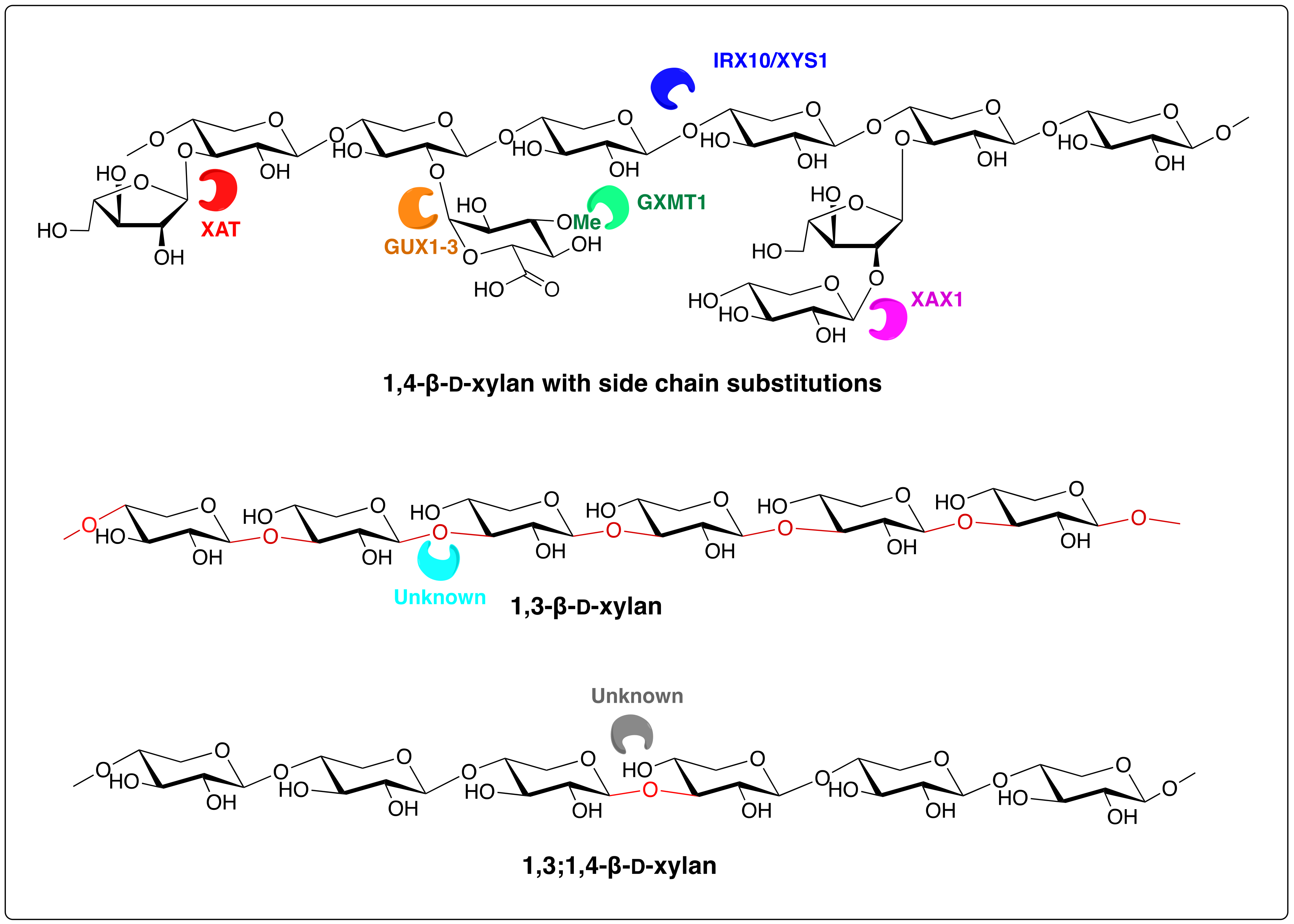
3. Biosynthesis of Algal Xylans
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ebringerová, A.; Heinze, T. Xylan and xylan derivatives—Biopolymers with valuable properties, 1. Naturally occurring xylans structures, isolation procedures and properties. Macromol. Rapid Commun. 2000, 21, 542–556. [Google Scholar] [CrossRef]
- Pauly, M.; Keegstra, K. Plant cell wall polymers as precursors for biofuels. Curr. Opin. Plant Biol. 2010, 13, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Ur-Rehman, S.; Mushtaq, Z.; Zahoor, T.; Jamil, A.; Murtaza, M.A. Xylitol: A review on bioproduction, application, health benefits, and related safety issues. Crit. Rev. Food Sci. Nutr. 2015, 55, 1514–1528. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.J.; Stone, B.A. Chemistry and Molecular Organization of Plant Cell Walls. In Biomass Recalcitrance: Deconstructing the Plant Cell Wall for Bioenergy; Himmel, M.E., Ed.; Wiley-Blackwell: Chichester, UK, 2009; pp. 61–93. [Google Scholar]
- Harris, P.J. Diversity in Plant Cell Walls. In Plant Diversity and Evolution: Genotypic and Phenotypic Variation in Higher Plants; Henry, R.J., Ed.; CAB International Publishing: Wallingford, UK, 2005; pp. 201–227. [Google Scholar]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Busse-Wicher, M.; Li, A.; Silveira, R.L.; Pereira, C.S.; Tryfona, T.; Gomes, T.C.; Skaf, M.S.; Dupree, P. Evolution of xylan substitution patterns in gymnosperms and angiosperms: Implications for xylan interaction with cellulose. Plant Physiol. 2016, 171, 2418–2431. [Google Scholar] [CrossRef] [PubMed]
- Peña, M.J.; Kulkarni, A.R.; Backe, J.; Boyd, M.; O’Neill, M.A.; York, W.S. Structural diversity of xylans in the cell walls of monocots. Planta 2016, 244, 589–606. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, K.C.B. The Non-Cellulosic Polysaccharides of Non-Endospermic Parts of Grasses. In New Approaches to Research on Cereal Carbohydrates; Hill, R.D., Munck, L., Eds.; Elsevier Science: Amsterdam, The Netherlands, 1985; pp. 231–240. [Google Scholar]
- Kulkarni, A.R.; Peña, M.J.; Avci, U.; Mazumder, K.; Urbanowicz, B.R.; Pattathil, S.; Yin, Y.; O’Neill, M.A.; Roberts, A.W.; Hahn, M.G.; et al. The ability of land plants to synthesize glucuronoxylans predates the evolution of tracheophytes. Glycobiology 2012, 22, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.; Clayden, S.; Reyes-Prieto, A. The Glaucophyta: The blue-green plants in a nutshell. Acta Soc. Bot. Pol. 2015, 84, 149–165. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Hardouin, K.; Potin, P.; Kloareg, B.; Herve, C. A review about brown algal cell walls and fucose-containing sulfated polysaccharides: Cell wall context, biomedical properties and key research challenges. Carbohydr. Polym. 2017, 175, 395–408. [Google Scholar] [CrossRef]
- Deniaud-Bouët, E.; Kervarec, N.; Michel, G.; Tonon, T.; Kloareg, B.; Hervé, C. Chemical and enzymatic fractionation of cell walls from Fucales: Insights into the structure of the extracellular matrix of brown algae. Ann. Bot. 2014, 114, 1203–1216. [Google Scholar] [CrossRef]
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; Clerck, O.D. Phylogeny and molecular evolution of the green algae. Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef]
- Sørensen, I.; Rose, J.K.C.; Doyle, J.J.; Domozych, D.S.; Willats, W.G.T. The charophycean green algae as model systems to study plant cell walls and other evolutionary adaptations that give rise to land plants. Plant Signal Behav. 2012, 7, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Sun, L.; Penny, D. The origin of land plants: A phylogenomic perspective. Evol. Bioinform. 2015, 11, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Xi, Z.; Goremykin, V.V.; Fong, R.; McLenachan, P.A.; Novis, P.M.; Davis, C.C.; Penny, D. Streptophyte algae and the origin of land plants revisited using heterogeneous models with three new algal chloroplast genomes. Mol. Biol. Evol. 2013, 31, 177–183. [Google Scholar] [CrossRef]
- Sørensen, I.; Pettolino, F.A.; Bacic, A.; Ralph, J.; Lu, F.; O’Neill, M.A.; Fei, Z.; Rose, J.K.; Domozych, D.S.; Willats, W.G. The charophycean green algae provide insights into the early origins of plant cell walls. Plant J. 2011, 68, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.K.; Busse-Wicher, M.; Poulsen, C.P.; Fangel, J.U.; Smith, P.J.; Yang, J.-Y.; Peña, M.-J.; Dinesen, M.H.; Martens, H.J.; Melkonian, M.; et al. Identification of an algal xylan synthase indicates that there is functional orthology between algal and plant cell wall biosynthesis. New Phytol. 2018, 218, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Domozych, D.S.; Sørensen, I.; Pettolino, F.A.; Bacic, A.; Willats, W.G.T. The cell wall polymers of the charophycean green alga Chara corallina: Immunobinding and biochemical screening. Int. J. Plant Sci. 2010, 171, 345–361. [Google Scholar] [CrossRef]
- Verbruggen, H.; Ashworth, M.; LoDuca, S.T.; Vlaeminck, C.; Cocquyt, E.; Sauvage, T.; Zechman, F.W.; Littler, D.S.; Littler, M.M.; Leliaert, F.; et al. A multi-locus time-calibrated phylogeny of the siphonous green algae. Mol. Phylogenet. Evol. 2009, 50, 642–653. [Google Scholar] [CrossRef]
- Frei, E.; Preston, R.D. Non-cellulosic structural polysaccharides in algal cell walls I. Xylan in siphoneous green algae. Proc. R. Soc. Lond. B Biol. Sci. 1964, 160, 293–313. [Google Scholar] [CrossRef]
- Frei, E.; Preston, R.D. Non-cellulosic structural polysaccharides in algal cell walls. III. Mannan in siphoneous green algae. Proc. R. Soc. Lond. B Biol. Sci. 1968, 169, 127–145. [Google Scholar] [CrossRef]
- Wutz, M.; Zetsche, K. Biochemistry and regulation of the heteromorphic life cycle of the green alga Derbesia-Halicystis. Planta 1976, 129, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Estevez, J.M.; Fernandez, P.V.; Kasulin, L.; Dupree, P.; Ciancia, M. Chemical and in situ characterization of macromolecular components of the cell walls from the green seaweed Codium fragile. Glycobiology 2009, 19, 212–228. [Google Scholar] [CrossRef] [PubMed]
- Ciancia, M.; Alberghina, J.; Arata, P.X.; Benavides, H.; Leliaert, F.; Verbruggen, H.; Estevez, J.M. Characterization of cell wall polysaccharides of the coencocytic green seaweed Bryopsis plumosa (Bryopsidaceae, Chlorophyta) from the Argentine coast. J. Phycol. 2012, 48, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Atkins, E.D.T.; Parker, K.D.; Preston, R.D. The helical structure of the β-1,3-linked xylan in some siphoneous green algae. Proc. R. Soc. Lond. B Biol. Sci. 1969, 173, 209–221. [Google Scholar] [CrossRef]
- Stone, B. Callose and Related Glucans. In eLS; John Wiley & Sons Ltd.: Chichester, UK, 2006. [Google Scholar]
- Huizing, H.J.; Rietma, H.; Sietsma, J.H. Cell wall constituents of several siphoneous green algae in relation to morphology and taxonomy. Br. Phycol. J. 1979, 14, 25–32. [Google Scholar] [CrossRef]
- Dunn, E.K.; Shoue, D.A.; Huang, X.; Kline, R.E.; MacKay, A.L.; Carpita, N.C.; Taylor, I.E.P.; Mandoli, D.F. Spectroscopic and biochemical analysis of regions of the cell wall of the unicellular ‘mannan weed’, Acetabularia acetabulum. Plant Cell Physiol. 2007, 48, 122–133. [Google Scholar] [CrossRef]
- Craigie, J.S. Cell Walls. In Biology of the Red Algae; Cole, K.M., Sheath, R.G., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 221–257. [Google Scholar]
- Adams, N.M.; Furneaux, R.H.; Miller, I.J.; Whitehouse, L.A. Xylan from Leptosarca simplex and carrageenans from Iridea, Cenacrum and Nemastoma species from the subantarctic islands of New Zealand. Bot. Mar. 1988, 31, 9–14. [Google Scholar] [CrossRef]
- Furneaux, R.H.; Miller, I.J. Isolation and 13C-NMR spectral study of the water soluble polysaccharides from four South African red algae. Bot. Mar. 1986, 29, 3–10. [Google Scholar] [CrossRef]
- Frei, E.; Preston, R.D. Non-cellulosic structural polysaccharides in algal cell walls—II. Association of xylan and mannan in Porphyra umbilicalis. Proc. R. Soc. Lond. B Biol. Sci. 1964, 160, 314–327. [Google Scholar] [CrossRef]
- Gretz, M.R.; Aronson, J.M.; Sommerfeld, M.R. Cellulose in the cell walls of the Bangiophyceae (Rhodophyta). Science 1980, 207, 779–781. [Google Scholar] [CrossRef]
- Gretz, M.R.; Aronson, J.M.; Sommerfeld, M.R. Taxonomic significance of cellulosic cell walls in the Bangiales (Rhodophyta). Phytochemistry 1984, 23, 2513–2514. [Google Scholar] [CrossRef]
- Yoon, H.S.; Nelson, W.; Lindtrom, S.C.; Boo, S.M.; Pueschel, C.; Qiu, H.; Bhattacharya, D. Rhodophyta. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 89–133. [Google Scholar]
- Turvey, J.R.; Williams, E.L. The structures of some xylans from red algae. Phytochemistry 1970, 9, 2383–2388. [Google Scholar] [CrossRef]
- Viana, A.G.; Noseda, M.D.; Gonçalves, A.G.; Duarte, M.E.R.; Yokoya, N.; Matulewicz, M.C.; Cerezo, A.S. β-d-(1→4), β-d-(1→3) ‘mixed linkage’ xylans from red seaweeds of the order Nemaliales and Palmariales. Carbohydr. Res. 2011, 346, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Usov, A.I. Polysaccharides of the red algae. Adv. Carbohydr. Chem. Biochem. 2011, 65, 115–217. [Google Scholar] [CrossRef] [PubMed]
- Morgan, K.C.; Wright, J.L.C.; Simpson, F.J. Review of chemical constituents of the red alga Palmaria palmata (dulse). Econ. Bot. 1980, 34, 27–50. [Google Scholar] [CrossRef]
- Deniaud, E.; Quemener, B.; Fleurence, J.; Lahaye, M. Structural studies of the mix-linked β-(1→3)/β-(1→4)-d-xylans from the cell wall of Palmaria palmata (Rhodophyta). Int. J. Biol. Macromol. 2003, 33, 9–18. [Google Scholar] [CrossRef]
- Nerinckx, W.; Broberg, A.; Duus, J.Ø.; Ntarima, P.; Parolis, L.A.S.; Parolis, H.; Claeyssens, M. Hydrolysis of Nothogenia erinacea xylan by xylanases from families 10 and 11. Carbohydr. Res. 2004, 339, 1047–1060. [Google Scholar] [CrossRef]
- Painter, T.J. Algal Polysaccharides. In The Polysaccharides; Aspinall, G.O., Ed.; Academic Press: New York, NY, USA, 1983; pp. 195–285. [Google Scholar]
- Lahaye, M.; Rondeau-Mouro, C.; Deniaud, E.; Buléon, A. Solid-state 13C NMR spectroscopy studies of xylans in the cell wall of Palmaria palmata (L. Kuntze, Rhodophyta). Carbohydr. Res. 2003, 338, 1559–1569. [Google Scholar] [CrossRef]
- Mandal, P.; Pujol, C.A.; Damonte, E.B.; Ghosh, T.; Ray, B. Xylans from Scinaia hatei: Structural features, sulfation and anti-HSV activity. Int. J. Biol. Macromol. 2010, 46, 173–178. [Google Scholar] [CrossRef]
- Rennie, E.A.; Scheller, H.V. Xylan biosynthesis. Curr. Opin. Biotechnol. 2014, 26, 100–107. [Google Scholar] [CrossRef]
- Smith, P.J.; Wang, H.T.; York, W.S.; Peña, M.J.; Urbanowicz, B.R. Designer biomass for next-generation biorefineries: Leveraging recent insights into xylan structure and biosynthesis. Biotechnol. Biofuels 2017, 10, 286. [Google Scholar] [CrossRef] [PubMed]
- Jensen, J.K.; Johnson, N.R.; Wilkerson, C.G. Arabidopsis thaliana IRX10 and two related proteins from psyllium and Physcomitrella patens are xylan xylosyltransferases. Plant J. 2014, 80, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Urbanowicz, B.R.; Peña, M.J.; Moniz, H.A.; Moremen, K.W.; York, W.S. Two Arabidopsis proteins synthesize acetylated xylan in vitro. Plant J. 2014, 80, 197–206. [Google Scholar] [CrossRef]
- Ren, Y.; Hansen, S.F.; Ebert, B.; Lau, J.; Scheller, H.V. Site-directed mutagenesis of IRX9, IRX9L and IRX14 proteins involved in xylan biosynthesis: Glycosyltransferase activity is not required for IRX9 function in Arabidopsis. PLoS ONE 2014, 9, e105014. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, Y.S.Y.; Harris, P.J. Xylans of Red and Green Algae: What Is Known about Their Structures and How They Are Synthesised? Polymers 2019, 11, 354. https://doi.org/10.3390/polym11020354
Hsieh YSY, Harris PJ. Xylans of Red and Green Algae: What Is Known about Their Structures and How They Are Synthesised? Polymers. 2019; 11(2):354. https://doi.org/10.3390/polym11020354
Chicago/Turabian StyleHsieh, Yves S.Y., and Philip J. Harris. 2019. "Xylans of Red and Green Algae: What Is Known about Their Structures and How They Are Synthesised?" Polymers 11, no. 2: 354. https://doi.org/10.3390/polym11020354