Protein–Polyelectrolyte Interaction: Thermodynamic Analysis Based on the Titration Method †


Abstract
:1. Introduction
2. Understanding of Protein–PE Binding Mechanisms
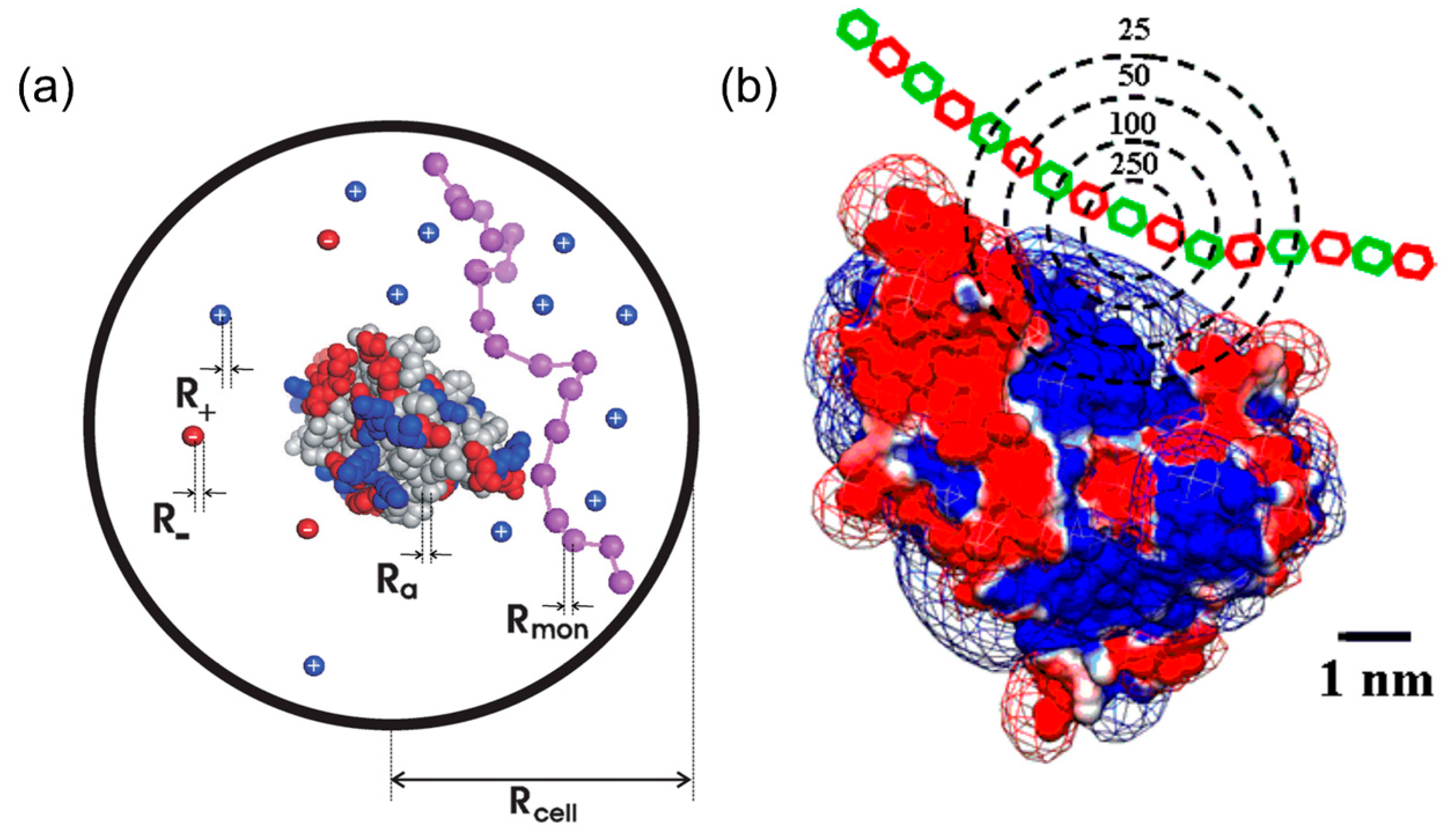
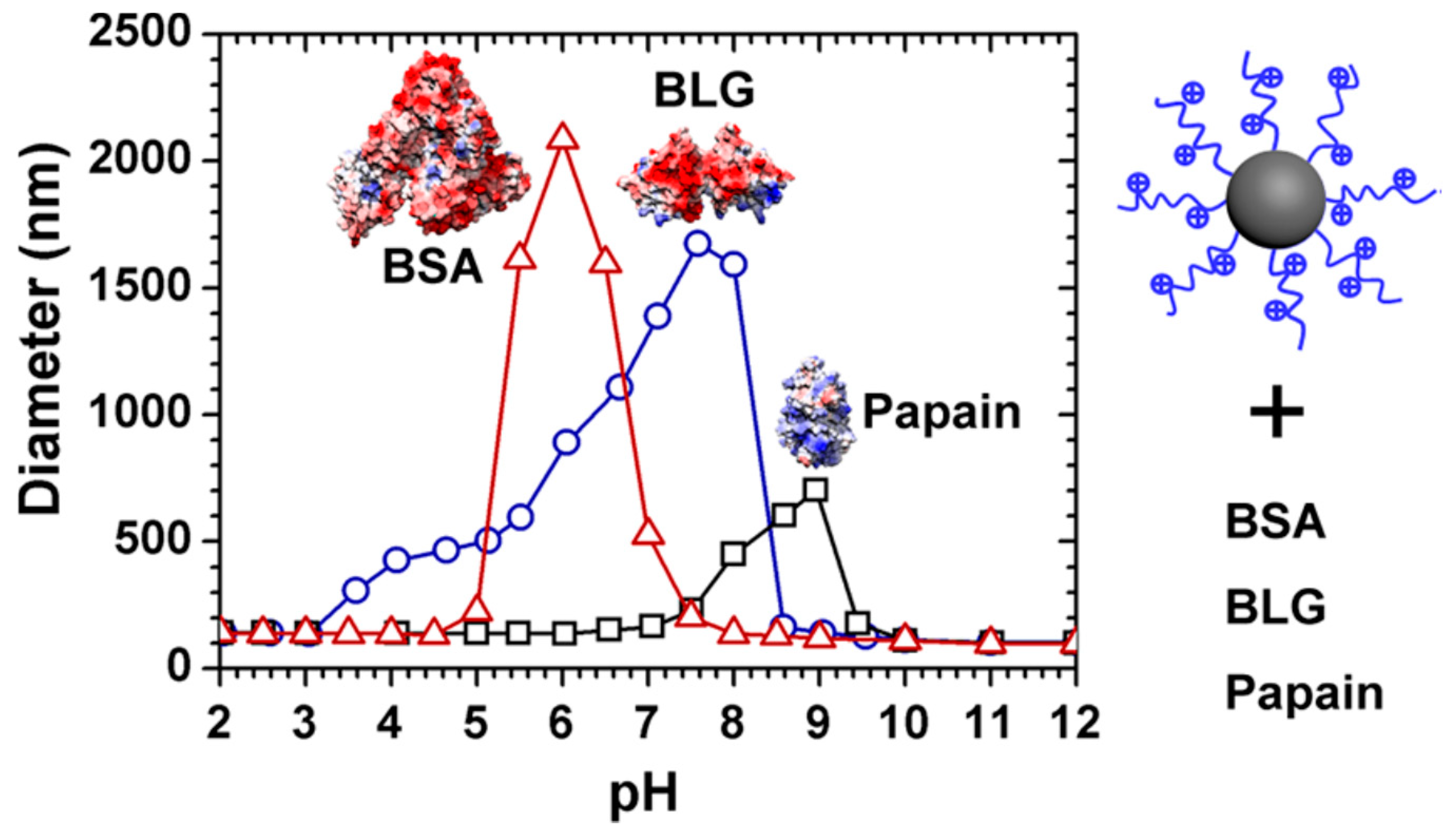
2.1. Charge Regulation and Charge Anisotropy
2.2. Titration Techniques for Protein–PE Binding Studies
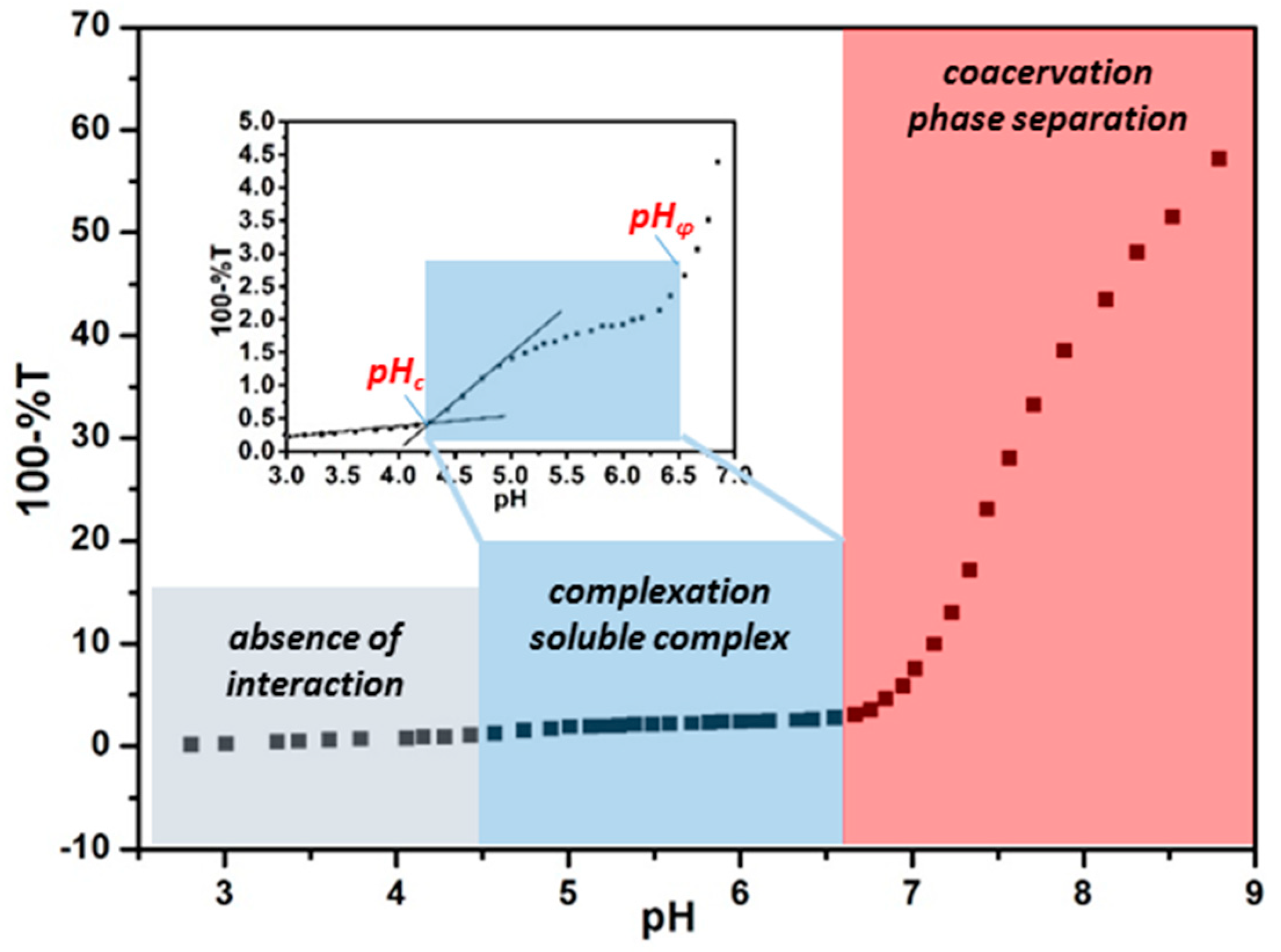
2.2.1. Turbidimetric Titration
2.2.2. Isothermal Titration Calorimetry (ITC)
3. Thermodynamic Studies of the Protein–PE Interaction
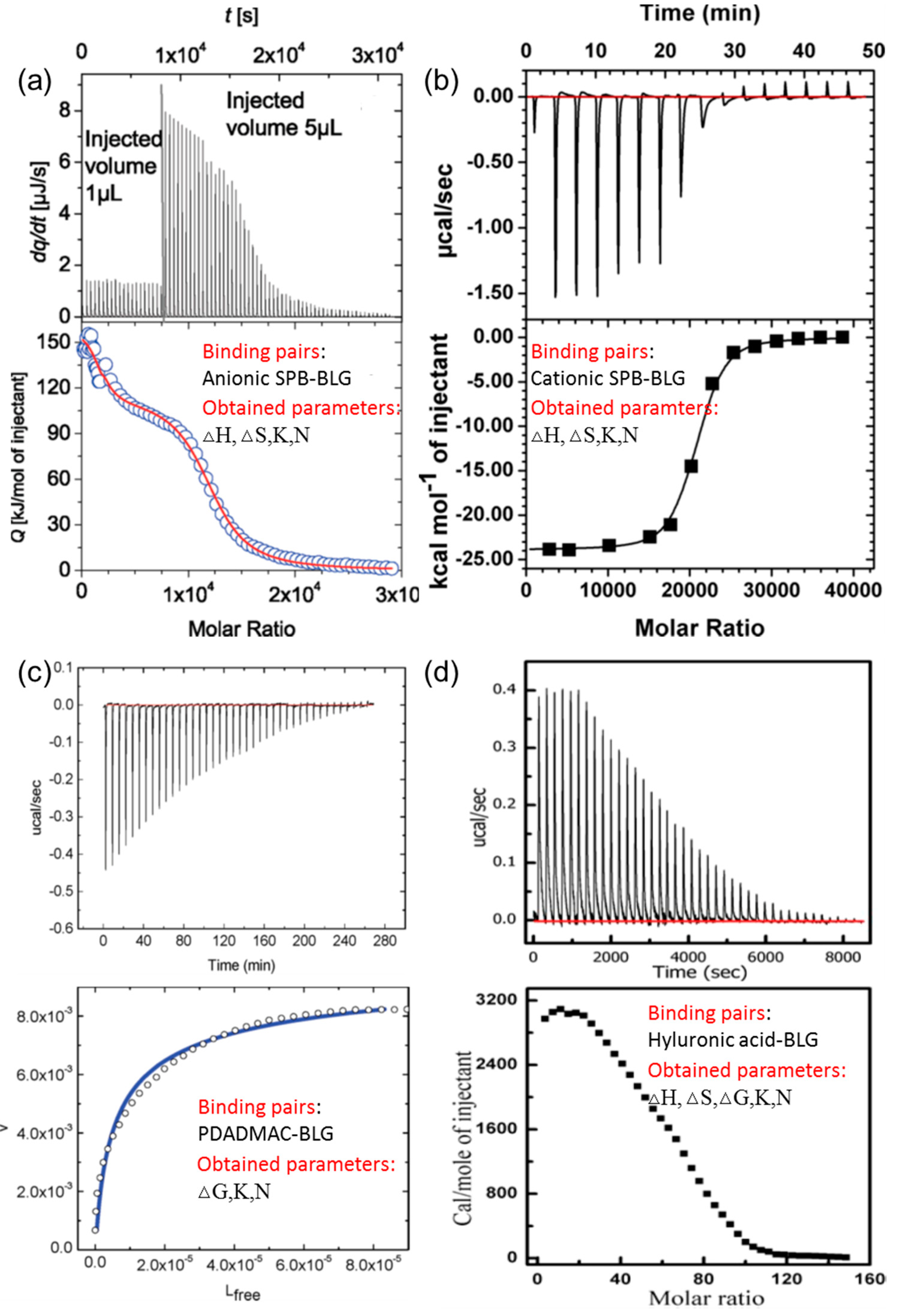
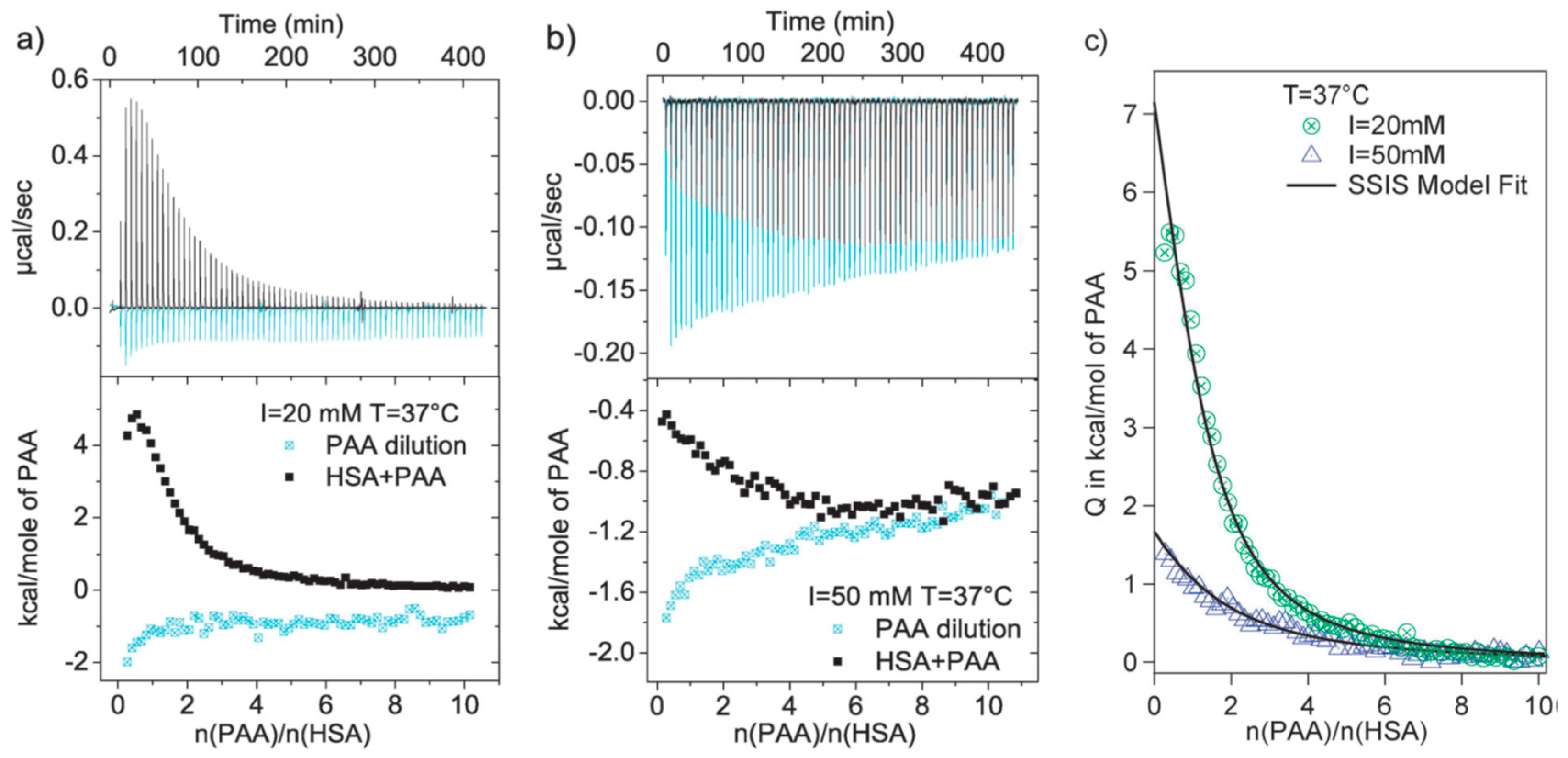
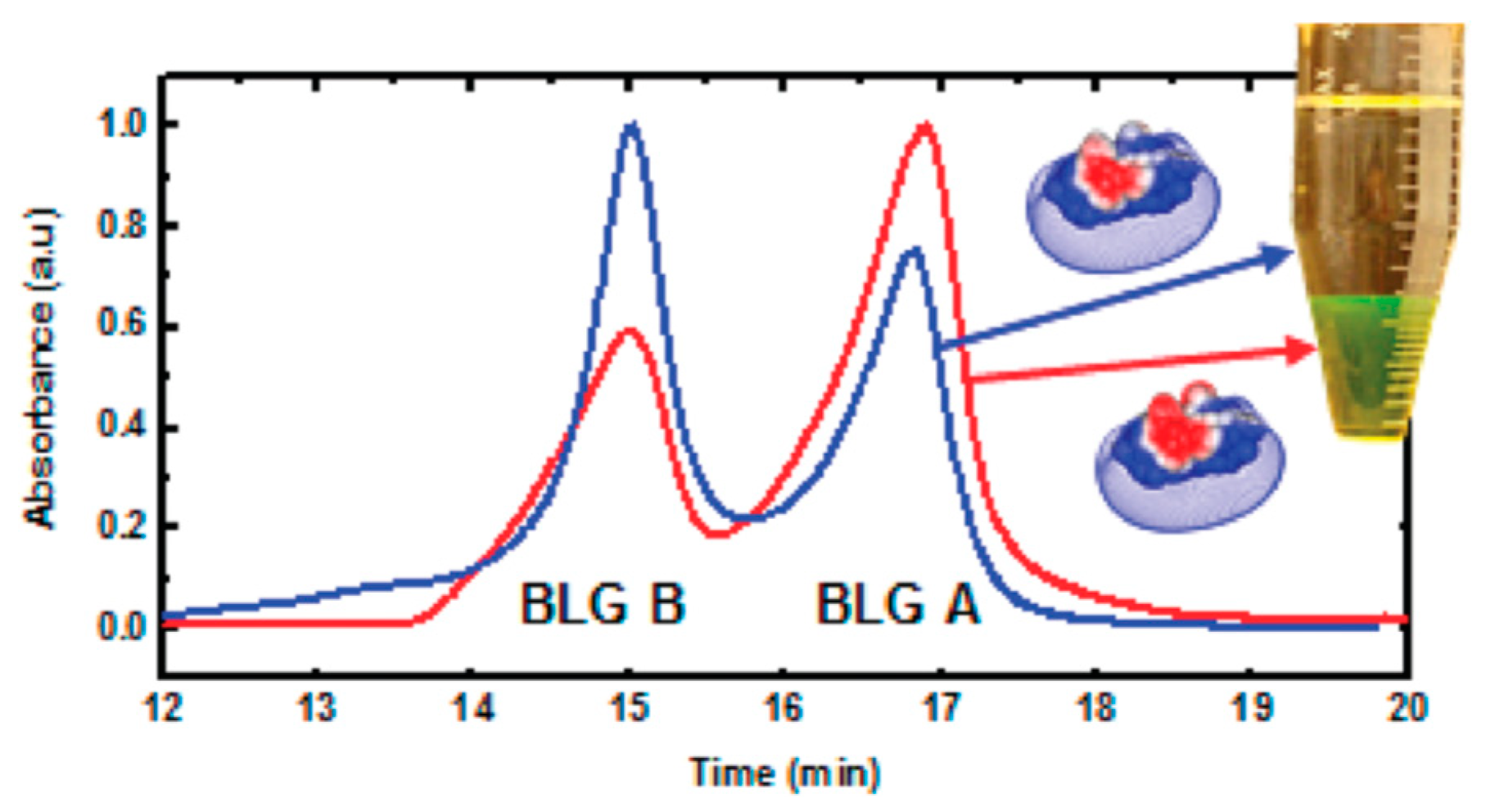
3.1. Linear Polyelectrolytes
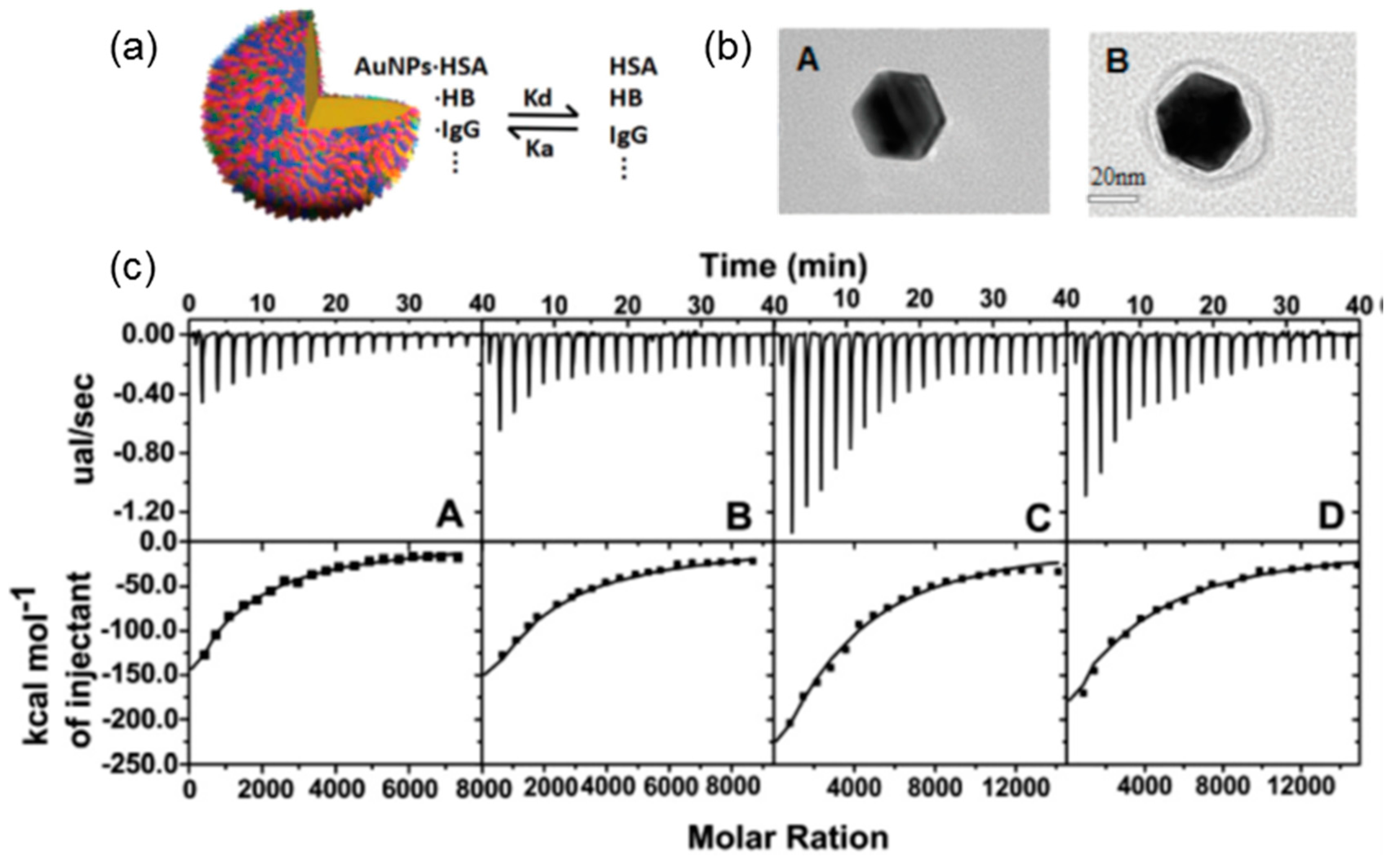
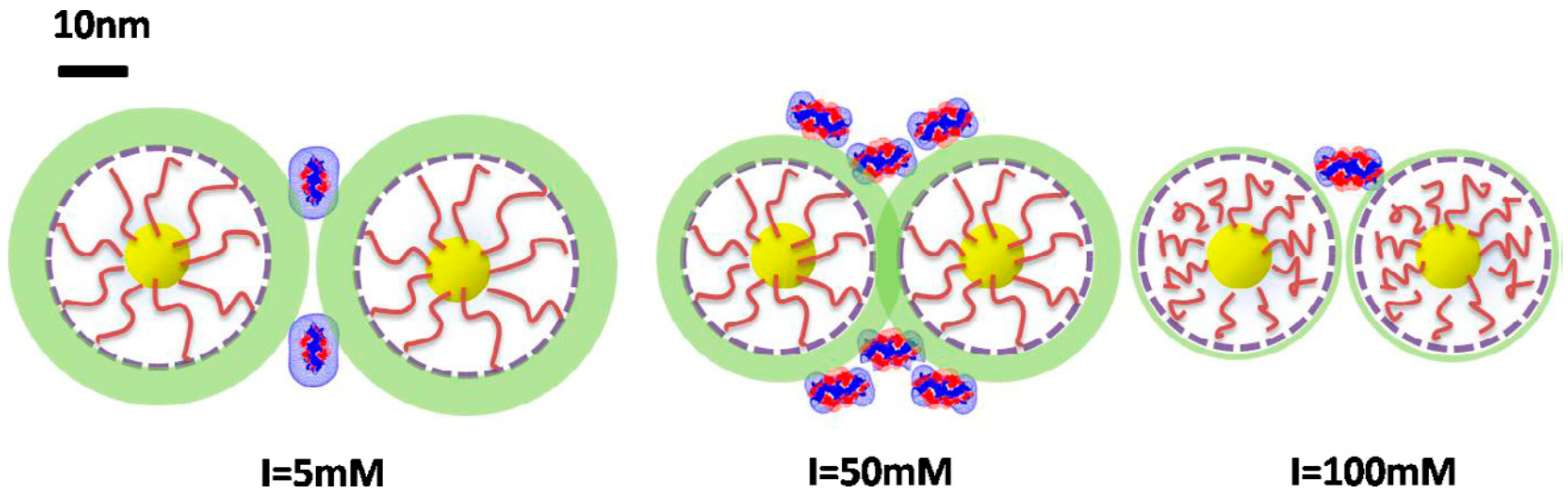
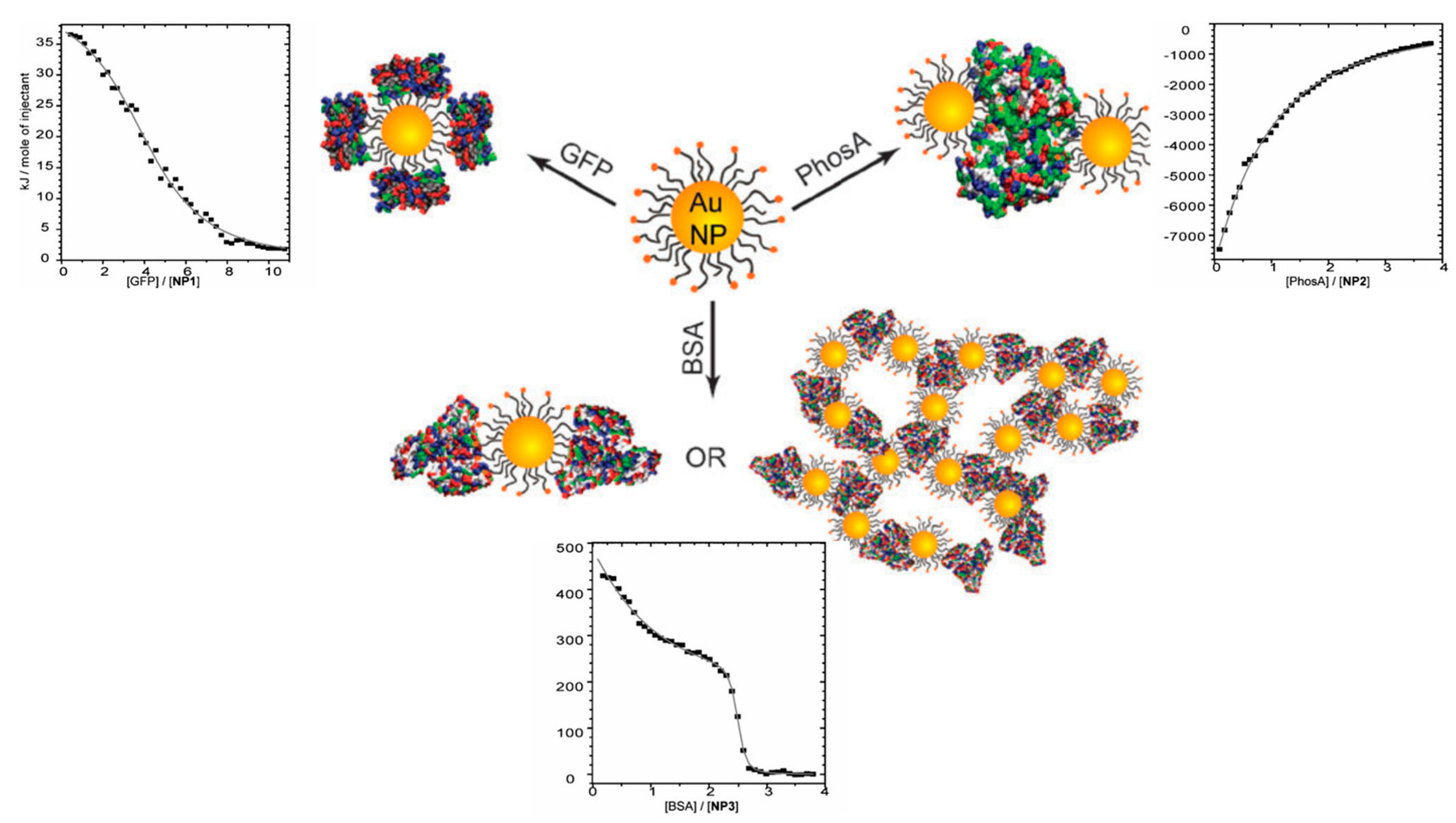
3.2. Polyelectrolytes Modified Nanoparticles
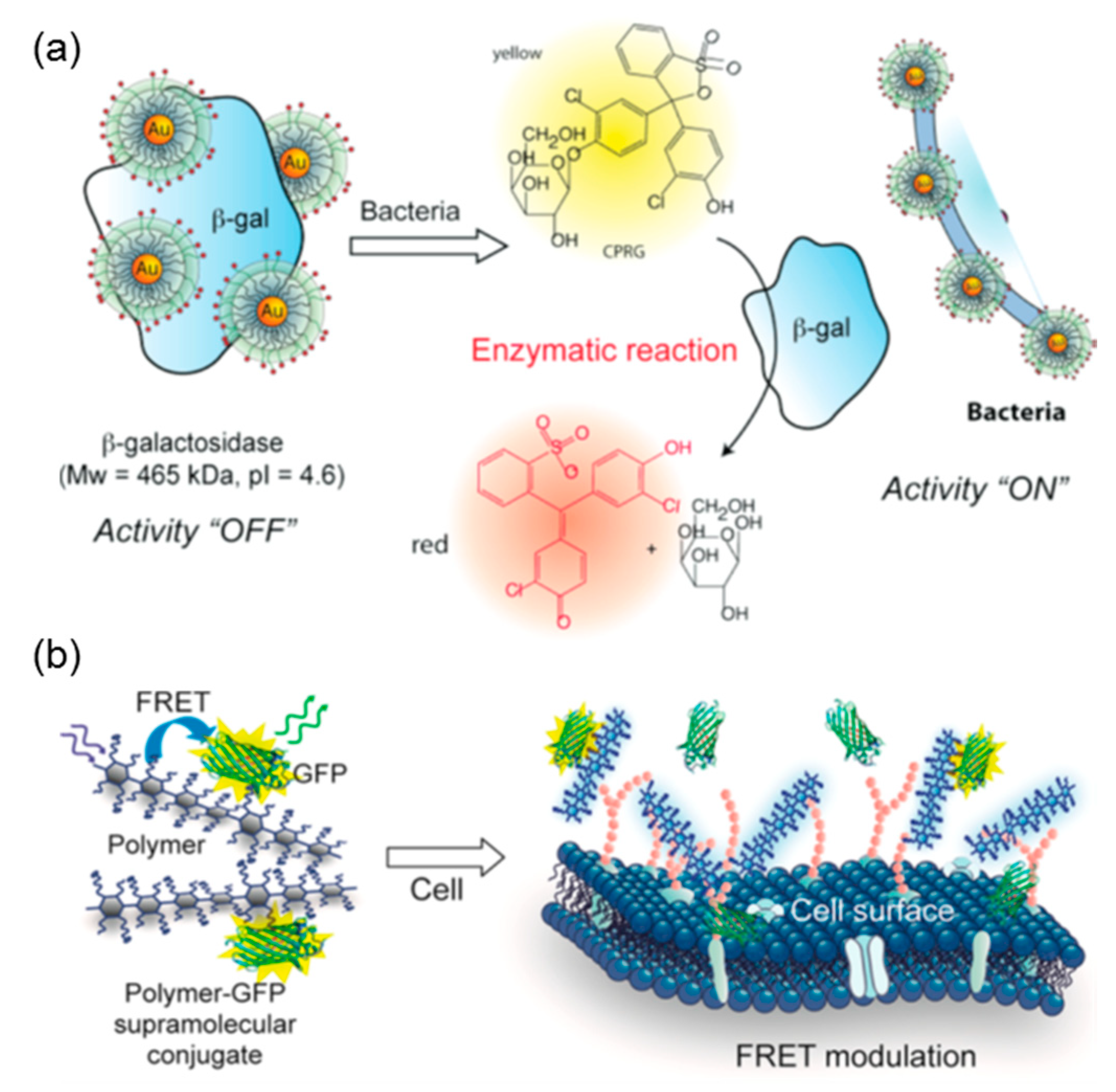
4. Thermodynamic Studies Guiding for Protein–PE Applications
4.1. Protein Immobilization
4.2. Protein Purification
5. Conclusions and Outlook
Funding
Conflicts of Interest
References
- Cousin, F.; Gummel, J.; Ung, D.; Boué, F. Polyelectrolyte−Protein Complexes: Structure and Conformation of Each Specie Revealed by SANS. Langmuir 2005, 21, 9675–9688. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.P.R.; Herland, A.; Hammarström, P.; Inganäs, O. Conjugated Polyelectrolytes: Conformation-Sensitive Optical Probes for Detection of Amyloid Fibril Formation. Biochemistry-us 2005, 44, 3718–3724. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, F.; Linse, P.; Malmsten, M. Monte Carlo Simulations of Polyelectrolyte−Protein Complexation. J. Phys. Chem. B 2001, 105, 9040–9049. [Google Scholar] [CrossRef]
- Han, J.; Bender, M.; Hahn, S.; Seehafer, K.; Bunz, U.H.F. Polyelectrolyte Complexes Formed from Conjugated Polymers: Array-Based Sensing of Organic Acids. Chem.-Eur. J. 2016, 22, 3230–3233. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Du, D.; Shao, Y.; Li, Z.; Wang, J.; Engelhard, M.H.; Li, J.; Lin, Y. Self assembly of acetylcholinesterase on a gold nanoparticles-graphene nanosheet hybrid for organophosphate pesticide detection using polyelectrolyte as a linker. J. Mater. Chem. 2011, 21, 5319–5325. [Google Scholar] [CrossRef]
- Islam, M.S.; Choi, H.N.; Choi, W.S.; Lee, H.-J. Polyelectrolyte-mediated hierarchical mesoporous calcium silicates: a platform for drug delivery carrier with ultrahigh loading capacity and controlled release behavior. J. Mater. Chem. B 2015, 3, 1001–1009. [Google Scholar] [CrossRef]
- Alvarez-Lorenzo, C.; Blanco-Fernandez, B.; Puga, A.M.; Concheiro, A. Crosslinked ionic polysaccharides for stimuli-sensitive drug delivery. Adv. Drug Deliv. Rev. 2013, 65, 1148–1171. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lin, Y.; Jia, L. Preparation of anionic polyelectrolyte modified magnetic nanoparticles for rapid and efficient separation of lysozyme from egg white. J. Chromatogr. A 2015, 1388, 43–51. [Google Scholar] [CrossRef]
- Yasar Mahlicli, F.; Alsoy Altinkaya, S.; Yurekli, Y. Preparation and characterization of polyacrylonitrile membranes modified with polyelectrolyte deposition for separating similar sized proteins. J. Membr. Sci. 2012, 415–416, 383–390. [Google Scholar] [CrossRef]
- Araujo, J.V.; Davidenko, N.; Danner, M.; Cameron, R.E.; Best, S.M. Novel porous scaffolds of pH responsive chitosan/carrageenan-based polyelectrolyte complexes for tissue engineering. J. Biomed. Mater. Res. Part A 2014, 102, 4415–4426. [Google Scholar] [CrossRef]
- Inturi, S.; Wang, G.; Chen, F.; Banda, N.K.; Holers, V.M.; Wu, L.; Moghimi, S.M.; Simberg, D. Modulatory Role of Surface Coating of Superparamagnetic Iron Oxide Nanoworms in Complement Opsonization and Leukocyte Uptake. ACS Nano 2015, 9, 10758–10768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koegler, P.; Clayton, A.; Thissen, H.; Santos, G.N.C.; Kingshott, P. The influence of nanostructured materials on biointerfacial interactions. Adv. Drug Deliv. Rev. 2012, 64, 1820–1839. [Google Scholar] [CrossRef]
- Smith, M.H.; Lyon, L.A. Tunable Encapsulation of Proteins within Charged Microgels. Macromolecules 2011, 44, 8154–8160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, L.; Henzler, K.; Lu, Y.; Wang, J.; Han, H.; Tian, Y.; Wang, Y.; Zhou, Z.; Lotze, G.; et al. Protein Immobilization onto Cationic Spherical Polyelectrolyte Brushes Studied by Small Angle X-ray Scattering. Biomacromolecules 2017, 18, 1574–1581. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, M.; Marradi, M.; Imberty, A.; Penadés, S.; Pérez, S. Multivalent Gold Glycoclusters: High Affinity Molecular Recognition by Bacterial Lectin PA-IL. Chem.-Eur. J. 2012, 18, 4264–4273. [Google Scholar] [CrossRef]
- Shaw, B.F.; Schneider, G.F.; Arthanari, H.; Narovlyansky, M.; Moustakas, D.; Durazo, A.; Wagner, G.; Whitesides, G.M. Complexes of Native Ubiquitin and Dodecyl Sulfate Illustrate the Nature of Hydrophobic and Electrostatic Interactions in the Binding of Proteins and Surfactants. J. Am. Chem. Soc. 2011, 133, 17681–17695. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Yoo, H.Y.; Huang, J.; Lee, Y.; Park, S.; Park, Y.; Jin, S.; Jung, Y.M.; Zeng, H.; Hwang, D.S.; et al. Salt Triggers the Simple Coacervation of an Underwater Adhesive When Cations Meet Aromatic pi Electrons in Seawater. ACS Nano 2017, 11, 6764–6772. [Google Scholar] [CrossRef] [PubMed]
- Braia, M.; Loureiro, D.; Tubio, G.; Lienqueo, M.E.; Romanini, D. Interaction between trypsin and alginate: An ITC and DLS approach to the formation of insoluble complexes. Colloids Surf. B 2017, 155, 507–511. [Google Scholar] [CrossRef]
- Xu, X.; Ran, Q.; Dey, P.; Nikam, R.; Haag, R.; Ballauff, M.; Dzubiella, J. Counterion-Release Entropy Governs the Inhibition of Serum Proteins by Polyelectrolyte Drugs. Biomacromolecules 2018, 19, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Winzen, S.; Schoettler, S.; Baier, G.; Rosenauer, C.; Mailaender, V.; Landfester, K.; Mohr, K. Complementary analysis of the hard and soft protein corona: sample preparation critically effects corona composition. Nanoscale 2015, 7, 2992–3001. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, F.L.B.; Jönsson, B. Polyelectrolyte–protein complexation driven by charge regulation. Soft Matter 2009, 5, 2862. [Google Scholar] [CrossRef]
- De Vos, W.M.; Leermakers, F.A.M.; de Keizer, A.; Cohen Stuart, M.A.; Kleijn, J.M. Field Theoretical Analysis of Driving Forces for the Uptake of Proteins by Like-Charged Polyelectrolyte Brushes: Effects of Charge Regulation and Patchiness. Langmuir 2010, 26, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.; Jönsson, B. Charge regulation in biomolecular solution. Q. Rev. Biophys. 2013, 46, 265–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayitmazer, A.B.; Quinn, B.; Kimura, K.; Ryan, G.L.; Tate, A.J.; Pink, D.A.; Dubin, P.L. Protein Specificity of Charged Sequences in Polyanions and Heparins. Biomacromolecules 2010, 11, 3325–3331. [Google Scholar] [CrossRef] [PubMed]
- Seyrek, E.; Dubin, P.L.; Tribet, C.; Gamble, E.A. Ionic strength dependence of protein-polyelectrolyte interactions. Biomacromolecules 2003, 4, 273–282. [Google Scholar] [PubMed]
- Silva, R.A.; Urzúa, M.D.; Petri, D.F.S.; Dubin, P.L. Protein Adsorption onto Polyelectrolyte Layers: Effects of Protein Hydrophobicity and Charge Anisotropy. Langmuir 2010, 26, 14032–14038. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Seeman, D.; Yan, Y.; Sun, L.; Post, J.; Dubin, P.L. Effect of Heparin on Protein Aggregation: Inhibition versus Promotion. Biomacromolecules 2012, 13, 1642–1651. [Google Scholar] [CrossRef] [Green Version]
- Henzler, K.; Haupt, B.; Lauterbach, K.; Wittemann, A.; Borisov, O.; Ballauff, M. Adsorption of β-Lactoglobulin on Spherical Polyelectrolyte Brushes: Direct Proof of Counterion Release by Isothermal Titration Calorimetry. J. Am. Chem. Soc. 2010, 132, 3159–3163. [Google Scholar] [CrossRef]
- Cooper, C.L.; Dubin, P.L.; Kayitmazer, A.B.; Turksen, S. Polyelectrolyte–protein complexes. Curr. Opin. Colloid Interface Sci. 2005, 10, 52–78. [Google Scholar]
- Du, X.; Dubin, P.L.; Hoagland, D.A.; Sun, L. Protein-Selective Coacervation with Hyaluronic Acid. Biomacromolecules 2014, 15, 726–734. [Google Scholar] [CrossRef]
- Stradner, A.; Sedgwick, H.; Cardinaux, F.; Poon, W.C.K.; Egelhaaf, S.U.; Schurtenberger, P. Equilibrium cluster formation in concentrated protein solutions and colloids. Nature 2004, 432, 492. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Mazzawi, M.; Chen, K.; Sun, L.; Dubin, P.L. Protein purification by polyelectrolyte coacervation: influence of protein charge anisotropy on selectivity. Biomacromolecules 2011, 12, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Antonov, M.; Mazzawi, M.; Dubin, P.L. Entering and Exiting the Protein−Polyelectrolyte Coacervate Phase via Nonmonotonic Salt Dependence of Critical Conditions. Biomacromolecules 2010, 11, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Xu, Y.; Han, H.; Liu, M.; Chen, K.; Wang, S.; Wang, J.; Xu, J.; Li, L.; Guo, X. beta-Lactoglobulin (BLG) binding to highly charged cationic polymer-grafted magnetic nanoparticles: effect of ionic strength. J. Colloid Interface Sci. 2015, 460, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Rana, S.; Moyano, D.F.; Xu, Y.; Guo, X.; Rotello, V.M. Optimizing the selective recognition of protein isoforms through tuning of nanoparticle hydrophobicity. Nanoscale 2014, 6, 6492–6495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Xu, Y.; Rana, S.; Miranda, O.R.; Dubin, P.L.; Rotello, V.M.; Sun, L.; Guo, X. Electrostatic selectivity in protein-nanoparticle interactions. Biomacromolecules 2011, 12, 2552–2561. [Google Scholar] [CrossRef] [PubMed]
- Hartvig, R.A.; van de Weert, M.; Østergaard, J.; Jorgensen, L.; Jensen, H. Protein Adsorption at Charged Surfaces: The Role of Electrostatic Interactions and Interfacial Charge Regulation. Langmuir 2011, 27, 2634–2643. [Google Scholar] [CrossRef] [PubMed]
- Barroso da Silva, F.L.; Boström, M.; Persson, C. Effect of Charge Regulation and Ion–Dipole Interactions on the Selectivity of Protein–Nanoparticle Binding. Langmuir 2014, 30, 4078–4083. [Google Scholar] [CrossRef] [PubMed]
- Kayitmazer, A.B.; Seeman, D.; Minsky, B.B.; Dubin, P.L.; Xu, Y. Protein–polyelectrolyte interactions. Soft Matter 2013, 9, 2553. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, S.; Xu, Y.; Zhao, X.; Guo, X. Ionic Strength-Responsive Binding between Nanoparticles and Proteins. Langmuir 2018, 34, 8264–8273. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Hallberg, R.; Dubin, P.L. Roles of Electrostatic Interaction and Polymer Structure in the Binding of β-Lactoglobulin to Anionic Polyelectrolytes: Measurement of Binding Constants by Frontal Analysis Continuous Capillary Electrophoresis. Langmuir 2000, 16, 9738–9743. [Google Scholar] [CrossRef]
- Kayitmazer, A.B.; Bohidar, H.B.; Mattison, K.W.; Bose, A.; Sarkar, J.; Hashidzume, A.; Russo, P.S.; Jaeger, W.; Dubin, P.L. Mesophase separation and probe dynamics in protein–polyelectrolyte coacervates. Soft Matter 2007, 3, 1064–1076. [Google Scholar] [CrossRef]
- Xu, Y.; Engel, Y.; Yan, Y.; Chen, K.; Moyano, D.F.; Dubin, P.L.; Rotello, V.M. Enhanced Electrostatic Discrimination of Proteins on Nanoparticle-Coated Surfaces. J. Mater. Chem. B 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Han, Y.; Qiao, R.; Zeng, J.; Jia, Q.; Wang, Y.; Gao, M. Investigations on the Interactions between Plasma Proteins and Magnetic Iron Oxide Nanoparticles with Different Surface Modifications. J. Phys. Chem. C 2010, 114, 21270–21276. [Google Scholar] [CrossRef]
- De, M.; You, C.-C.; Srivastava, S.; Rotello, V.M. Biomimetic Interactions of Proteins with Functionalized Nanoparticles: A Thermodynamic Study. J. Am. Chem. Soc. 2007, 129, 10747–10753. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Carney, R.P.; Ikuma, K.; Stellacci, F.; Lau, B.L.T. Effects of Surface Compositional and Structural Heterogeneity on Nanoparticle–Protein Interactions: Different Protein Configurations. ACS Nano 2014, 8, 5402–5412. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, K.; Li, L.; Guo, X. Binding between proteins and cationic spherical polyelectrolyte brushes: effect of pH, ionic strength, and stoichiometry. Biomacromolecules 2013, 14, 818–827. [Google Scholar] [CrossRef]
- Huang, R.; Lau, B.L. Biomolecule-nanoparticle interactions: Elucidation of the thermodynamics by isothermal titration calorimetry. Biochim. Biophys. Acta 2016, 1860, 945–956. [Google Scholar] [CrossRef]
- De, M.; Miranda, O.R.; Rana, S.; Rotello, V.M. Size and geometry dependent protein-nanoparticle self-assembly. Chem. Commun. 2009, 2157–2159. [Google Scholar] [CrossRef]
- Srinivasan, K.; Parimal, S.; Lopez, M.M.; McCallum, S.A.; Cramer, S.M. Investigation into the Molecular and Thermodynamic Basis of Protein Interactions in Multimodal Chromatography Using Functionalized Nanoparticles. Langmuir 2014, 30, 13205–13216. [Google Scholar] [CrossRef]
- Marangoni, V.S.; Paino, I.M.; Zucolotto, V. Synthesis and characterization of jacalin-gold nanoparticles conjugates as specific markers for cancer cells. Colloids Surf. B 2013, 112, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, S.; Han, H.; Chen, K.; Qin, L.; Xu, J.; Wang, J.; Li, L.; Guo, X. Enhancement of Enzymatic Activity by Magnetic Spherical Polyelectrolyte Brushes: A Potential Recycling Strategy for Enzymes. Langmuir 2014, 30, 11156–11164. [Google Scholar] [CrossRef] [PubMed]
- Pathak, J.; Rawat, K.; Aswal, V.K.; Bohidar, H.B. Interactions in globular proteins with polyampholyte: coacervation route for protein separation. RSC Adv. 2015, 5, 13579–13589. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zhang, F.; Yu, S. Probing the binding affinity of plasma proteins adsorbed on Au nanoparticles. Nanoscale 2017, 9, 4787–4792. [Google Scholar] [CrossRef] [PubMed]
- Aiken, G.R.; Hsu-Kim, H.; Ryan, J.N. Influence of Dissolved Organic Matter on the Environmental Fate of Metals, Nanoparticles, and Colloids. Environ. Sci. Technol. 2011, 45, 3196–3201. [Google Scholar] [CrossRef] [PubMed]
- Gagner, J.E.; Shrivastava, S.; Qian, X.; Dordick, J.S.; Siegel, R.W. Engineering Nanomaterials for Biomedical Applications Requires Understanding the Nano-Bio Interface: A Perspective. J. Phys. Chem. Lett. 2012, 3, 3149–3158. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wang, G.; Griffin, J.I.; Brenneman, B.; Banda, N.K.; Holers, V.M.; Backos, D.S.; Wu, L.; Moghimi, S.M.; Simberg, D. Complement proteins bind to nanoparticle protein corona and undergo dynamic exchange in vivo. Nat. Nanotechnol. 2016, 12, 387. [Google Scholar] [CrossRef]
- Huang, R.; Carney, R.P.; Stellacci, F.; Lau, B.L.T. Protein–nanoparticle interactions: the effects of surface compositional and structural heterogeneity are scale dependent. Nanoscale 2013, 5, 6928–6935. [Google Scholar] [CrossRef]
- Yu, S.; Xu, X.; Yigit, C.; van der Giet, M.; Zidek, W.; Jankowski, J.; Dzubiella, J.; Ballauff, M. Interaction of human serum albumin with short polyelectrolytes: a study by calorimetry and computer simulations. Soft Matter 2015, 11, 4630–4639. [Google Scholar] [CrossRef] [Green Version]
- Idris, N.M.; Gnanasammandhan, M.K.; Zhang, J.; Ho, P.C.; Mahendran, R.; Zhang, Y. In vivo photodynamic therapy using upconversion nanoparticles as remote-controlled nanotransducers. Nat. Med. 2012, 18, 1580–1585. [Google Scholar] [CrossRef]
- Rana, S.; Le, N.D.; Mout, R.; Saha, K.; Tonga, G.Y.; Bain, R.E.; Miranda, O.R.; Rotello, C.M.; Rotello, V.M. A multichannel nanosensor for instantaneous readout of cancer drug mechanisms. Nat. Nanotechnol. 2015, 10, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, Z.; Huang, D.; Liu, Z.; Guo, X.; Zhong, H. Synergistic effect of chemo-photothermal therapy using PEGylated graphene oxide. Biomaterials 2011, 32, 8555–8561. [Google Scholar] [CrossRef] [PubMed]
- Monopoli, M.P.; Walczyk, D.; Campbell, A.; Elia, G.; Lynch, I.; Baldelli Bombelli, F.; Dawson, K.A. Physical-Chemical aspects of protein corona: Relevance to in vitro and in vivo biological impacts of nanoparticles. J. Am. Chem. Soc. 2011, 133, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Walkey, C.D.; Olsen, J.B.; Guo, H.; Emili, A.; Chan, W.C.W. Nanoparticle size and surface chemistry determine serum protein adsorption and macrophage uptake. J. Am. Chem. Soc. 2012, 134, 2139–2147. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Kim, C.S.; Solfiell, D.J.; Rana, S.; Mout, R.; Velazquez-Delgado, E.M.; Chompoosor, A.; Jeong, Y.; Yan, B.; Zhu, Z.J.; et al. Direct Delivery of Functional Proteins and Enzymes to the Cytosol Using Nanoparticle-Stabilized Nanocapsules. ACS Nano 2013, 7, 6667–6673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, O.R.; Li, X.; Garcia-Gonzalez, L.; Zhu, Z.J.; Yan, B.; Bunz, U.H.; Rotello, V.M. Colorimetric bacteria sensing using a supramolecular enzyme-nanoparticle biosensor. J. Am. Chem. Soc. 2011, 133, 9650–9653. [Google Scholar] [CrossRef] [PubMed]
- Henzler, K.; Wittemann, A.; Breininger, E.; Ballauff, M.; Rosenfeldt, S. Adsorption of Bovine Hemoglobin onto Spherical Polyelectrolyte Brushes Monitored by Small-Angle X-ray Scattering and Fourier Transform Infrared Spectroscopy. Biomacromolecules 2007, 8, 3674–3681. [Google Scholar] [CrossRef]
- Henzler, K.; Haupt, B.; Ballauff, M. Enzymatic activity of immobilized enzyme determined by isothermal titration calorimetry. Anal. Biochem. 2008, 378, 184–189. [Google Scholar] [CrossRef]
- Wang, S.; Chen, K.; Kayitmazer, A.B.; Li, L.; Guo, X. Tunable adsorption of bovine serum albumin by annealed cationic spherical polyelectrolyte brushes. Colloids Surf. B Biointerfaces 2013, 107, 251–256. [Google Scholar] [CrossRef]
- Wang, S.; Chen, K.; Xu, Y.; Yu, X.; Wang, W.; Li, L.; Guo, X. Protein immobilization and separation using anionic/cationic spherical polyelectrolyte brushes based on charge anisotropy. Soft Matter 2013, 9, 11276–11287. [Google Scholar] [CrossRef]
- Kumar, S.; Tong, X.; Dory, Y.L.; Lepage, M.; Zhao, Y. A CO2-switchable polymer brush for reversible capture and release of proteins. Chem. Commun. 2013, 49, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Yan, B.; Zhang, L.; Wu, M.; Wang, J.; Huang, J.; Deng, Y.; Zeng, H. Tuning protein adsorption on charged polyelectrolyte brushes via salinity adjustment. Colloids Surf. A 2018, 539, 37–45. [Google Scholar] [CrossRef]
- Kayitmazer, A.B.; Strand, S.P.; Tribet, C.; Jaeger, W.; Dubin, P.L. Effect of Polyelectrolyte Structure on Protein−Polyelectrolyte Coacervates: Coacervates of Bovine Serum Albumin with Poly(diallyldimethylammonium chloride) versus Chitosan. Biomacromolecules 2007, 8, 3568–3577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.T.; Saha, K.; Kim, C.; Rotello, V.M. The Role of Surface Functionality in Determining Nanoparticle Cytotoxicity. Acc. Chem. Res. 2013, 46, 681–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyano, D.F.; Saha, K.; Prakash, G.; Yan, B.; Kong, H.; Yazdani, M.; Rotello, V.M. Fabrication of Corona-Free Nanoparticles with Tunable Hydrophobicity. Acs Nano 2014, 8, 6748–6755. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.C.; Rana, S.; Mout, R.; Yan, B.; Alfonso, F.S.; Rotello, V.M. Supramolecular tailoring of protein-nanoparticle interactions using cucurbituril mediators. Chem. Commun. 2014, 50, 5565–5568. [Google Scholar] [CrossRef] [PubMed]
- De, M.; Rana, S.; Akpinar, H.; Miranda, O.R.; Arvizo, R.R.; Bunz, U.H.F.; Rotello, V.M. Sensing of proteins in human serum using conjugates of nanoparticles and green fluorescent protein. Nat. Chem. 2009, 1, 461. [Google Scholar] [CrossRef] [PubMed]
- Duncan, B.; Le, N.D.B.; Alexander, C.; Gupta, A.; Yesilbag Tonga, G.; Yazdani, M.; Landis, R.F.; Wang, L.-S.; Yan, B.; Burmaoglu, S.; et al. Sensing by Smell: Nanoparticle–Enzyme Sensors for Rapid and Sensitive Detection of Bacteria with Olfactory Output. ACS Nano 2017, 11, 5339–5343. [Google Scholar] [CrossRef] [PubMed]
- Miranda, O.R.; Chen, H.T.; You, C.C.; Mortenson, D.E.; Yang, X.C.; Bunz, U.H.F.; Rotello, V.M. Enzyme-Amplified Array Sensing of Proteins in Solution and in Biofluids. J. Am. Chem. Soc. 2010, 132, 5285–5289. [Google Scholar] [CrossRef] [Green Version]
- Rana, S.; Elci, S.G.; Mout, R.; Singla, A.K.; Yazdani, M.; Bender, M.; Bajaj, A.; Saha, K.; Bunz, U.H.F.; Jirik, F.R.; et al. Ratiometric Array of Conjugated Polymers–Fluorescent Protein Provides a Robust Mammalian Cell Sensor. J. Am. Chem. Soc. 2016, 138, 4522–4529. [Google Scholar] [CrossRef] [Green Version]
- Li, X.N.; Kong, H.; Mout, R.; Saha, K.; Moyano, D.F.; Robinson, S.M.; Rana, S.; Zhang, X.R.; Riley, M.A.; Rotello, V.M. Rapid Identification of Bacterial Biofilms and Biofilm Wound Models Using a Multichannel Nanosensor. ACS Nano 2014, 8, 12014–12019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, S.; Singla, A.K.; Bajaj, A.; Elci, S.G.; Miranda, O.R.; Mout, R.; Yan, B.; Jirik, F.R.; Rotello, V.M. Array-Based Sensing of Metastatic Cells and Tissues Using Nanoparticle–Fluorescent Protein Conjugates. ACS Nano 2012, 6, 8233–8240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajaj, A.; Miranda, O.R.; Kim, I.-B.; Phillips, R.L.; Jerry, D.J.; Bunz, U.H.F.; Rotello, V.M. Detection and differentiation of normal, cancerous, and metastatic cells using nanoparticle-polymer sensor arrays. Proc. Natl. Acad. Sci. USA 2009, 106, 10912. [Google Scholar] [CrossRef] [PubMed]
- Creran, B.; Li, X.; Duncan, B.; Kim, C.S.; Moyano, D.F.; Rotello, V.M. Detection of bacteria using inkjet-printed enzymatic test strips. ACS Appl. Mater. Interfaces 2014, 6, 19525–19530. [Google Scholar] [CrossRef] [PubMed]
- Peveler, W.J.; Landis, R.F.; Yazdani, M.; Day, J.W.; Modi, R.; Carmalt, C.J.; Rosenberg, W.M.; Rotello, V.M. A Rapid and Robust Diagnostic for Liver Fibrosis Using a Multichannel Polymer Sensor Array. Adv. Mater. 2018, 30, 1800634. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymer/Protein | N | Kobs (M−1) | △H (kcal/mol) | T△S (cal/mol) |
|---|---|---|---|---|
| HA/BSA | 38 ± 1 | 389 ± 31 | 4.77 ± 0.01 | 8.30 ± 0.01 |
| PDADMAC/BSA | 80 ± 2 | 740 ± 30 | −4.15 ± 0.02 | −0.26 ± 0.03 |
| HA/BLG | 51 ± 1 | 228 ± 22 | 2.97 ± 0.01 | 6.18 ± 0.02 |
| PDADMAC/BLG | 50 ± 1 | 1900 ± 340 | −4.67 ± 0.02 | −0.2 ± 0.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zheng, K.; Si, Y.; Guo, X.; Xu, Y. Protein–Polyelectrolyte Interaction: Thermodynamic Analysis Based on the Titration Method †. Polymers 2019, 11, 82. https://doi.org/10.3390/polym11010082
Wang X, Zheng K, Si Y, Guo X, Xu Y. Protein–Polyelectrolyte Interaction: Thermodynamic Analysis Based on the Titration Method †. Polymers. 2019; 11(1):82. https://doi.org/10.3390/polym11010082
Chicago/Turabian StyleWang, Xiaohan, Kai Zheng, Yi Si, Xuhong Guo, and Yisheng Xu. 2019. "Protein–Polyelectrolyte Interaction: Thermodynamic Analysis Based on the Titration Method †" Polymers 11, no. 1: 82. https://doi.org/10.3390/polym11010082